
Sugar Coating: Utilisation of Host Serum Sialoglycoproteins by Schistosoma mansoni as a Potential Immune Evasion Mechanism


Abstract
:1. Introduction
2. Results
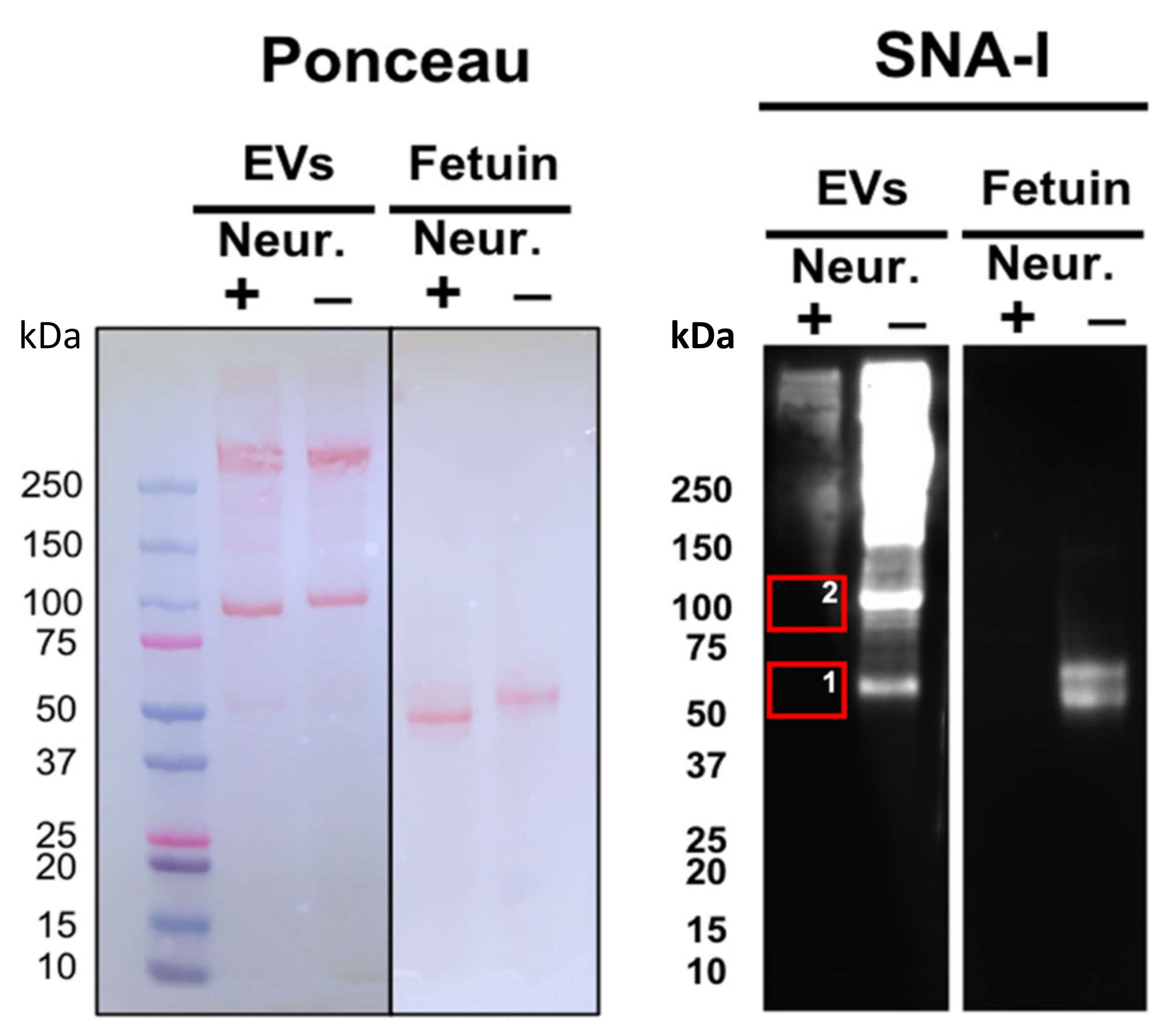
2.1. Lectin-Probed Western Blots and Mass Spectrometry Analysis of Reactive Bands
2.2. Identification of Glycosylated Proteins
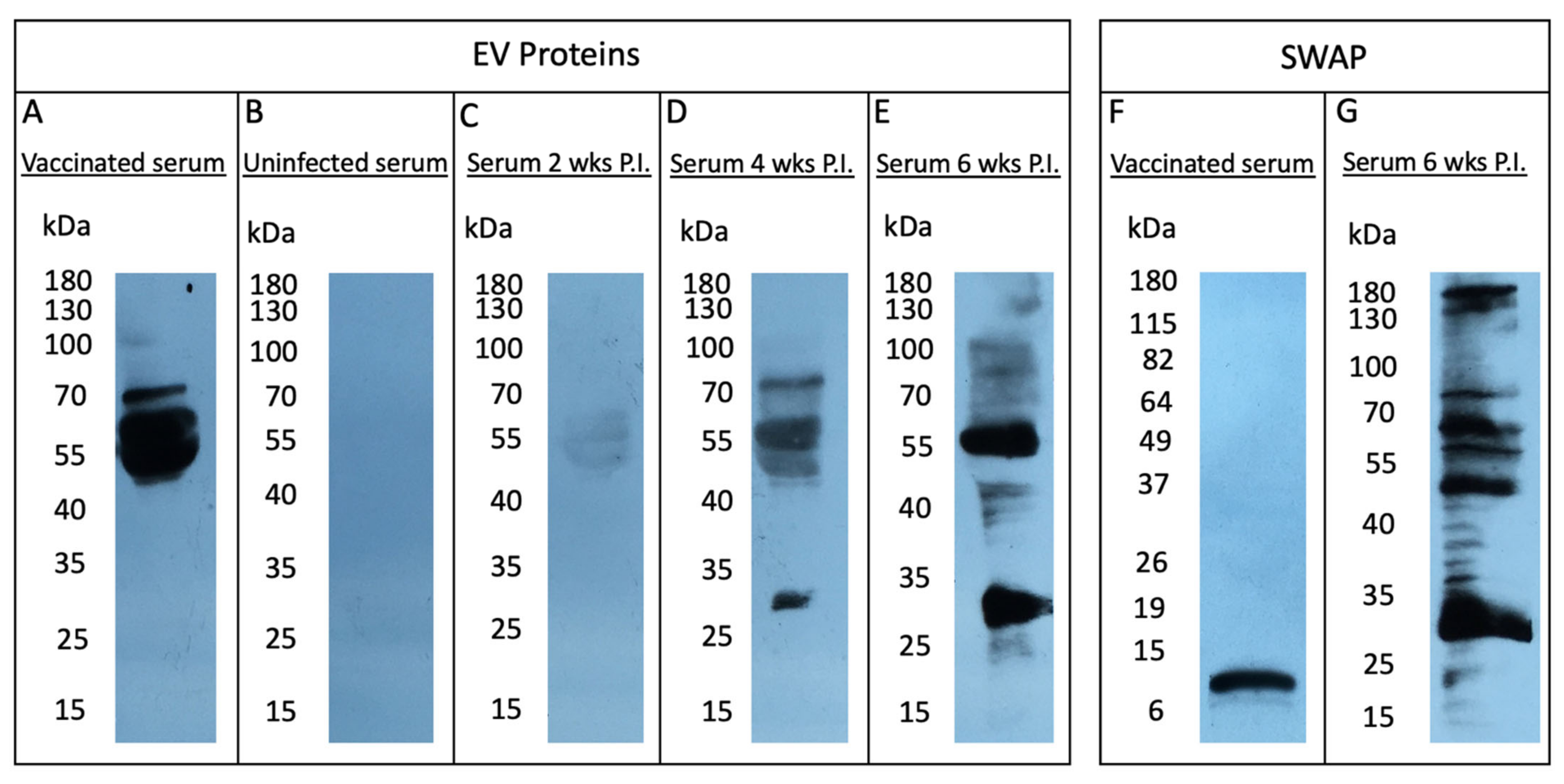
2.3. Sialylated EV Proteins Do Not Elicit a Humoral Response in the Murine Host
3. Discussion
4. Materials and Methods
4.1. Parasites
4.2. Isolation of Adult Schistosoma mansoni Extracellular Vesicles
4.3. Neuraminidase Treatment
4.4. Lectin-Probed Western Blot Assay and Extraction of Reactive Bands
4.5. Lectin Pulldown
4.6. Protein Digestion
4.7. Mass Spectrometry
4.8. Database Searching
4.9. Criteria for Protein Identification
4.10. Soluble Worm Antigenic Preparation
4.11. Mouse Infection and Vaccination
4.12. Immunogenicity Western Blot Assays
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mutapi, F.; Maizels, R.; Fenwick, A.; Woolhouse, M. Human schistosomiasis in the post mass drug administration era. Lancet Infect. Dis. 2017, 17, e42–e48. [Google Scholar] [CrossRef] [Green Version]
- Colley, D.G.; Bustinduy, A.L.; Secor, W.E.; King, C.H. Human schistosomiasis. Lancet 2014, 383, 2253–2264. [Google Scholar] [CrossRef]
- Harris, A.R.; Russell, R.J.; Charters, A.D. A review of schistosomiasis in immigrants in Western Australia, demonstrating the unusual longevity of Schistosoma mansoni. Trans. R. Soc. Trop. Med. Hyg. 1984, 78, 385–388. [Google Scholar] [CrossRef]
- Pearce, E.; Sher, A. Mechanisms of immune evasion in schistosomiasis. Contrib. Microbiol. Immunol. 1987, 8, 219–232. [Google Scholar] [PubMed]
- Zaccone, P.; Burton, O.T.; Gibbs, S.; Miller, N.; Jones, F.M.; Dunne, D.W.; Cooke, A. Immune modulation by Schistosoma mansoni antigens in NOD mice: Effects on both innate and adaptive immune systems. J. Biomed. Biotechnol. 2010, 2010, 795210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samoil, V.; Dagenais, M.; Ganapathy, V.; Aldridge, J.; Glebov, A.; Jardim, A.; Ribeiro, P. Vesicle-based secretion in schistosomes: Analysis of protein and microRNA (miRNA) content of exosome-like vesicles derived from Schistosoma mansoni. Sci. Rep. 2018, 8, 3286. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, F.C.; Swain, M.T.; Klychnikov, O.I.; Niazi, U.; Ivens, A.; Quintana, J.F.; Hensbergen, P.J.; Hokke, C.H.; Buck, A.H.; Hoffmann, K.F. Protein and small non-coding RNA-enriched extracellular vesicles are released by the pathogenic blood fluke Schistosoma mansoni. J. Extracell. Vesicles 2015, 4, 28665. [Google Scholar] [CrossRef] [Green Version]
