
Differential Expression of Immune Genes in the Rhipicephalus microplus Gut in Response to Theileria equi Infection

 , , , ,
, , , ,  and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
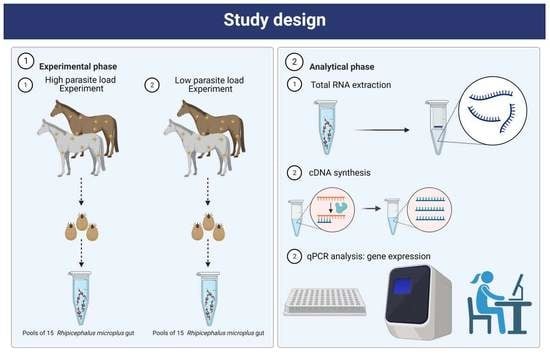
2. Materials and Methods
2.1. Consent of the Ethics Committee
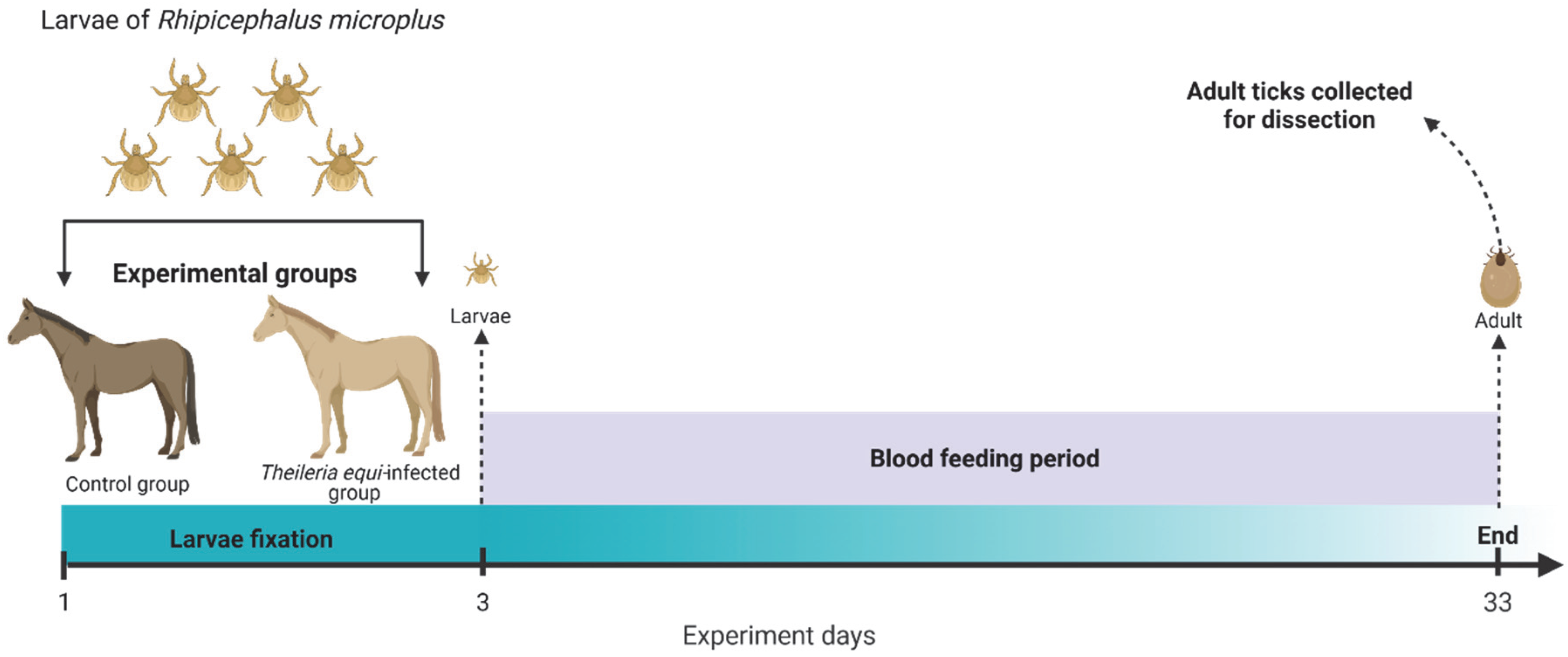
2.2. Experimental Animals
2.3. Experimental Manipulation of Theileria equi Parasitemia
2.4. DNA Extraction
2.5. Tick Infestation and Collection
2.6. Tick Dissection
2.7. RNA Extraction and cDNA Synthesis
2.8. Detection and Quantification of Theileria equi in Horses and Ticks
2.9. Gene Expression Assays
2.10. Statistical Analysis
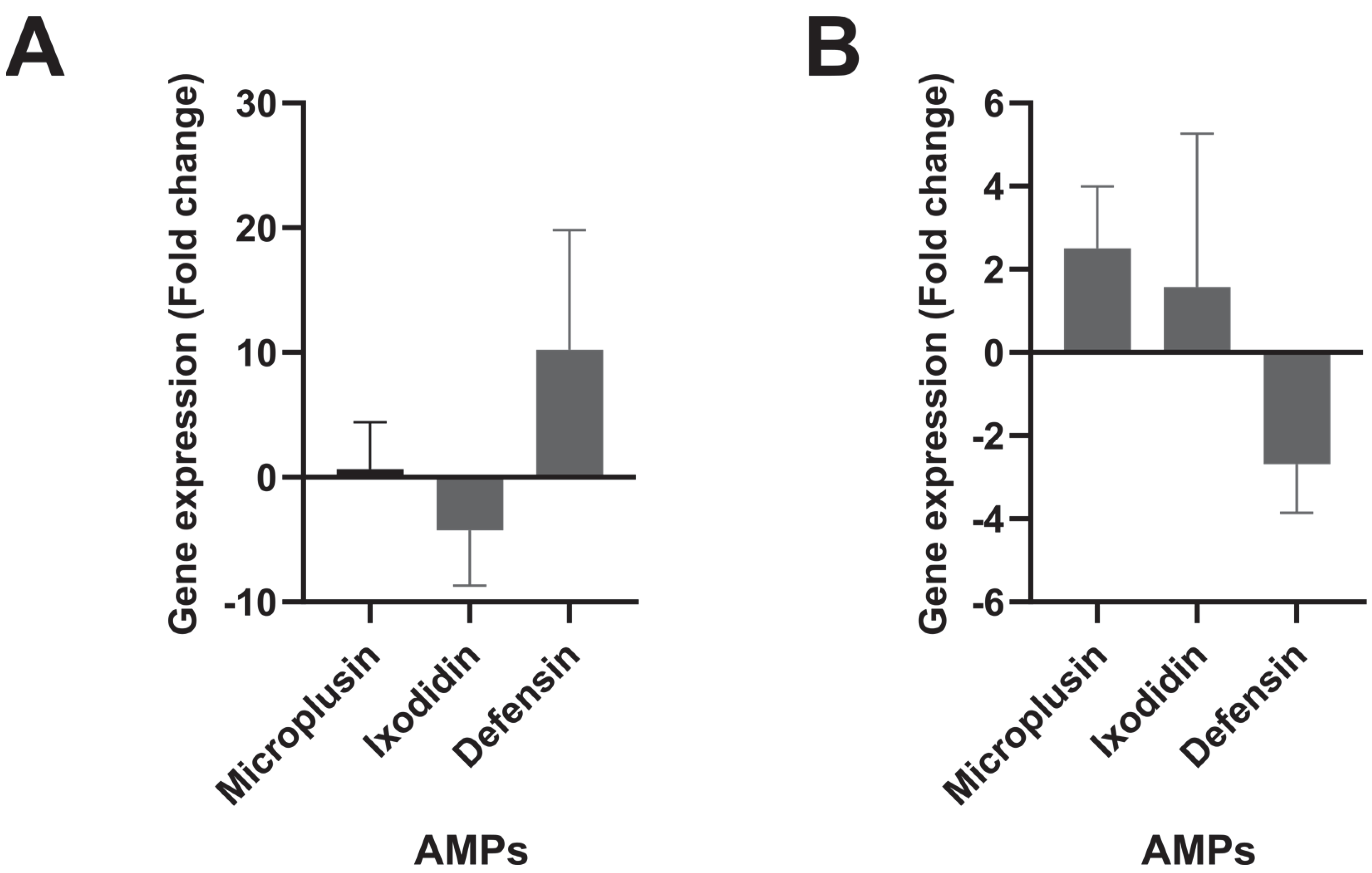
3. Results
3.1. Differential Theileria equi Parasitemia in Infected Horses and Collected Ticks
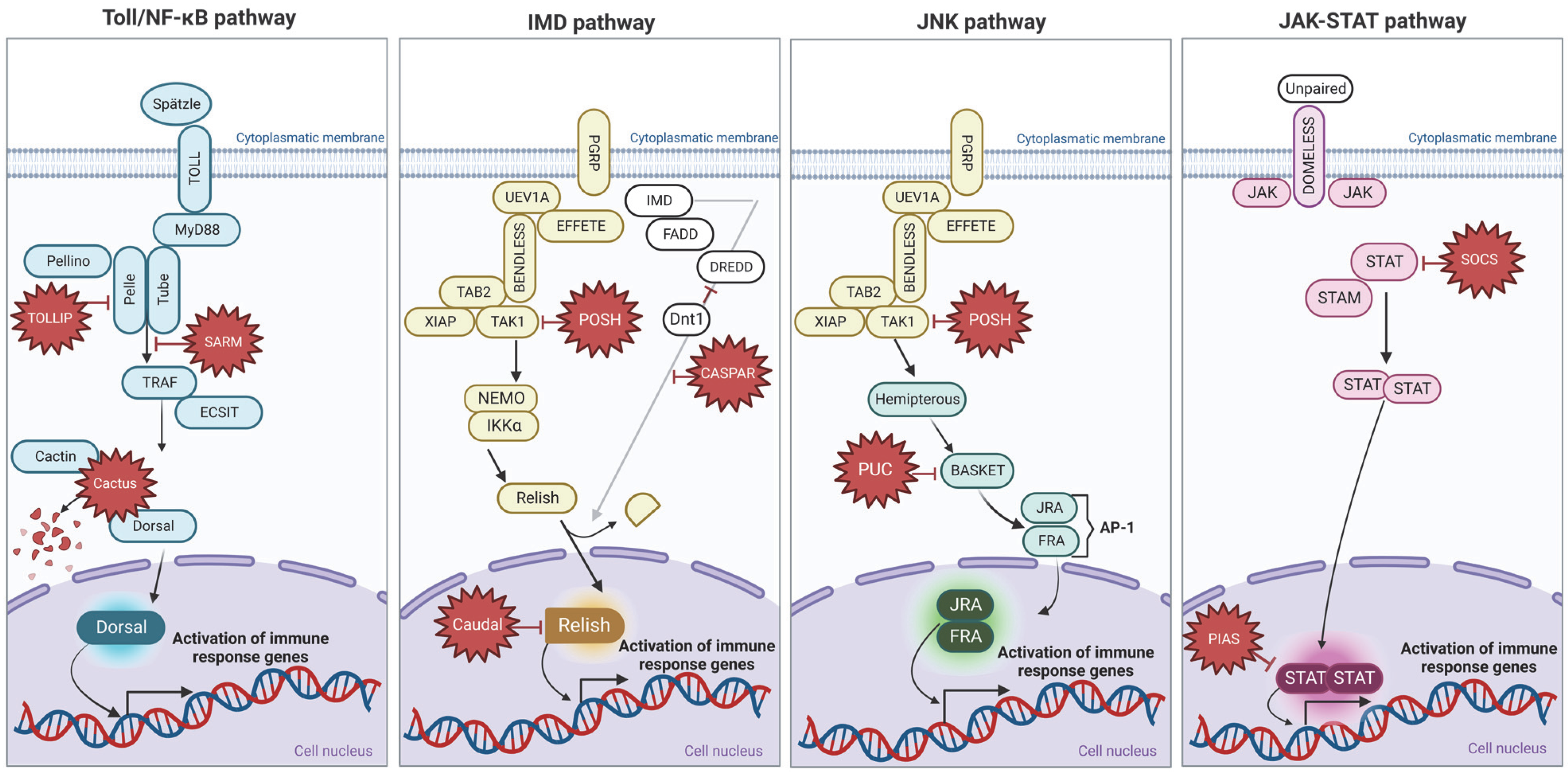
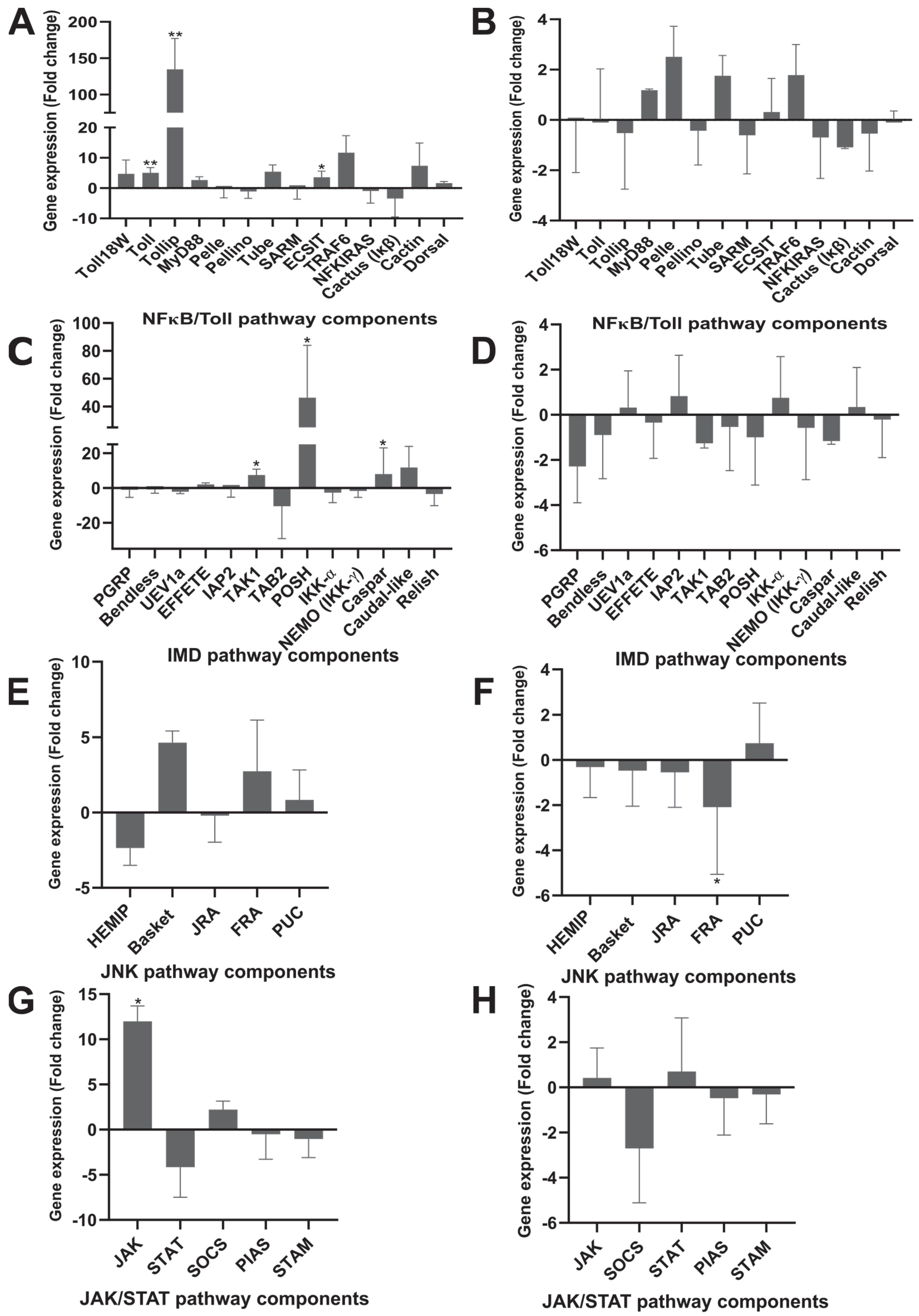
3.2. Toll Pathway
3.3. IMD Pathway
3.4. JNK Pathway
3.5. JAK/STAT Signaling Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tirosh-Levy, S.; Gottlieb, Y.; Fry, L.M.; Knowles, D.P.; Steinman, A. Twenty Years of Equine Piroplasmosis Research: Global Distribution, Molecular Diagnosis, and Phylogeny. Pathogens 2020, 9, 926. [Google Scholar] [CrossRef] [PubMed]
- Sears, K.; Knowles, D.; Dinkel, K.; Mshelia, P.W.; Onzere, C.; Silva, M.; Fry, L. Imidocarb dipropionate lacks efficacy against Theileria haneyi and fails to consistently clear Theileria equi in horses co-infected with T. haneyi. Pathogens 2020, 9, 1035. [Google Scholar] [CrossRef] [PubMed]
- Friedhoff, K.T.; Tenter, A.M.; Müller, I. Haemoparasites of Equines: Impact on International Trade of Horses. Rev. Sci. Et Tech. 1990, 9, 1187–1194. [Google Scholar]
