
Babesiosis and Theileriosis in North America



Abstract
:1. Introduction
2. Babesia species in North America
General Life Cycle of Babesia
3. Human Babesiosis
3.1. Pathogenesis
3.2. Diagnosis
3.3. Epidemiology
4. Bovine Babesiosis
4.1. Pathogenesis
4.2. Diagnosis
4.3. Epidemiology
5. Canine Babesiosis
5.1. Pathogenesis
5.2. Diagnosis
5.3. Epidemiology
6. Babesiosis in Cervids
6.1. Pathogenesis
6.2. Diagnosis
6.3. Epidemiology
7. Equine Piroplasmosis
7.1. Pathogenesis
7.2. Diagnosis
7.3. Epidemiology
8. Theileriosis in Cattle
8.1. Pathogenesis
8.2. Diagnosis
8.3. Epidemiology
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nicholson, W.L.; Sonenshine, D.E.; Noden, B.H.; Brown, R.N. Ticks (Ixodida). In Medical and Veterinary Entomology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 603–672. [Google Scholar]
- Sonenshine, D.E. Range Expansion of Tick Disease Vectors in North America: Implications for Spread of Tick-Borne Disease. Int. J. Environ. Res. Public Health 2018, 15, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guglielmone, A.A.; Robbins, R.G. Hard Ticks (Acari: Ixodida: Ixodidae) Parasitizing Humans; Springer International Publishing: Cham, Switzerland, 2018; Volume 230. [Google Scholar]
- Bullard, J.M.; Ahsanuddin, A.N.; Perry, A.M.; Lindsay, L.R.; Iranpour, M.; Dibernardo, A.; Van Caeseele, P.G. The First Case of Locally Acquired Tick-Borne Babesia Microti Infection in Canada. Can. J. Infect. Dis. Med. Microbiol. 2014, 25, e87–e89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, D.A.; Thomas, K.R.; Jajosky, R.A.; Foster, L.; Sharp, P.; Onweh, D.H.; Schley, A.W.; Anderson, W.J. Summary of Notifiable Infectious Diseases and Conditions—United States. MMWR 2014, 63, 54. [Google Scholar]
- Bishop, A.; Wang, H.-H.; Grant, W.E. Using Data Surveillance to Understand the Rising Incidence of Babesiosis in the United States, 2011–2018. Vector-Borne Zoonotic Dis. 2021, 21, 391–395. [Google Scholar] [CrossRef]
- Krause, P.J.; Spielman, A.; Telford, S.R.; Sikand, V.K.; McKay, K.; Christianson, D.; Pollack, R.J.; Brassard, P.; Magera, J.; Ryan, R. Persistent Parasitemia after Acute Babesiosis. N. Engl. J. Med. 1998, 339, 160–165. [Google Scholar] [CrossRef]
- Krause, P.J. Babesiosis Diagnosis and Treatment. Vector-Borne Zoonotic Dis. 2003, 3, 45–51. [Google Scholar] [CrossRef]
- Vannier, E.; Krause, P.J. Update on Babesiosis. Interdiscip. Perspect. Infect. Dis. 2009, 2009. [Google Scholar] [CrossRef] [Green Version]
- Swei, A.; O’Connor, K.E.; Couper, L.I.; Thekkiniath, J.; Conrad, P.A.; Padgett, K.A.; Burns, J.; Yoshimizu, M.H.; Gonzales, B.; Munk, B. Evidence for Transmission of the Zoonotic Apicomplexan Parasite Babesia Duncani by the Tick Dermacentor Albipictus. Int. J. Parasitol. 2019, 49, 95–103. [Google Scholar] [CrossRef]
- Krause, P.J. Human Babesiosis. Int. J. Parasitol. 2019, 49, 165–174. [Google Scholar] [CrossRef]
- Almazan, C.; Tipacamu, G.A.; Rodriguez, S.; Mosqueda, J.; Perez de Leon, A. Immunological Control of Ticks and Tick-Borne Diseases That Impact Cattle Health and Production. Front. Biosci. 2018, 23, 1535–1551. [Google Scholar] [CrossRef] [Green Version]
- de León, A.P.; Vannier, E.; Alamazán, C.; Krause, P.J. Tick-Borne Protozoa. In Biology of Ticks; Oxford University Press: New York, NY, USA, 2014; Voloume 2, pp. 147–179. [Google Scholar]
- Alvarez, J.A.; Rojas, C.; Figueroa, J.V. Diagnostic Tools for the Identification of Babesia Sp. in Persistently Infected Cattle. Pathogens 2019, 8, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starkey, L.A.; Little, S.E. Canine Tick-Borne Diseases. Infection 2015, 5, 55–60. [Google Scholar]
- Kjemtrup, A.M.; Wainwright, K.; Miller, M.; Penzhorn, B.L.; Carreno, R.A. Babesia Conradae, Sp. Nov., a Small Canine Babesia Identified in California. Vet. Parasitol. 2006, 138, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Barash, N.R.; Thomas, B.; Birkenheuer, A.J.; Breitschwerdt, E.B.; Lemler, E.; Qurollo, B.A. Prevalence of Babesia Spp. and Clinical Characteristics of Babesia Vulpes Infections in North American Dogs. J. Vet. Intern. Med. 2019, 33, 2075–2081. [Google Scholar] [CrossRef] [Green Version]
- Scoles, G.; Ueti, M. Vector Ecology of Equine Piroplasmosis. Annu. Rev. Entomol. 2015, 60, 561–580. [Google Scholar] [CrossRef]
- Osorno, B.M.; Solana, P. Aislamiento e Identificación de Babesia Equi y Babesia Caballi En Caballos de México. Rev. Mex. De Cienc. Pecu. 1972, 20, 39–41. [Google Scholar]