- Sotillo, J.; Pearson, M.; Potriquet, J.; Becker, L.; Pickering, D.; Mulvenna, J.; Loukas, A. Extracellular vesicles secreted by Schistosoma mansoni contain protein vaccine candidates. Int. J. Parasitol. 2016, 46, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Kifle, D.W.; Pearson, M.S.; Becker, L.; Pickering, D.; Loukas, A.; Sotillo, J. Proteomic analysis of two populations of Schistosoma mansoni-derived extracellular vesicles: 15k pellet and 120k pellet vesicles. Mol. Biochem. Parasitol. 2020, 236, 111264. [Google Scholar] [CrossRef]
- Kuipers, M.E.; Nolte-’t Hoen, E.N.; van der Ham, A.J.; Ozir-Fazalalikhan, A.; Nguyen, D.L.; de Korne, C.M.; Koning, R.I.; Tomes, J.J.; Hoffmann, K.F.; Smits, H.H.; et al. DC-SIGN mediated internalisation of glycosylated extracellular vesicles from Schistosoma mansoni increases activation of monocyte-derived dendritic cells. J. Extracell. Vesicles 2020, 9, 1753420. [Google Scholar] [CrossRef]
- Reamtong, O.; Simanon, N.; Thiangtrongjit, T.; Limpanont, Y.; Chusongsang, P.; Chusongsang, Y.; Anuntakarun, S.; Payungporn, S.; Phuphisut, O.; Adisakwattana, P. Proteomic analysis of adult Schistosoma mekongi somatic and excretory-secretory proteins. Acta Trop. 2020, 202, 105247. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, Z.; Shen, J.; Liu, Z.; Liang, J.; Wu, X.; Sun, X.; Wu, Z. Exosome-like vesicles derived by Schistosoma japonicum adult worms mediates M1 type immune-activity of macrophage. Parasitol. Res. 2015, 114, 1865–1873. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Liu, J.; Dao, J.; Lu, K.; Li, H.; Gu, H.; Liu, J.; Feng, X.; Cheng, G. Molecular characterization of S. japonicum exosome-like vesicles reveals their regulatory roles in parasite-host interactions. Sci. Rep. 2016, 6, 25885. [Google Scholar] [CrossRef] [PubMed]
- Sotillo, J.; Robinson, M.W.; Kimber, M.J.; Cucher, M.; Ancarola, M.E.; Nejsum, P.; Marcilla, A.; Eichenberger, R.M.; Tritten, L. The protein and microRNA cargo of extracellular vesicles from parasitic helminths-current status and research priorities. Int. J. Parasitol. 2020, 50, 635–645. [Google Scholar] [CrossRef]
- Du, P.; Giri, B.R.; Liu, J.; Xia, T.; Grevelding, C.G.; Cheng, G. Proteomic and deep sequencing analysis of extracellular vesicles isolated from adult male and female Schistosoma japonicum. PLoS Negl. Trop. Dis. 2020, 14, e0008618. [Google Scholar] [CrossRef]