- Ikadai, H.; Sasaki, M.; Ishida, H.; Matsuu, A.; Igarashi, I.; Fujisaki, K.; Oyamada, T. Molecular evidence of Babesia equi transmission in Haemaphysalis Longicornis. Am. J. Trop. Med. Hyg. 2007, 76, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Scoles, G.A.; Ueti, M.W. Vector Ecology of Equine Piroplasmosis. Annu. Rev. Entomol. 2015, 60, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, A.M.; Lima, J.D.; Ribeiro, M.F.B.; Camargos, E.R.S.; Bozzi, I.A. Ultrastructure of sporogony in Babesia equi in salivary glands of adult female Boophilus microplus ticks. Parasitol. Res. 1997, 84, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, A.M.; Lima, J.D.; Ribeiro, M.F.B. Sporogony and experimental transmission of Babesia equi by Boophilus microplus. Parasitol. Res. 1998, 84, 323–327. [Google Scholar] [CrossRef]
- Peckle, M.; Santos, H.; Pires, M.; Silva, C.; Costa, R.; Vitari, G.; Camilo, T.; Meireles, N.; Paulino, P.; Massard, C. Dynamics of Theileria equi Infection in Rhipicephalus (Boophilus) microplus during the Parasitic Phase in a Chronically Infected Horse. Pathogens 2022, 11, 525. [Google Scholar] [CrossRef]
- Kopáček, P.; Hajdušek, O.; Burešová, V.; Daffre, S. Tick innate immunity. In Invertebrate Immunity. Advances in Experimental Medicine and Biology; Söderhäll, K., Ed.; Springer: Boston, MA, USA, 2010; Volume 708. [Google Scholar] [CrossRef]
- Fogaca, A.C.; Sousa, G.; Pavanelo, D.B.; Esteves, E.; Martins, L.A.; Urbanová, V.; Kopáček, P.; Daffre, S. Tick immune system: What is known, the interconnections, the gaps, and the challenges. Front. Immunol. 2021, 12, 628054. [Google Scholar] [CrossRef]