- Mathieu, A.; Pastor, A.R.; Berkvens, C.N.; Gara-Boivin, C.; Hébert, M.; Léveillé, A.N.; Barta, J.R.; Smith, D.A. Babesia Odocoilei as a Cause of Mortality in Captive Cervids in Canada. Can. Vet. J. 2018, 59, 52. [Google Scholar]
- Knowles, D.P.; Kappmeyer, L.S.; Haney, D.; Herndon, D.R.; Fry, L.M.; Munro, J.B.; Sears, K.; Ueti, M.W.; Wise, L.N.; Silva, M. Discovery of a Novel Species, Theileria Haneyi n. Sp., Infective to Equids, Highlights Exceptional Genomic Diversity within the Genus Theileria: Implications for Apicomplexan Parasite Surveillance. Int. J. Parasitol. 2018, 48, 679–690. [Google Scholar] [CrossRef]
- Oakes, V.J.; Yabsley, M.J.; Schwartz, D.; LeRoith, T.; Bissett, C.; Broaddus, C.; Schlater, J.L.; Todd, S.M.; Boes, K.M.; Brookhart, M. Theileria Orientalis Ikeda Genotype in Cattle, Virginia, USA. Emerg. Infect. Dis. 2019, 25, 1653. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.; Johnson, E.M.; Allen, K.E.; Campbell, G.A.; Rezabek, G.; Bradway, D.S.; Pittman, L.L.; Little, S.E.; Panciera, R.J. Merogonic Stages of Theileria Cervi in Mule Deer (Odocoileus Hemionus). J. Vet. Diagn. Investig. 2013, 25, 662–665. [Google Scholar] [CrossRef] [Green Version]
- Chae, J.; Levy, M.; Hunt, J., Jr.; Schlater, J.; Snider, G.; Waghela, S.D.; Holman, P.J.; Wagner, G.G. Theileria Sp. Infections Associated with Bovine Fatalities in the United States Confirmed by Small-Subunit RRNA Gene Analyses of Blood and Tick Samples. J. Clin. Microbiol. 1999, 37, 3037–3040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peniche-Lara, G.; Balmaceda, L.; Perez-Osorio, C.; Munoz-Zanzi, C. Human Babesiosis, Yucatán State, Mexico, 2015. Emerg. Infect. Dis. 2018, 24, 2061. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Vivas, R.I.; Grisi, L.; Pérez de León, A.A.; Villela, H.S.; Torres-Acosta, J.F.d.J.; Fragoso Sánchez, H.; Romero Salas, D.; Rosario Cruz, R.; Saldierna, F.; García Carrasco, D. Potential Economic Impact Assessment for Cattle Parasites in Mexico. Review. Rev. Mex. De Cienc. Pecu. 2017, 8, 61–74. [Google Scholar] [CrossRef]
- Esteve-Gasent, M.D.; Rodríguez-Vivas, R.I.; Medina, R.F.; Ellis, D.; Schwartz, A.; Cortés Garcia, B.; Hunt, C.; Tietjen, M.; Bonilla, D.; Thomas, D. Research on Integrated Management for Cattle Fever Ticks and Bovine Babesiosis in the United States and Mexico: Current Status and Opportunities for Binational Coordination. Pathogens 2020, 9, 871. [Google Scholar] [CrossRef] [PubMed]
- Dear, J.D.; Owens, S.D.; Lindsay, L.L.; Biondo, A.W.; Chomel, B.B.; Marcondes, M.; Sykes, J.E. Babesia Conradae Infection in Coyote Hunting Dogs Infected with Multiple Blood-Borne Pathogens. J. Vet. Intern. Med. 2018, 32, 1609–1617. [Google Scholar] [CrossRef]
- Schoelkopf, L.; Hutchinson, C.E.; Bendele, K.G.; Goff, W.L.; Willette, M.; Rasmussen, J.M.; Holman, P.J. New Ruminant Hosts and Wider Geographic Range Identified for Babesia Odocoilei (Emerson and Wright 1970). J. Wildl. Dis. 2005, 41, 683–690. [Google Scholar] [CrossRef] [Green Version]
- Short, M.A.; Clark, C.K.; Harvey, J.W.; Wenzlow, N.; Hawkins, I.K.; Allred, D.R.; Knowles, D.P.; Corn, J.L.; Grause, J.F.; Hennager, S.G. Outbreak of Equine Piroplasmosis in Florida. J. Am. Vet. Med. Assoc. 2012, 240, 588–595. [Google Scholar] [CrossRef]
- Scoles, G.A.; Hutcheson, H.J.; Schlater, J.L.; Hennager, S.G.; Pelzel, A.M.; Knowles, D.P. Equine Piroplasmosis Associated with Amblyomma Cajennense Ticks, Texas, USA. Emerg. Infect. Dis. 2011, 17, 1903. [Google Scholar] [CrossRef]
- Waldrup, K.A.; Collisson, E.; Bentsen, S.E.; Winkler, C.K.; Wagner, G.G. Prevalence of Erythrocytic Protozoa and Serologic Reactivity to Selected Pathogens in Deer in Texas. Prev. Vet. Med. 1989, 7, 49–58. [Google Scholar] [CrossRef]
- Pavón-Rocha, A.J.; Cárdenas-Flores, A.; Rábago-Castro, J.L.; Barrón-Vargas, C.A.; Mosqueda, J. First Molecular Evidence of Theileria Cervi Infection in White-Tailed Deer (Odocoileus Virginianus) in Mexico. Vet. Parasitol. Reg. Stud. Rep. 2020, 22, 100482. [Google Scholar] [CrossRef]
- Kjemtrup, A.M.; Conrad, P.A. Human Babesiosis: An Emerging Tick-Borne Disease. Int. J. Parasitol. 2000, 30, 1323–1337. [Google Scholar] [CrossRef]
- Bock, R.; Jackson, L.; de Vos, A.; Jorgensen, W. Babesiosis of Cattle. Parasitology 2004, 129 (Suppl. Sl), S247–S269. [Google Scholar] [CrossRef] [PubMed]