- Dagenais, M.; Gerlach, J.Q.; Collins III, J.J.; Atkinson, L.E.; Mousley, A.; Geary, T.G.; Long, T. Analysis of Schistosoma mansoni Extracellular Vesicles Surface Glycans Reveals Potential Immune Evasion Mechanism and New Insights on Their Origins of Biogenesis. Pathogens 2021, 10, 1401. [Google Scholar] [CrossRef]
- De la Torre-Escudero, E.; Gerlach, J.Q.; Bennett, A.P.; Cwiklinski, K.; Jewhurst, H.L.; Huson, K.M.; Joshi, L.; Kilcoyne, M.; O’Neill, S.; Dalton, J.P.; et al. Surface molecules of extracellular vesicles secreted by the helminth pathogen Fasciola hepatica direct their internalisation by host cells. PLoS Negl. Trop. Dis. 2019, 13, e0007087. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.; Pazos, R.; Royo, F.; González, E.; Roura-Ferrer, M.; Martinez, A.; Gamiz, J.; Reichardt, N.-C.; Falcón-Pérez, J.M. Assessing the role of surface glycans of extracellular vesicles on cellular uptake. Sci. Rep. 2019, 9, 11920. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.; Royo, F.; Aizpurua-Olaizola, O.; Pazos, R.; Boons, G.-J.; Reichardt, N.-C.; Falcon-Perez, J.M. Glycosylation of extracellular vesicles: Current knowledge, tools and clinical perspectives. J. Extracell. Vesicles 2018, 7, 1442985. [Google Scholar] [CrossRef]
- Shimoda, A.; Tahara, Y.; Sawada, S.-i.; Sasaki, Y.; Akiyoshi, K. Glycan profiling analysis using evanescent-field fluorescence-assisted lectin array: Importance of sugar recognition for cellular uptake of exosomes from mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2017, 491, 701–707. [Google Scholar] [CrossRef]
- Hokke, C.H.; van Diepen, A. Helminth glycomics–glycan repertoires and host-parasite interactions. Mol. Biochem. Parasitol. 2017, 215, 47–57. [Google Scholar] [CrossRef] [PubMed]
- McVeigh, P.; Cwiklinski, K.; Garcia-Campos, A.; Mulcahy, G.; O’Neill, S.M.; Maule, A.G.; Dalton, J.P. In silico analyses of protein glycosylating genes in the helminth Fasciola hepatica (liver fluke) predict protein-linked glycan simplicity and reveal temporally-dynamic expression profiles. Sci. Rep. 2018, 8, 11700. [Google Scholar] [CrossRef] [PubMed]
- Schauer, R.; Kamerling, J.P. Exploration of the sialic acid world. Adv. Carbohydr. Chem. Biochem. 2018, 75, 1–213. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Zhang, X.Q.; Lo, C.W.; Liu, P.F.; Liu, Y.T.; Gallo, R.L.; Hsieh, M.F.; Schooley, R.T.; Huang, C.M. The essentiality of α-2-macroglobulin in human salivary innate immunity against new H1N1 swine origin influenza A virus. Proteomics 2010, 10, 2396–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvert, L.; Atkinson, H.; Berry, L.; Chan, A. Age-dependent variation in glycosylation features of alpha-2-macroglobulin. Cell Biochem. Biophys. 2019, 77, 335–342. [Google Scholar] [CrossRef]