- Shaw, D.K.; Wang, X.; Brown, L.J.; Chavez, A.S.; Reif, K.E.; Smith, A.A.; Scott, A.J.; McClure, E.E.; Boradia, V.M.; Hammond, H.L.; et al. Infection-derived lipids elicit an immune deficiency circuit in arthropods. Nat. Commun. 2017, 8, 14401. [Google Scholar] [CrossRef]
- Aguilar-Díaz, H.; Quiroz-Castañeda, R.E.; Salazar-Morales, K.; Cossío-Bayúgar, R.; Miranda-Miranda, E. Tick Immunobiology and Extracellular Traps: An Integrative Vision to Control of Vectors. Pathogens 2021, 10, 1511. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.D.; Capelli-Peixoto, J.; Mesquita, R.D.; Kalil, S.P.; Pohl, P.C.; Braz, G.R.; Daffre, S. Exploring the immune signaling pathway-related genes of the cattle tick Rhipicephalus microplus: From molecular characterization to transcriptional profile upon microbial challenge. Dev. Comp. Immunol. 2016, 59, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Capelli-Peixoto, J.; Carvalho, D.D.; Johnson, W.C.; Scoles, G.A.; Fogaça, A.C.; Daffre, S.; Ueti, M.W. The transcription factor Relish controls Anaplasma marginale infection in the bovine tick Rhipicephalus microplus. Dev. Comp. Immunol. 2017, 74, 32–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Dai, J.; Zhao, Y.O.; Narasimhan, S.; Yang, Y.; Zhang, L.; Fikrig, E. Ixodes scapularis JAK-STAT pathway regulates tick antimicrobial peptides, thereby controlling the agent of human granulocytic anaplasmosis. J. Infect. Dis. 2012, 206, 1233–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldani, C.D.; Machado, R.Z.; Botteon, P.D.T.L.; Takakura, F.S.; Massard, C.L. An enzyme-linked immunosorbent assay for the detection of IgG antibodies against Babesia equi in horses. Ciência Rural 2004, 34, 1525–1529. [Google Scholar] [CrossRef]