- Baneth, G. Antiprotozoal Treatment of Canine Babesiosis. Vet. Parasitol. 2018, 254, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Laha, R.; Das, M.; Sen, A. Morphology, Epidemiology, and Phylogeny of Babesia: An Overview. Trop. Parasitol. 2015, 5, 94. [Google Scholar] [CrossRef] [Green Version]
- Riek, R.F. The Life Cycle of Babesia Bigemina (Smith and Kilborne, 1893) in the Tick Vector Boophilus Microplus (Canestrini). Aust. J. Agric. Res. 1964, 15, 802–821. [Google Scholar] [CrossRef]
- Gohil, S.; Kats, L.M.; Sturm, A.; Cooke, B.M. Recent Insights into Alteration of Red Blood Cells by Babesia Bovis: Moovin’forward. Trends Parasitol. 2010, 26, 591–599. [Google Scholar] [CrossRef]
- Gray, E.B.; Herwaldt, B.L. Babesiosis Surveillance—United States, 2011–2015. MMWR Surveill. Summ. 2019, 68, 1–11. [Google Scholar] [CrossRef]
- Madison-Antenucci, S.; Kramer, L.D.; Gebhardt, L.L.; Kauffman, E. Emerging Tick-Borne Diseases. Clin. Microbiol. Rev. 2020, 33. [Google Scholar] [CrossRef]
- Shah, J.S.; Caoili, E.; Patton, M.F.; Tamhankar, S.; Myint, M.M.; Poruri, A.; Mark, O.; Horowitz, R.I.; Ashbaugh, A.D.; Ramasamy, R. Combined Immunofluorescence (IFA) and Fluorescence In Situ Hybridization (FISH) Assays for Diagnosing Babesiosis in Patients from the USA, Europe and Australia. Diagnostics 2020, 10, 761. [Google Scholar] [CrossRef]
- Jalovecka, M.; Sojka, D.; Ascencio, M.; Schnittger, L. Babesia Life Cycle–When Phylogeny Meets Biology. Trends Parasitol. 2019, 35, 356–368. [Google Scholar] [CrossRef]
- Osorno, B.M.; Ristic, M. Babesia Canis En Perros En México. Rev. Mex. De Cienc. Pecu. 1974, 1, 36. [Google Scholar]
- De Vos, A.J.; De Waal, D.T.; Jackson, L.A. Bovine Babesiosis. Infect. Dis. Livest. Vol. One 2004, 2, 406–424. [Google Scholar]
- Mosqueda, J.; Olvera-Ramirez, A.; Aguilar-Tipacamu, G.; Canto, G.J. Current Advances in Detection and Treatment of Babesiosis. Curr. Med. Chem. 2012, 19, 1504–1518. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, F.D.; Bendele, K.G.; Davey, R.B.; George, J.E. Detection of Babesia Bigemina Infection in Strains of Rhipicephalus (Boophilus) Microplus Collected from Outbreaks in South Texas. Vet. Parasitol. 2007, 145, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Ramos, J.A.; Alvarez, J.A.; Figueroa, J.V.; Solis, J.; Rodriguez, R.I.; Hernandez, R.; Buening, G.M.; Vega, C.A. Evaluation of a Colorimetric Babesia Bigemina-DNA Probe within an Epidemiological Survey. Mem. Do Inst. Oswaldo Cruz 1992, 87, 213–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solorio-Rivera, J.L.; Rodríguez-Vivas, R.I.; Pérez-Gutierrez, E.; Wagner, G. Management Factors Associated with Babesia Bovis Seroprevalence in Cattle from Eastern Yucatan, Mexico. Prev. Vet. Med. 1999, 40, 261–269. [Google Scholar] [CrossRef]
- Teclaw, R.F.; Romo, S.; Garcia, Z.; Castaneda, M.; Wagner, G.G. A Seroepidemiologic Study of Bovine Babesiosis in the Mexican States of Nuevo Leon, Tamaulipas and Coahuila. Prev. Vet. Med. 1985, 3, 403–415. [Google Scholar] [CrossRef]
- Romero-Salas, D.; Mira, A.; Mosqueda, J.; García-Vázquez, Z.; Hidalgo-Ruiz, M.; Vela, N.A.O.; de León, A.A.P.; Florin-Christensen, M.; Schnittger, L. Molecular and Serological Detection of Babesia Bovis-and Babesia Bigemina-Infection in Bovines and Water Buffaloes Raised Jointly in an Endemic Field. Vet. Parasitol. 2016, 217, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Vivas, R.I.; Cob-Galera, L.A.; Domínguez-Alpizar, J.L. Hemoparásitos En Bovinos, Caninos y Equinos Diagnosticados En El Laboratorio de Parasitología de La Facultad de Medicina Veterinaria y Zootecnia de La Universidad Autónoma de Yucatan (1984–1999). Rev. Biomédica 2000, 11, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Cantu-C, A.; Ortega-S, J.A.; García-Vázquez, Z.; Mosqueda, J.; Henke, S.E.; George, J.E. Epizootiology of Babesia Bovis and Babesia Bigemina in Free-Ranging White-Tailed Deer in Northeastern Mexico. J. Parasitol. 2009, 95, 536–542. [Google Scholar] [CrossRef]
- Ramos, C.M.; Cooper, S.M.; Holman, P.J. Molecular and Serologic Evidence for Babesia Bovis-like Parasites in White-Tailed Deer (Odocoileus Virginianus) in South Texas. Vet. Parasitol. 2010, 172, 214–220. [Google Scholar] [CrossRef]