- Kragh-Hansen, U.; Donaldson, D.; Jensen, P.H. The glycan structure of albumin Redhill, a glycosylated variant of human serum albumin. Biochim. Et Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 2001, 1550, 20–26. [Google Scholar] [CrossRef]
- Kolarich, D.; Weber, A.; Turecek, P.L.; Schwarz, H.P.; Altmann, F. Comprehensive glyco-proteomic analysis of human α1-antitrypsin and its charge isoforms. Proteomics 2006, 6, 3369–3380. [Google Scholar] [CrossRef]
- Coddeville, B.; Stratil, A.; Wieruszeski, J.M.; Strecker, G.; Montreuil, J.; Spik, G. Primary structure of horse serotransferrin glycans: Demonstration that heterogeneity is related to the number of glycans and to the presence of N-acetylneuraminic acid and N-acetyl-4-O-acetylneuraminic acid. Eur. J. Biochem. 1989, 186, 583–590. [Google Scholar] [CrossRef]
- Wubbolts, R.; Leckie, R.S.; Veenhuizen, P.T.; Schwarzmann, G.; Mobius, W.; Hoernschemeyer, J.; Slot, J.-W.; Geuze, H.J.; Stoorvogel, W. Proteomic and biochemical analyses of human B cell-derived exosomes: Potential implications for their function and multivesicular body formation. J. Biol. Chem. 2003, 278, 10963–10972. [Google Scholar] [CrossRef] [Green Version]
- Buschow, S.I.; van Balkom, B.W.; Aalberts, M.; Heck, A.J.; Wauben, M.; Stoorvogel, W. MHC class II-associated proteins in B-cell exosomes and potential functional implications for exosome biogenesis. Immunol. Cell Biol. 2010, 88, 851–856. [Google Scholar] [CrossRef]
- Hassani, K.; Olivier, M. Immunomodulatory impact of leishmania-induced macrophage exosomes: A comparative proteomic and functional analysis. PLoS Negl. Trop. Dis. 2013, 7, e2185. [Google Scholar] [CrossRef] [PubMed]
- Steenbeek, S.C.; Pham, T.V.; de Ligt, J.; Zomer, A.; Knol, J.C.; Piersma, S.R.; Schelfhorst, T.; Huisjes, R.; Schiffelers, R.M.; Cuppen, E. Cancer cells copy migratory behavior and exchange signaling networks via extracellular vesicles. EMBO J. 2018, 37, e98357. [Google Scholar] [CrossRef] [PubMed]
- Nelwan, M.L. Schistosomiasis: Life cycle, diagnosis, and control. Curr. Ther. Res. 2019, 91, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Duguet, T.B.; Soichot, J.; Kuzyakiv, R.; Malmström, L.; Tritten, L. Extracellular vesicle-contained microRNA of C. elegans as a tool to decipher the molecular basis of nematode parasitism. Front. Cell. Infect. Microbiol. 2020, 10, 217. [Google Scholar] [CrossRef]