- Paulino, P.; Vitari, G.; Rezende, A.; Couto, J.; Antunes, S.; Domingos, A.; Peckle, M.; Massard, C.; Araújo, F.; Santos, H. Characterization of the Rhipicephalus (Boophilus) microplus sialotranscriptome profile in response to Theileria equi infection. Pathogens 2021, 10, 167. [Google Scholar] [CrossRef]
- Reck, J., Jr.; Berger, M.; Terra RM, S.; Marks, F.S.; da Silva Vaz, I., Jr.; Guimarães, J.A.; Termignoni, C. Systemic alterations of bovine hemostasis due to Rhipicephalus (Boophilus) microplus infestation. Res. Vet. Sci. 2009, 86, 56–62. [Google Scholar] [CrossRef]
- Ceci, L.; Carelli, G.; Boscia, D.; Lacinio, R. The effect of corticosteroid administration on the pathogenicity of Babesia equi infection in horses [Campania]. Atti della Societa Italiana delle Scienze Veterinarie 1997, 51, 543–544. [Google Scholar]
- Oladosu, L.A. Effects of Intravenous Corticosteroid on the Pathogenicity of Babesia equi Infection of Donkeys (Equus asinus). J. Vet. Med. 1988, 35, 509–514. [Google Scholar] [CrossRef]
- Antunes, S.; Galindo, R.C.; Almazán, C.; Rudenko, N.; Golovchenko, M.; Grubhoffer, L.; Shkape, V.; Rosário, V.; de la Fuente, J.; Domingos, A. Functional genomics studies of Rhipicephalus (Boophilus) annulatus ticks in response to infection with the cattle protozoan parasite, Babesia bigemina. Int. J. Parasitol. 2012, 42, 187–195. [Google Scholar] [CrossRef]
- Edwards, K.T.; Goddard, J.; Varela-Stokes, A.S. Examination of the internal morphology of the Ixodid tick, Amblyomma maculatum Koch,(Acari: Ixodidae); a “how-to” pictorial dissection guide. Midsouth Entomol. 2009, 2, 28–39. [Google Scholar]