- Holman, P.J.; Carroll, J.E.; Pugh, R.; Davis, D.S. Molecular Detection of Babesia Bovis and Babesia Bigemina in White-Tailed Deer (Odocoileus Virginianus) from Tom Green County in Central Texas. Vet. Parasitol. 2011, 177, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas-Canales, E.M.; Ortega-Santos, J.A.; Campbell, T.A.; García-Vázquez, Z.; Cantú-Covarrubias, A.; Figueroa-Millán, J.V.; DeYoung, R.W.; Hewitt, D.G.; Bryant, F.C. Nilgai Antelope in Northern Mexico as a Possible Carrier for Cattle Fever Ticks and Babesia Bovis and Babesia Bigemina. J. Wildl. Dis. 2011, 47, 777–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Vázquez, Z.; Ortega-S, J.A.; Cantu-Covarruvias, A.; Mosqueda, J.; Hewitt, D.G.; DeYoung, R.W.; Campbell, T.A.; Bryant, F.C. Tick-Borne Diseases in Syntopic Populations of Fallow Deer (Dama Dama) and Axis Deer (Axis Axis) in Northern Mexico. J. Wildl. Dis. 2015, 51, 527–529. [Google Scholar] [CrossRef] [Green Version]
- Lira-Amaya, J.J.; Rojas-Martinez, C.; Alvarez, A.; Pelaezflores, A.; Martinez-Ibañez, F.; Rosa, D.P.; Figueroamillan, J.V. First Molecular Detection of Babesia Canis Vogeli in Dogs and Rhipicephalus Sanguineus from Mexico. Arch. Palliat. Care 2017, 2, 2–5. [Google Scholar]
- Birkenheuer, A.J.; Correa, M.T.; Levy, M.G.; Breitschwerdt, E.B. Geographic Distribution of Babesiosis among Dogs in the United States and Association with Dog Bites: 150 Cases (2000–2003). J. Am. Vet. Med. Assoc. 2005, 227, 942–947. [Google Scholar] [CrossRef]
- Stayton, E.M.; Lineberry, M.; Thomas, J.; Bass, T.; Allen, K.; Chandrashekar, R.; Yost, G.; Reichard, M.; Miller, C. Emergence of Babesia Conradae Infection in Coyote-Hunting Greyhounds in Oklahoma, USA. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Allison, R.W.; Yeagley, T.J.; Levis, K.; Reichard, M.V. Babesia Canis Rossi Infection in a Texas Dog. Vet. Clin. Pathol. 2011, 40, 345–350. [Google Scholar] [CrossRef]
- Medrano-Bugarini, R.A.; Figueroa-Millán, J.V.; Rivera-Chavira, B.E.; Lira-Amaya, J.J.; Rodríguez-Alarcón, C.A.; Beristain-Ruiz, D.M.; Adame-Gallegos, J.R. Detection of Theileria Equi, Babesia Caballi, and Anaplasma Phagocytophilum DNA in Soft Ticks and Horses at Ciudad Juarez, Mexico. Southwest. Entomol. 2019, 44, 647–658. [Google Scholar] [CrossRef]
- Cantú-Martínez, M.A.; Segura-Correa, J.C.; Silva-Páez, M.L.; Avalos-Ramírez, R.; Wagner, G.G. Prevalence of Antibodies to Theileria Equi and Babesia Caballi in Horses from Northeastern Mexico. J. Parasitol. 2012, 98, 869–870. [Google Scholar] [CrossRef]
- Scott, J.D.; Pascoe, E.L.; Sajid, M.S.; Foley, J.E. Detection of Babesia Odocoilei in Ixodes Scapularis Ticks Collected in Southern Ontario, Canada. Pathogens 2021, 10, 327. [Google Scholar] [CrossRef] [PubMed]
- Olafson, P.U.; Thomas, D.B.; May, M.A.; Buckmeier, B.G.; Duhaime, R.A. Tick Vector and Disease Pathogen Surveillance of Nilgai Antelope (Boselaphus Tragocamelus) in Southeastern Texas, USA. J. Wildl. Dis. 2018, 54, 734–744. [Google Scholar] [CrossRef]
- Ueti, M.W.; Olafson, P.U.; Freeman, J.M.; Johnson, W.C.; Scoles, G.A. A Virulent Babesia Bovis Strain Failed to Infect White-Tailed Deer (Odocoileus Virginianus). PLoS ONE 2015, 10, e0131018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vishwakarma, P.; Nandini, M.K. Overview of Canine Babesiosis. In Veterinary Medicine and Pharmaceuticals; IntechOpen: London, UK, 2019; p. 109. [Google Scholar]
- Boozer, A.L.; Macintire, D.K. Canine Babesiosis. Vet. Clin. Small Anim. Pract. 2003, 33, 885–904. [Google Scholar] [CrossRef]
- Solano-Gallego, L.; Sainz, Á.; Roura, X.; Estrada-Peña, A.; Miró, G. A Review of Canine Babesiosis: The European Perspective. Parasites Vectors 2016, 9, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emerson, H.R.; Wright, W.T. The Isolation of a Babesia in White-Tailed Deer. Bull. Wildl. Dis. Assoc. 1968, 4, 142–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirosh-Levy, S.; Gottlieb, Y.; Fry, L.M.; Knowles, D.P.; Steinman, A. Twenty Years of Equine Piroplasmosis Research: Global Distribution, Molecular Diagnosis, and Phylogeny. Pathogens 2020, 9, 926. [Google Scholar] [CrossRef]
- Wise, L.N.; Kappmeyer, L.S.; Mealey, R.H.; Knowles, D.P. Review of Equine Piroplasmosis. J. Vet. Intern. Med. 2013, 27, 1334–1346. [Google Scholar] [CrossRef]
- Onyiche, T.E.; Suganuma, K.; Igarashi, I.; Yokoyama, N.; Xuan, X.; Thekisoe, O. A Review on Equine Piroplasmosis: Epidemiology, Vector Ecology, Risk Factors, Host Immunity, Diagnosis and Control. Int. J. Environ. Res. Public Health 2019, 16, 1736. [Google Scholar] [CrossRef] [Green Version]
- Rothschild, C.M. Equine Piroplasmosis. J. Equine Vet. Sci. 2013, 33, 497–508. [Google Scholar] [CrossRef]