- Drurey, C.; Maizels, R.M. Helminth extracellular vesicles: Interactions with the host immune system. Mol. Immunol. 2021, 137, 124–133. [Google Scholar] [CrossRef]
- Whitehead, B.; Boysen, A.T.; Mardahl, M.; Nejsum, P. Unique glycan and lipid composition of helminth-derived extracellular vesicles may reveal novel roles in host-parasite interactions. Int. J. Parasitol. 2020, 50, 647–654. [Google Scholar] [CrossRef]
- Simbari, F.; McCaskill, J.; Coakley, G.; Millar, M.; Maizels, R.M.; Fabriás, G.; Casas, J.; Buck, A.H. Plasmalogen enrichment in exosomes secreted by a nematode parasite versus those derived from its mouse host: Implications for exosome stability and biology. J. Extracell. Vesicles 2016, 5, 30741. [Google Scholar] [CrossRef] [Green Version]
- Khoo, K.-H.; Nieto, A.; Morris, H.R.; Dell, A. Structural characterization of the N-glycans from Echinococcus granulosus hydatid cyst membrane and protoscoleces. Mol. Biochem. Parasitol. 1997, 86, 237–248. [Google Scholar] [CrossRef]
- Lee, J.J.; Dissanayake, S.; Panico, M.; Morris, H.R.; Dell, A.; Haslam, S.M. Mass spectrometric characterisation of Taenia crassiceps metacestode N-glycans. Mol. Biochem. Parasitol. 2005, 143, 245–249. [Google Scholar] [CrossRef]
- Ravidà, A.; Aldridge, A.M.; Driessen, N.N.; Heus, F.A.; Hokke, C.H.; O’Neill, S.M. Fasciola hepatica surface coat glycoproteins contain mannosylated and phosphorylated N-glycans and exhibit immune modulatory properties independent of the mannose receptor. PLoS Negl. Trop. Dis. 2016, 10, e0004601. [Google Scholar] [CrossRef] [Green Version]
- Schauer, R. Sialic acids and their role as biological masks. Trends Biochem. Sci. 1985, 10, 357–360. [Google Scholar] [CrossRef]
- Pilatte, Y.; Bignon, J.; Lambre, C.R. Sialic acids as important molecules in the regulation of the immune system: Pathophysiological implications of sialidases in immunity. Glycobiology 1993, 3, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Varki, A.; Kannagi, R.; Toole, B.P. Glycosylation changes in cancer. In Essentials of Glycobiology, 2nd ed.; Cold Spring Harbor Laboratory Press: Long Island, NY, USA, 2009. [Google Scholar]
- Häuselmann, I.; Borsig, L. Altered tumor-cell glycosylation promotes metastasis. Front. Oncol. 2014, 4, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büll, C.; den Brok, M.H.; Adema, G.J. Sweet escape: Sialic acids in tumor immune evasion. Biochim. Et Biophys. Acta (BBA)-Rev. Cancer 2014, 1846, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Crocker, P.R.; Paulson, J.C.; Varki, A. Siglecs and their roles in the immune system. Nat. Rev. Immunol. 2007, 7, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Varki, A.; Gagneux, P. Multifarious roles of sialic acids in immunity. Ann. N. Y. Acad. Sci. 2012, 1253, 16. [Google Scholar] [CrossRef] [Green Version]