- Kim, C.; Blanco, L.B.C.; Alhassan, A.; Iseki, H.; Yokoyama, N.; Xuan, X.; Igarashi, I. Diagnostic Real-Time PCR assay for the quantitative detection of Theileria equi from equine blood samples. Vet. Parasitol. 2008, 151, 158–163. [Google Scholar] [CrossRef]
- Nijhof, A.M.; Balk, J.A.; Postigo, M.; Jongejan, F. Selection of reference genes for quantitative RT-PCR studies in Rhipicephalus (Boophilus) microplus and Rhipicephalus appendiculatus ticks and determination of the expression profile of Bm86. BMC Mol. Biol. 2009, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Ghosh, S. Negative regulation of toll-like receptor-mediated signaling by Tollip. J. Biol. Chem. 2002, 277, 7059–7065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopp, E.; Medzhitov, R.; Carothers, J.; Xiao, C.; Douglas, I.; Janeway, C.A.; Ghosh, S. ECSIT is an evolutionarily conserved intermediate in the Toll/IL-1 signal transduction pathway. Genes Dev. 1999, 13, 2059–2071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, S.; Roy, S.; Mistry, H.U.; Murthy, S.; George, N.; Bhandari, V.; Sharma, P. Potential sabotage of host cell physiology by apicomplexan parasites for their survival benefits. Front. Immunol. 2017, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Lee, F.S.; Peters, R.T.; Dang, L.C.; Maniatis, T. MEKK1 activates both IκB kinase α and IκB kinase β. Proc. Natl. Acad. Sci. USA 1998, 95, 9319–9324. [Google Scholar] [CrossRef] [Green Version]
- Silverman, N.; Zhou, R.; Stöven, S.; Pandey, N.; Hultmark, D.; Maniatis, T. A Drosophila IκB kinase complex required for Relish cleavage and antibacterial immunity. Genes Dev. 2000, 14, 2461–2471. [Google Scholar] [CrossRef] [Green Version]