- Sears, K.P.; Kappmeyer, L.S.; Wise, L.N.; Silva, M.; Ueti, M.W.; White, S.; Reif, K.E.; Knowles, D.P. Infection Dynamics of Theileria Equi and Theileria Haneyi, a Newly Discovered Apicomplexan of the Horse. Vet. Parasitol. 2019, 271, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Sumbria, D.; Moudgil, A.D.; Singla, L.D. Equine Piroplasmosis: Current Status. Veterinaria 2014, 1, 9–14. [Google Scholar]
- Laus, F.; Spaterna, A.; Faillace, V.; Veronesi, F.; Ravagnan, S.; Beribé, F.; Cerquetella, M.; Meligrana, M.; Tesei, B. Clinical Investigation on Theileria Equi and Babesia Caballi Infections in Italian Donkeys. BMC Vet. Res. 2015, 11, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kouam, M.K.; Kantzoura, V.; Gajadhar, A.A.; Theis, J.H.; Papadopoulos, E.; Theodoropoulos, G. Seroprevalence of Equine Piroplasms and Host-Related Factors Associated with Infection in Greece. Vet. Parasitol. 2010, 169, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, D.F.; Wright, I.G.; Frerichs, W.M.; Groenendyk, S.; O’Sullivan, B.M.; Roberts, M.C.; Waddell, A.H. The Identification of Babesia Equi in Australia. Aust. Vet. J. 1977, 53, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Zobba, R.; Ardu, M.; Niccolini, S.; Chessa, B.; Manna, L.; Cocco, R.; Parpaglia, M.L.P. Clinical and Laboratory Findings in Equine Piroplasmosis. J. Equine Vet. Sci. 2008, 28, 301–308. [Google Scholar] [CrossRef]
- Mahmoud, M.S.; El-Ezz, N.T.A.; Abdel-Shafy, S.; Nassar, S.A.; El Namaky, A.H.; Khalil, W.K.; Knowles, D.; Kappmeyer, L.; Silva, M.G.; Suarez, C.E. Assessment of Theileria Equi and Babesia Caballi Infections in Equine Populations in Egypt by Molecular, Serological and Hematological Approaches. Parasites Vectors 2016, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Friedhoff, K.T.; Tenter, A.M.; Müller, I. Haemoparasites of Equines: Impact on International Trade of Horses. Rev. Sci. Et Tech. (Int. Off. Epizoot.) 1990, 9, 1187–1194. [Google Scholar]
- Brüning, A. Equine Piroplasmosis an Update on Diagnosis, Treatment and Prevention. Br. Vet. J. 1996, 152, 139–151. [Google Scholar] [CrossRef]
- Awinda, P.O. Serum Antibodies from Babesia Caballi Seropositive Horses in the United States Demonstrate a Protein Recognition Pattern That Is Not Consistent with Infection. Ph.D. Thesis, Washington State University, Washington, DC, USA, 2013. [Google Scholar]
- Alhassan, A.; Govind, Y.; Tam, N.T.; Thekisoe, O.M.; Yokoyama, N.; Inoue, N.; Igarashi, I. Comparative Evaluation of the Sensitivity of LAMP, PCR and in Vitro Culture Methods for the Diagnosis of Equine Piroplasmosis. Parasitol. Res. 2007, 100, 1165–1168. [Google Scholar] [CrossRef]
- Hirato, K.; Ninomiya, M.; Uwano, Y.; Kuth, T. Studies on the Complement Fixation Reaction for Equine Piroplasmosis. Jpn. J. Vet. Sci. 1945, 7, 197–205. [Google Scholar] [CrossRef]
- Knowles, D.P., Jr.; Kappmeyer, L.S.; Stiller, D.; Hennager, S.G.; Perryman, L.E. Antibody to a Recombinant Merozoite Protein Epitope Identifies Horses Infected with Babesia Equi. J. Clin. Microbiol. 1992, 30, 3122–3126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayala-Valdovinos, M.A.; Lemus-Flores, C.; Galindo-García, J.; Bañuelos-Pineda, J.; Rodríguez-Carpena, J.G.; Sánchez-Chiprés, D.; Duifhuis-Rivera, T. Diagnosis and Prevalence of Theileria Equi Horses in Western Mexico by Nested PCR. Parasitol. Int. 2017, 66, 821–824. [Google Scholar] [CrossRef] [PubMed]
- Romero-Salas, D.; Solis-Cortés, M.; Zazueta-Islas, H.M.; Flores-Vásquez, F.; Cruz-Romero, A.; Aguilar-Domínguez, M.; Salguero-Romero, J.L.; de León, A.P.; Fernández-Figueroa, E.A.; Lammoglia-Villagómez, M.Á. Molecular Detection of Theileria Equi in Horses from Veracruz, Mexico. Ticks Tick-Borne Dis. 2021, 12, 101671. [Google Scholar] [CrossRef] [PubMed]
- Bryant, J.E.; Anderson, J.B.; Willers, K.H. Control of Equine Piroplasmosis in Florida. J. Am. Vet. Med. Assoc. 1969, 154, 1034–1036. [Google Scholar]
- Stockham, S.L.; Kjemtrup, A.M.; Conrad, P.A.; Schmidt, D.A.; Scott, M.A.; Robinson, T.W.; Tyler, J.W.; Johnson, G.C.; Carson, C.A.; Cuddihee, P. Theileriosis in a Missouri Beef Herd Caused by Theileria Buffeli: Case Report, Herd Investigation, Ultrastructure, Phylogenetic Analysis, and Experimental Transmission. Vet. Pathol. 2000, 37, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Splitter, E.J. Theileria Mutans Associated with Bovine Anaplasmosis in the United States. J. Am. Vet. Med. Assoc. 1950, 117, 134–135. [Google Scholar]