- Meri, S.; Pangburn, M.K. Discrimination between activators and nonactivators of the alternative pathway of complement: Regulation via a sialic acid/polyanion binding site on factor H. Proc. Natl. Acad. Sci. USA 1990, 87, 3982–3986. [Google Scholar] [CrossRef] [Green Version]
- Severi, E.; Hood, D.W.; Thomas, G.H. Sialic acid utilization by bacterial pathogens. Microbiology 2007, 153, 2817–2822. [Google Scholar] [CrossRef] [Green Version]
- Schauer, R.; Kamerling, J.P. The chemistry and biology of trypanosomal trans-sialidases: Virulence factors in Chagas disease and sleeping sickness. ChemBioChem 2011, 12, 2246–2264. [Google Scholar] [CrossRef]
- Rempel, H.; Calosing, C.; Sun, B.; Pulliam, L. Sialoadhesin expressed on IFN-induced monocytes binds HIV-1 and enhances infectivity. PLoS ONE 2008, 3, e1967. [Google Scholar] [CrossRef]
- Corfield, T. Bacterial sialidases—roles in pathogenicity and nutrition. Glycobiology 1992, 2, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Alviano, C.S.; Travassos, L.R.; Schauer, R. Sialic acids in fungi. Glycoconj. J. 1999, 16, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Eneva, R.; Engibarov, S.; Abrashev, R.; Krumova, E.; Angelova, M. Sialic acids, sialoconjugates and enzymes of their metabolism in fungi. Biotechnol. Biotechnol. Equip. 2021, 35, 364–375. [Google Scholar] [CrossRef]
- Mehr, K.; Withers, S.G. Mechanisms of the sialidase and trans-sialidase activities of bacterial sialyltransferases from glycosyltransferase family 80. Glycobiology 2016, 26, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Bouchet, V.; Hood, D.W.; Li, J.; Brisson, J.-R.; Randle, G.A.; Martin, A.; Li, Z.; Goldstein, R.; Schweda, E.K.; Pelton, S.I. Host-derived sialic acid is incorporated into Haemophilus influenzae lipopolysaccharide and is a major virulence factor in experimental otitis media. Proc. Natl. Acad. Sci. USA 2003, 100, 8898–8903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohanpal, B.K.; El-Labany, S.; Lahooti, M.; Plumbridge, J.A.; Blomfield, I.C. Integrated regulatory responses of fimB to N-acetylneuraminic (sialic) acid and GlcNAc in Escherichia coli K-12. Proc. Natl. Acad. Sci. USA 2004, 101, 16322–16327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohanpal, B.K.; Friar, S.; Roobol, J.; Plumbridge, J.A.; Blomfield, I.C. Multiple co-regulatory elements and IHF are necessary for the control of fimB expression in response to sialic acid and N-acetylglucosamine in Escherichia coli K-12. Mol. Microbiol. 2007, 63, 1223–1236. [Google Scholar] [CrossRef]