- Tsapras, P.; Petridi, S.; Chan, S.; Geborys, M.; Jacomin, A.C.; Sagona, A.P.; Meier, P.; Nezis, I.P. Selective autophagy controls innate immune response through a TAK1/TAB2/SH3PX1 axis. Cell Rep. 2022, 110286. [Google Scholar] [CrossRef] [PubMed]
- Silverman, N.; Zhou, R.; Erlich, R.L.; Hunter, M.; Bernstein, E.; Schneider, D.; Maniatis, T. Immune activation of NF-κB and JNK requires Drosophila TAK1. J. Biol. Chem. 2003, 278, 48928–48934. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.; Langmann, C.; Harden, N.; Aigaki, T. The RING-finger scaffold protein Plenty of SH3s targets TAK1 to control immunity signaling in Drosophila. EMBO Rep. 2006, 6, 1082–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.M.; Brady, H.; Ruocco, M.G.; Sun, H.; Williams, D.; Lee, S.J.; Kato, T., Jr.; Richards, N.; Chan, K.; Mercurio, F.; et al. Targeting of TAK1 by the NF-κB protein Relish regulates the JNK-mediated immune response in Drosophila. Genes Dev. 2004, 18, 584–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, J.H.; Kim, S.H.; Lee, H.Y.; Bai, J.Y.; Nam, Y.D.; Bae, J.W.; Lee, D.G.; Shin, S.C.; Ha, E.M.; Lee, W.J. Innate immune homeostasis by the homeobox gene caudal and commensal-gut mutualism in Drosophila. Science 2008, 319, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Clayton, A.M.; Cirimotich, C.M.; Dong, Y.; Dimopoulos, G. Caudal is a negative regulator of the Anopheles IMD pathway that controls resistance to Plasmodium falciparum infection. Dev. Comp. Immunol. 2013, 39, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Lizundia, R.; Chaussepied, M.; Huerre, M.; Werling, D.; Di Santo, J.P.; Langsley, G. C-Jun NH2-terminal kinase/c-Jun signaling promotes survival and metastasis of B lymphocytes transformed by Theileria. Cancer Res. 2006, 66, 6105–6110. [Google Scholar] [CrossRef] [Green Version]