- Cossio-Bayugar, R.; Pillars, R.; Schlater, J.; Holman, P.J. Theileria Buffeli Infection of a Michigan Cow Confirmed by Small Subunit Ribosomal RNA Gene Analysis. Vet. Parasitol. 2002, 105, 105–110. [Google Scholar] [CrossRef]
- Robinson, R.M.; Kuttler, K.L.; Thomas, J.W.; Marburger, R.G. Theileriasis in Texas White-Tailed Deer. J. Wildl. Manag. 1967, 31, 455–459. [Google Scholar] [CrossRef]
- Chae, J.; Waghela, S.D.; Craig, T.M.; Kocan, A.A.; Wagner, G.G.; Holman, P.J. Two Theileria Cervi SSU RRNA Gene Sequence Types Found in Isolates from White-Tailed Deer and Elk in North America. J. Wildl. Dis. 1999, 35, 458–465. [Google Scholar] [CrossRef] [Green Version]
- Yabsley, M.J.; Quick, T.C.; Little, S.E. Theileriosis in a White-Tailed Deer (Odocoileus Virginianus) Fawn. J. Wildl. Dis. 2005, 41, 806–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cauvin, A.; Hood, K.; Shuman, R.; Orange, J.; Blackburn, J.K.; Sayler, K.A.; Wisely, S.M. The Impact of Vector Control on the Prevalence of Theileria Cervi in Farmed Florida White-Tailed Deer, Odocoileus Virginianus. Parasites Vectors 2019, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kjemtrup, A.M.; Robinson, T.; Conrad, P.A. Description and Epidemiology of Theileria Youngi n. Sp. from a Northern Californian Dusky-Footed Woodrat (Neotoma Fuscipes) Population. J. Parasitol. 2001, 87, 373–378. [Google Scholar] [CrossRef]
- Camacho, A.T.; Guitián, F.J.; Pallas, E.; Gestal, J.J.; Olmeda, A.S.; Goethert, H.K.; Telford, S.R. Infection of Dogs in North-West Spain with a Babesia Microti-like Agent. Vet. Rec. 2001, 149, 552–555. [Google Scholar] [CrossRef]
- Uilenberg, G.; Perie, N.M.; Spanjer, A.A.M.; Franssen, F.F.J. Theileria Orientalis, a Cosmopolitan Blood Parasite of Cattle: Demonstration of the Schizont Stage. Res. Vet. Sci. 1985, 38, 352–360. [Google Scholar] [CrossRef]
- McKeever, D.J. Bovine Immunity–a Driver for Diversity in Theileria Parasites? Trends Parasitol. 2009, 25, 269–276. [Google Scholar] [CrossRef]
- Mans, B.J.; Pienaar, R.; Latif, A.A. A Review of Theileria Diagnostics and Epidemiology. Int. J. Parasitol. Parasites Wildl. 2015, 4, 104–118. [Google Scholar] [CrossRef] [Green Version]
- Irvin, A.D. Characterization of Species and Strains of Theileria. Adv. Parasitol. 1987, 26, 145–197. [Google Scholar]
- Bishop, R.; Musoke, A.; Morzaria, S.; Gardner, M.; Nene, V. Theileria: Intracellular Protozoan Parasites of Wild and Domestic Ruminants Transmitted by Ixodid Ticks. Parasitology 2004, 129, S271–S283. [Google Scholar] [CrossRef]
- Aktas, M.; Altay, K.; Dumanli, N. A Molecular Survey of Bovine Theileria Parasites among Apparently Healthy Cattle and with a Note on the Distribution of Ticks in Eastern Turkey. Vet. Parasitol. 2006, 138, 179–185. [Google Scholar] [CrossRef]
- Kamau, J.; de Vos, A.J.; Playford, M.; Salim, B.; Kinyanjui, P.; Sugimoto, C. Emergence of New Types of Theileria Orientalis in Australian Cattle and Possible Cause of Theileriosis Outbreaks. Parasites Vectors 2011, 4, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perera, P.K.; Gasser, R.B.; Firestone, S.M.; Anderson, G.A.; Malmo, J.; Davis, G.; Beggs, D.S.; Jabbar, A. Oriental Theileriosis in Dairy Cows Causes a Significant Milk Production Loss. Parasites Vectors 2014, 7, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, K.E.; Forsyth, S.F.; Vaatstra, B.L.; McFadden, A.M.J.; Pulford, D.J.; Govindaraju, K.; Pomroy, W.E. Clinical Haematology and Biochemistry Profiles of Cattle Naturally Infected with Theileria Orientalis Ikeda Type in New Zealand. N. Z. Vet. J. 2018, 66, 21–29. [Google Scholar] [CrossRef] [PubMed]
- McFadden, A.M.J.; Hart, M.; Bueno, I.M.; Ha, H.J.; Heath, A.C.G.; Pulford, D.J. Monitoring Theileria Orientalis (Ikeda)-Associated Bovine Anaemia in Affected Cattle over Time. Vet. Parasitol. 2017, 245, 29–33. [Google Scholar] [CrossRef]
- Swilks, E.; Fell, S.A.; Hammer, J.F.; Sales, N.; Krebs, G.L.; Jenkins, C. Transplacental Transmission of Theileria Orientalis Occurs at a Low Rate in Field-Affected Cattle: Infection in Utero Does Not Appear to Be a Major Cause of Abortion. Parasites Vectors 2017, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Gebrekidan, H.; Perera, P.K.; Ghafar, A.; Abbas, T.; Gasser, R.B.; Jabbar, A. An Appraisal of Oriental Theileriosis and the Theileria Orientalis Complex, with an Emphasis on Diagnosis and Genetic Characterisation. Parasitol. Res. 2020, 119, 11–22. [Google Scholar] [CrossRef]