- Shakhnovich, E.A.; King, S.J.; Weiser, J.N. Neuraminidase expressed by Streptococcus pneumoniae desialylates the lipopolysaccharide of Neisseria meningitidis and Haemophilus influenzae: A paradigm for interbacterial competition among pathogens of the human respiratory tract. Infect. Immun. 2002, 70, 7161–7164. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, M.E.; de Lederkremer, R.M. Trans-sialidase and mucins of Trypanosoma cruzi: An important interplay for the parasite. Carbohydr. Res. 2011, 346, 1389–1393. [Google Scholar] [CrossRef]
- Dias, W.B.; Fajardo, F.D.; Graça-Souza, A.V.; Freire-de-Lima, L.; Vieira, F.; Girard, M.F.; Bouteille, B.; Previato, J.O.; Mendonça-Previato, L.; Todeschini, A.R. Endothelial cell signalling induced by trans-sialidase from Trypanosoma cruzi. Cell. Microbiol. 2008, 10, 88–99. [Google Scholar] [CrossRef]
- Magdesian, M.H.; Tonelli, R.R.; Fessel, M.R.; Silveira, M.S.; Schumacher, R.I.; Linden, R.; Colli, W.; Alves, M.J.M. A conserved domain of the gp85/trans-sialidase family activates host cell extracellular signal-regulated kinase and facilitates Trypanosoma cruzi infection. Exp. Cell Res. 2007, 313, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Rubin-de-Celis, S.S.; Uemura, H.; Yoshida, N.; Schenkman, S. Expression of trypomastigote trans-sialidase in metacyclic forms of Trypanosoma cruzi increases parasite escape from its parasitophorous vacuole. Cell. Microbiol. 2006, 8, 1888–1898. [Google Scholar] [CrossRef] [PubMed]
- Vanderheijden, N.; Delputte, P.L.; Favoreel, H.W.; Vandekerckhove, J.; van Damme, J.; van Woensel, P.A.; Nauwynck, H.J. Involvement of sialoadhesin in entry of porcine reproductive and respiratory syndrome virus into porcine alveolar macrophages. J. Virol. 2003, 77, 8207–8215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornilov, R.; Puhka, M.; Mannerström, B.; Hiidenmaa, H.; Peltoniemi, H.; Siljander, P.; Seppänen-Kaijansinkko, R.; Kaur, S. Efficient ultrafiltration-based protocol to deplete extracellular vesicles from fetal bovine serum. J. Extracell. Vesicles 2018, 7, 1422674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laemmli, U. SDS-page Laemmli method. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Total Spectrum Count | |||