- Kershaw, N.J.; Murphy, J.M.; Liau, N.P.; Varghese, L.N.; Laktyushin, A.; Whitlock, E.L.; Lucet, I.S.; Nicola, N.A.; Babon, J.J. SOCS3 binds specific receptor–JAK complexes to control cytokine signaling by direct kinase inhibition. Nat. Struct. Mol. Biol. 2013, 20, 469–476. [Google Scholar] [CrossRef]
- Ueti, W.M.; Palmer, G.H.; Scoles, G.A.; Kappmeyer, L.S.; Knowles, P.D. Persistently infected horses are reservoirs for intrastadial tick-borne transmission of the apicomplexan parasite Babesia equi. Infect. Immun. 2008, 76, 3525–3529. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paulino, P.G.; Peckle, M.; Mendonça, L.P.; Massard, C.L.; Antunes, S.; Couto, J.; Domingos, A.; Guedes Junior, D.d.S.; Cabezas-Cruz, A.; Santos, H.A. Differential Expression of Immune Genes in the Rhipicephalus microplus Gut in Response to Theileria equi Infection. Pathogens 2022, 11, 1478. https://doi.org/10.3390/pathogens11121478
Paulino PG, Peckle M, Mendonça LP, Massard CL, Antunes S, Couto J, Domingos A, Guedes Junior DdS, Cabezas-Cruz A, Santos HA. Differential Expression of Immune Genes in the Rhipicephalus microplus Gut in Response to Theileria equi Infection. Pathogens. 2022; 11(12):1478. https://doi.org/10.3390/pathogens11121478
Chicago/Turabian StylePaulino, Patrícia Gonzaga, Maristela Peckle, Leo Paulis Mendonça, Carlos Luiz Massard, Sandra Antunes, Joana Couto, Ana Domingos, Daniel da Silva Guedes Junior, Alejandro Cabezas-Cruz, and Huarrisson Azevedo Santos. 2022. "Differential Expression of Immune Genes in the Rhipicephalus microplus Gut in Response to Theileria equi Infection" Pathogens 11, no. 12: 1478. https://doi.org/10.3390/pathogens11121478