- Watts, J.G.; Playford, M.C.; Hickey, K.L. Theileria Orientalis: A Review. N. Z. Vet. J. 2016, 64, 3–9. [Google Scholar] [CrossRef]
- Charaya, G.; Rakha, N.K.; Maan, S.; Kumar, A.; Kumar, T.; Jhambh, R. Comparative Evaluation of Polymerase Chain Reaction Assay with Microscopy for Detection of Asymptomatic Carrier State of Theileriosis in a Herd of Crossbred Cattle. Vet. World 2016, 9, 1039. [Google Scholar] [CrossRef] [Green Version]





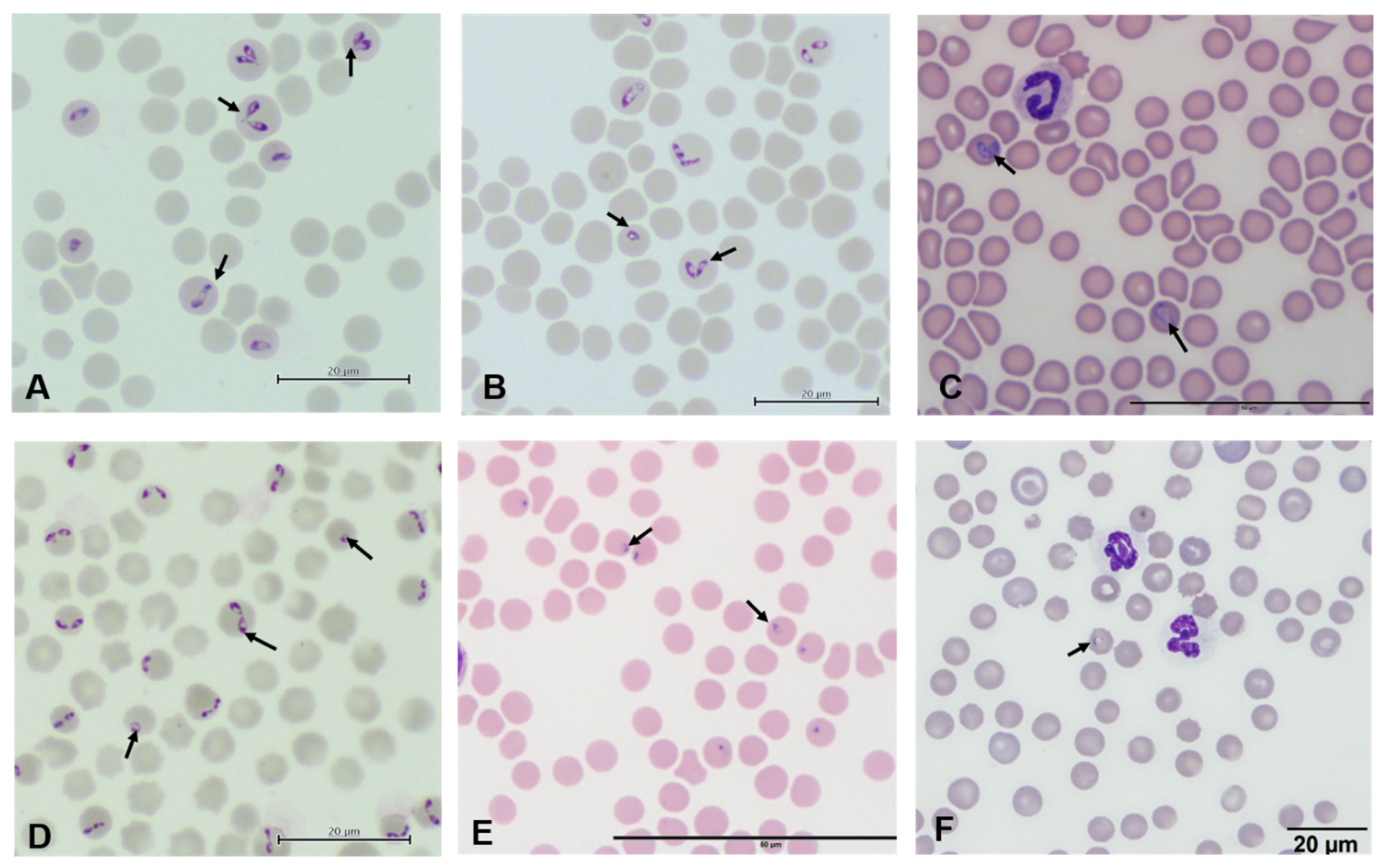{kind=link}
{kind=link}
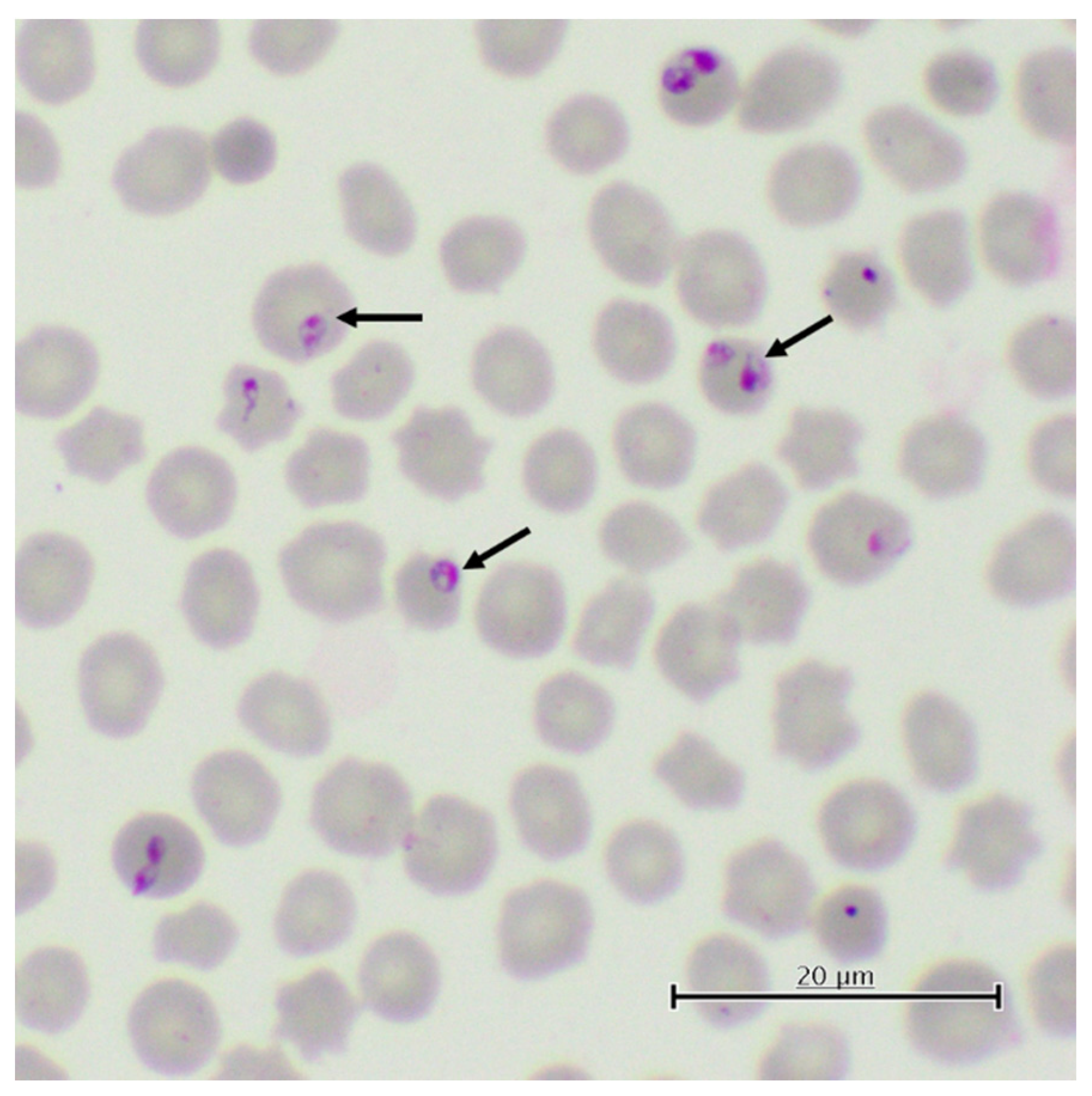{kind=link}
{kind=link}
| Pathogen | Tick Vectors | Affected Hosts | References |
|---|---|---|---|
| Babesia microti | Ixodes scapularis | Humans | [7,8,9] |
| B. duncani | Dermacentor albipictus | Humans | [10] |
| B. divergens-like | ND | Humans | [11] |
| B. bigemina | Rhipicephalus microplus, R. annulatus | Cattle | [12,13,14] |
| B. bovis | R. microplus, R. annulatus | Cattle | [12,13,14] |
| B. gibsoni | R. sanguineus * | Dogs | [15] |
| B. vogeli | R. sanguineus | Dogs | [15] |
| B. conradae | ND | Dogs | [16] |
| B. vulpes | ND | Dogs | [17] |
| B. caballi | D. albipictus, D. variabilis, D. nitens | Equids | [18,19] |
| B. odocoilei | I. scapularis | White-tailed deer, reindeer, and elk | [20] |
| Theileria equi | D. nitens; Amblyomma mixtum | Horses | [18] |
| T. haneyi | ND | Horses | [21] |
| T. orientalis genotype ikeda | Haemaphysalis longicornis | Cattle | [22] |
| T. cervi | A. americanum | White-tailed deer, elk, and mule deer | [23,24] |
| Species | Samples | Number of Confirmed Cases | Year | Confirmation Method | Geographical Location | References |
|---|---|---|---|---|---|---|
| B. microti | Blood, serum | 7612 cases | 2011–2015 | Microscopy, PCR, Animal inoculation, and IFAT | New York, US * Massachusetts, US Connecticut, US New Jersey, US Rhode Island, US Wisconsin, US Minnesota, US | [40] |
| B. microti | Blood, serum | 14,159 | 2011–2018 | Microscopy, PCR, Animal inoculation, and IFAT | New York, US * Massachusetts, US Connecticut, US New Jersey, US Rhode Island, US Wisconsin, US New York City, US Maine, US Minnesota, US | [6] |
| B. microti | Blood | 4 | 2015 | PCR for 18S rRNA | Yucatan, MX | [25] |
| B. microti | Blood | 1 | 2013 | PCR for 18S rRNA | Manitoba; Can | [4] |
| B. duncani | Serum, blood | 14 | 1966–2009 | IFAT and PCR for ITS | California, US Washington, US Oregon, US | [10] |
| Species | Sample/Host | Host | Method | Prevalence (%) | Location | References |
|---|---|---|---|---|---|---|
| B. bovis | Serum | Cattle | IFAT | 50 | Nuevo Leon, MX | [50] |
| Blood | Cattle | Microscopy | 2.78 | Yucatan, MX | [52] | |
| Serum | Cattle | Indirect ELISA | 73.8 | Yucatan, MX | [49] | |
| Cow Serum | Cattle | IFAT | 98 | Veracruz, MX | [51] | |
| Serum | Water buffalo | IFAT | 71.4 | Veracruz, MX | [51] | |
| Blood | Cattle | Nested PCR for CYTb | 82.3 | Veracruz, MX | [51] | |
| Blood | Water buffalo | Nested PCR for CYTb | 16.2 | Veracruz, MX | [51] | |
| Blood | WTD | Nested PCR | 1.7 | Northeast | [53] | |
| Blood | WTD | IFAT | 16 & 4 | La Salle and Webb counties, TX, US | [54] | |
| Blood | WTD | PCR for 18S rDNA | 12 | Tom Green Co, TX, US | [55] | |
| Serum | WTD | IFAT | 59.9 | Northeast, MX | [53] | |
| Blood | Nilgai | PCR for Rap-1 | 5 | Coahuila, MX | [56] | |
| Blood | Fallow deer | PCR for Rap-1 | 9.5 | Tamaulipas, MX | [57] | |
| B. bigemina | Serum | Cattle | IFAT | 56 | Nuevo Leon, MX | [50] |
| Blood | Cattle | Microscopy | 1.2 | Yucatan, MX | [52] | |
| Serum | Water buffalo | IFAT | 85 | Veracruz, MX | [51] | |
| Serum | Cattle | IFAT | 100 | Veracruz, MX | [51] | |
| Blood | Cattle | Nested PCR for CYTb | 94.1 | Veracruz, MX | [51] | |
| Blood | Water buffalo | Nested PCR for CYTb | 24 | Veracruz, MX | [51] | |
| Blood | Cattle | Microscopy | 2.5 | Yucatan, MX | [52] | |
| Serum | Cattle | IFAT | 57 | Southeast MX | [48] | |
| Blood | Nilgai | PCR for Rap-1 | 25 | Coahuila, MX | [56] | |
| Blood | WTD | Nested PCR | 4.2 | Northeast, MX | [53] | |
| Serum | WTD | IFAT | 5.4 | Northeast, MX | [53] | |
| Blood | Fallow deer | PCR for Rap-1 | 4.7 | Tamaulipas, MX | [57] | |
| B. vogeli | Blood | Dogs | Microscopy | NA, Detection | Veracruz, MX * | [44] |
| Blood | Dogs | PCR for 18S rRNA | 6.6 | Morelos, MX | [58] | |
| R. sanguineus | Dogs | PCR 18S rRNA | 5.5 | Morelos, MX | [58] | |