|---|---|---|---|
| Accession Number | Identified Proteins | Band 1 | Band 2 |
| A0A140T897 | Albumin [Bt] | 17 | 233 |
| P34955 | Alpha-1-antiproteinase [Bt] | 7 | 62 |
| F1MM86 (+1) | Complement component C6 [Bt] | 13 | |
| Q7SIH1 | Alpha-2-macroglobulin [Bt] | 4 | 8 |
| A0A3Q1MU98 (+1) | Complement component C9 [Bt] | 4 | |
| A0A3Q1NJR8 | Antithrombin-III [Bt] | 7 | |
| A0A3Q1LQ21 (+1) | Inter-alpha-trypsin inhibitor heavy chain H3 [Bt] | 6 | |
| G3X6N3 | Beta-1 metal-binding globulin [Bt] | 3 | 2 |
| UniProt Accession Number | Identified Protein | Total Spectrum Count | Unique Peptides |
|---|---|---|---|
| A2MG_BOVIN | Alpha-2-macroglobulin [Bt] | 65 | 38 |
| A1AT_BOVIN | Alpha-1-antiproteinase [Bt] | 46 | 18 |
| F1MJK3_BOVIN | Uncharacterised protein LOC506828 [Bt] | 13 | 10 |
| CLUS_BOVIN | Clusterin [Bt] | 10 | 9 |
| A0A3Q1LK49_BOVIN | Inter-alpha-trypsin inhibitor heavy chain H2 [Bt] | 10 | 9 |
| HBBF_BOVIN | Hemoglobin fetal subunit beta [Bt] | 10 | 8 |
| A0A3Q1M2B2_BOVIN | Complement C3 [Bt] | 7 | 6 |
| A0A3Q1LVV7_BOVIN | Fibrinogen alpha chain [Bt] | 6 | 6 |
| FETUA_BOVIN | Alpha-2-HS-glycoprotein [Bt] | 6 | 6 |
| A0A3Q1LQ21_BOVIN | Inter-alpha-trypsin inhibitor heavy chain H3 [Bt] | 6 | 6 |
| ACTB_BOVIN | Actin, cytoplasmic 1 [Bt] | 6 | 5 |
| A0A3Q1MLQ7_BOVIN | Talin 1 [Bt] | 6 | 5 |
| A0A3Q1LKN2_BOVIN | Thyroglobulin [Bt] | 6 | 4 |
| A0A140T881_BOVIN | Apolipoprotein E [Bt] | 3 | 3 |
| A0A3Q1LGY9_BOVIN | Angiotensinogen [Bt] | 3 | 3 |
| APOA1_BOVIN | Apolipoprotein A-I [Bt] | 3 | 3 |
| A2AP_BOVIN | Alpha-2-antiplasmin [Bt] | 3 | 3 |
| G3P_BOVIN | Glyceraldehyde-3-phosphate dehydrogenase [Bt] | 3 | 3 |
| Q3ZBS7_BOVIN | Vitronectin [Bt] | 2 | 2 |
| B7FAU9 | Filamin, alpha [Mm] | 2 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dagenais, M.; Gerlach, J.Q.; Geary, T.G.; Long, T. Sugar Coating: Utilisation of Host Serum Sialoglycoproteins by Schistosoma mansoni as a Potential Immune Evasion Mechanism. Pathogens 2022, 11, 426. https://doi.org/10.3390/pathogens11040426
Dagenais M, Gerlach JQ, Geary TG, Long T. Sugar Coating: Utilisation of Host Serum Sialoglycoproteins by Schistosoma mansoni as a Potential Immune Evasion Mechanism. Pathogens. 2022; 11(4):426. https://doi.org/10.3390/pathogens11040426
Chicago/Turabian StyleDagenais, Maude, Jared Q. Gerlach, Timothy G. Geary, and Thavy Long. 2022. "Sugar Coating: Utilisation of Host Serum Sialoglycoproteins by Schistosoma mansoni as a Potential Immune Evasion Mechanism" Pathogens 11, no. 4: 426. https://doi.org/10.3390/pathogens11040426