| Blood | Dogs | Microscopy | 3.9 | Yucatan, MX * | [52] | |
| Blood | Dogs | PCR | 6.9 | 29 states, US | [59] | |
| Blood | Dogs | PCR for 18S rRNA | 0.31 & 1.7 | North Carolina | [17] | |
| B.gibsoni | Blood | Dogs | PCR | 91 | 29 states, US, and Ontario, CA | [59] |
| B. conradae | Blood | Dogs | Microscopy and PCR for 18S rRNA and ITS-2 | NA (Isolation and detection) | California, US | [16] |
| Blood | Dogs | PCR for 18S rRNA and ITS-2 region | 52.7 | South-central California, US | [28] | |
| Blood | Dogs | 18S rRNA | 37.5 | Oklahoma, US | [60] | |
| B. rossi | Blood | Dogs | PCR for 18S rRNA | NA Only detection | Texas, US | [61] |
| B. vulpes | Blood | Dogs | PCR for 18S rRNA | 0.20 | North Carolina | [17] |
| B. caballi | Blood | Horses | Microscopy | NA | Veracruz, MX | [19] |
| Blood | Horses | PCR, for BC48 | 20.8 | Juarez, MX | [62] | |
| Otobius megnini | Horses | PCR | 5.9 | Juarez, MX | [62] | |
| Sera | Horses | IFAT | 27.4 | Nuevo Leon, MX | [63] | |
| B. odocoilei | Blood | White-tailed deer | Microscopy | 2 | Texas | [32] |
| Blood | Reindeer and elk | PCR | - | Quebec, Ontario, and Manitoba, CA | [20] | |
| Ixodes scapularis | Dogs, cats, and questing ticks | PCR | 71 & 12.5 | Southern Ontario, CA | [64] | |
| Blood | Free-ranging desert bighorn sheep (Ovis canadensis nelsoni) | Microscopy and PCR for SSU rRNA gene | - | California, US | [29] | |
| Blood | Captive musk oxen (Ovibos moschatus) | Microscopy and PCR for SSU rRNA gene | 2 cases | Minnessota, US | [29] | |
| Sera | Elk | IFAT | 100, 100, & 26 in 3 farms | New Hampshire, US | [29] | |
| Blood | Elk | PCR | Detection of one case | New Hampshire, US | [29] | |
| Blood | Reindeer | PCR | Detection of three cases | Pennsylvania and New York, US | [29] |
| Species | Sample | Host | Method | Prevalence (%) | Location | References |
|---|---|---|---|---|---|---|
| T. equi | blood | Horse | Microscopy and | Veracruz, MX | [19] | |
| Nested PCR for EMA-1 | 6.9 | |||||
| Soft ticks | Nested PCR for EMA-1 | 5.9 | Juarez, MX | [62] | ||
| Blood | Nested PCR for EMA-1 | 19.7 | Jalisco, MX | [88] | ||
| Blood | Horse | Microscopy | 3.79 | Yucatan, MX | [52] | |
| Serum | Horse | IFAT | 45.2 | Nuevo Leon, MX | [63] | |
| Serum | Horse | CFT, IFAT, and cELISA | 9.5 | Florida, US | [30] | |
| serum | Horse | c-ELISA | 81.1 | Southern Texas, US | [31] | |
| T. haneyi | blood | Horse | PCR for 18S rDNA and nPCR for EMA | First report | Southern Texas, US | [21] |
| T. orientalis genotype ikeda | blood | Cattle (beef cattle) | PCR for SSU rRNA | Detection in 3 beef cattle | Virginia, US | [22] |
| T.orientalis genotype buffeli | blood | Cattle (beef cattle) | Microscopy IFAT | 39 35 | Missouri, US (1 herd) | [91] |
| blood | Cattle (Angus) | PCR for SSU rRNA | Detected in a cow | North Carolina, US | [92] | |
| Blood and ticks | Cattle (mixed breed cow) | PCR for SSU rRNA | Detection in 3 cows and A.americanun and D. variabilis ticks | Missouri, US | [24] | |
| blood | Cattle (beef cattle) | PCR for SSU rRNA | Detection | Michigan, US | [93] | |
| T. cervi | Blood | White-tailed deer * | Microscopy | 72 | Texas, US | [32] |
| Blood | White-tailed deer | Microscopy | 57 | Texas, US | [94] | |
| Blood | Free-ranging elk | PCR for 18S rRNA | Case report | Canada ** | [95] | |
| Blood | Free-ranging elk | PCR for 18S rRNA | Case report | Oklahoma, US | [95] | |
| Blood | White-tailed deer | PCR for 18S rRNA | Case report | Texas, Oklahoma, and Missouri, US | [95,96] | |
| Blood, liver, lymph nodes, and spleen | Mule deer | Microscopy and PCR for 18S sRNA | Case report | Oklahoma, US | [23] | |
| Blood | White-tailed deer | PCR for 18S rRNA | 97.6 in Wild animals 40.4 in farmed animals | Florida, US | [97] | |
| Blood | White-tailed deer | Microscopy and PCR for 18S rRNA | Detection in 3 animals | Northeastern, MX | [33] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almazán, C.; Scimeca, R.C.; Reichard, M.V.; Mosqueda, J. Babesiosis and Theileriosis in North America. Pathogens 2022, 11, 168. https://doi.org/10.3390/pathogens11020168
Almazán C, Scimeca RC, Reichard MV, Mosqueda J. Babesiosis and Theileriosis in North America. Pathogens. 2022; 11(2):168. https://doi.org/10.3390/pathogens11020168
Chicago/Turabian StyleAlmazán, Consuelo, Ruth C. Scimeca, Mason V. Reichard, and Juan Mosqueda. 2022. "Babesiosis and Theileriosis in North America" Pathogens 11, no. 2: 168. https://doi.org/10.3390/pathogens11020168