
Antimicrobial Resistance of Salmonella Strains Isolated from Human, Wild Boar, and Environmental Samples in 2018–2020 in the Northwest of Italy

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
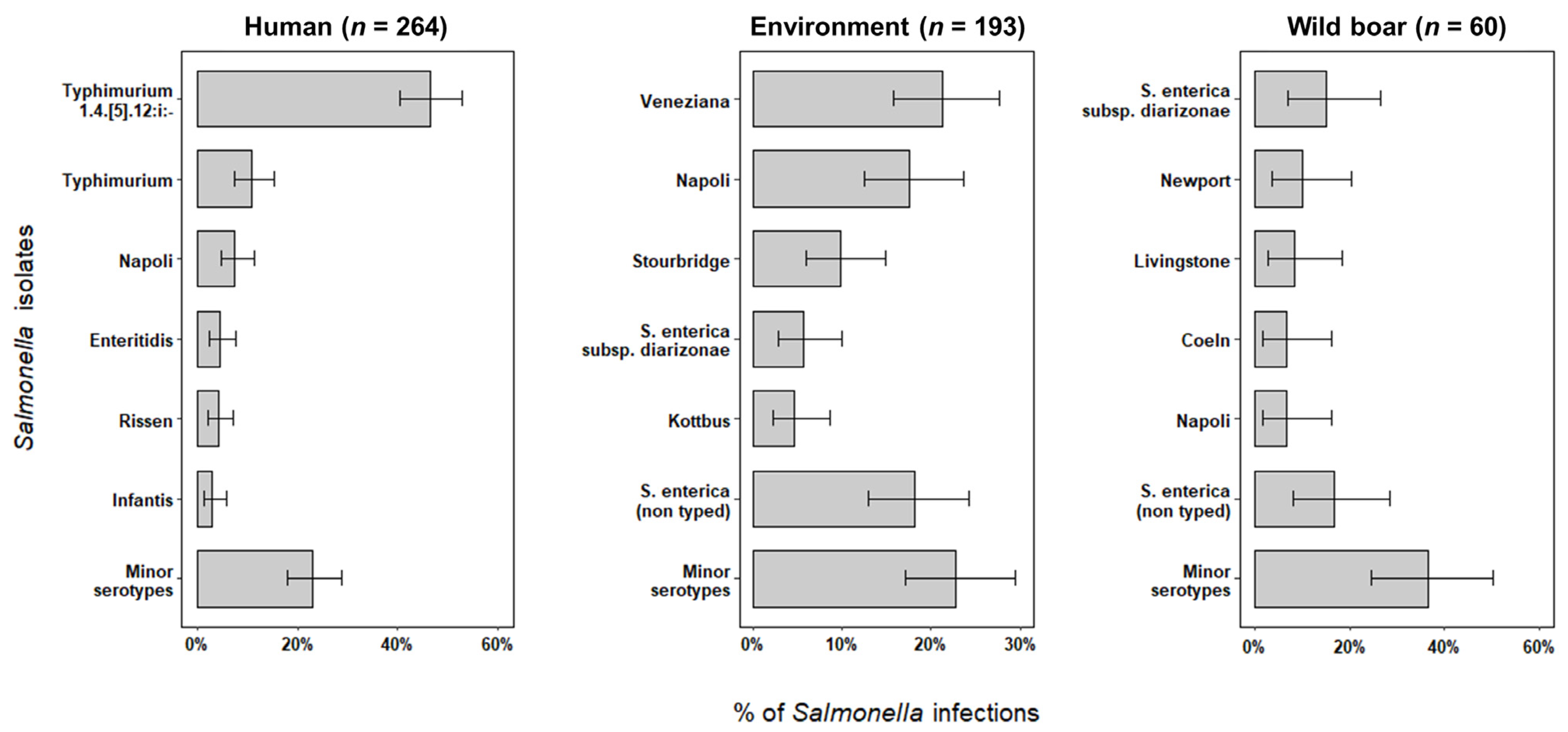
2.1. Salmonella Serotypes and Subtypes Identification
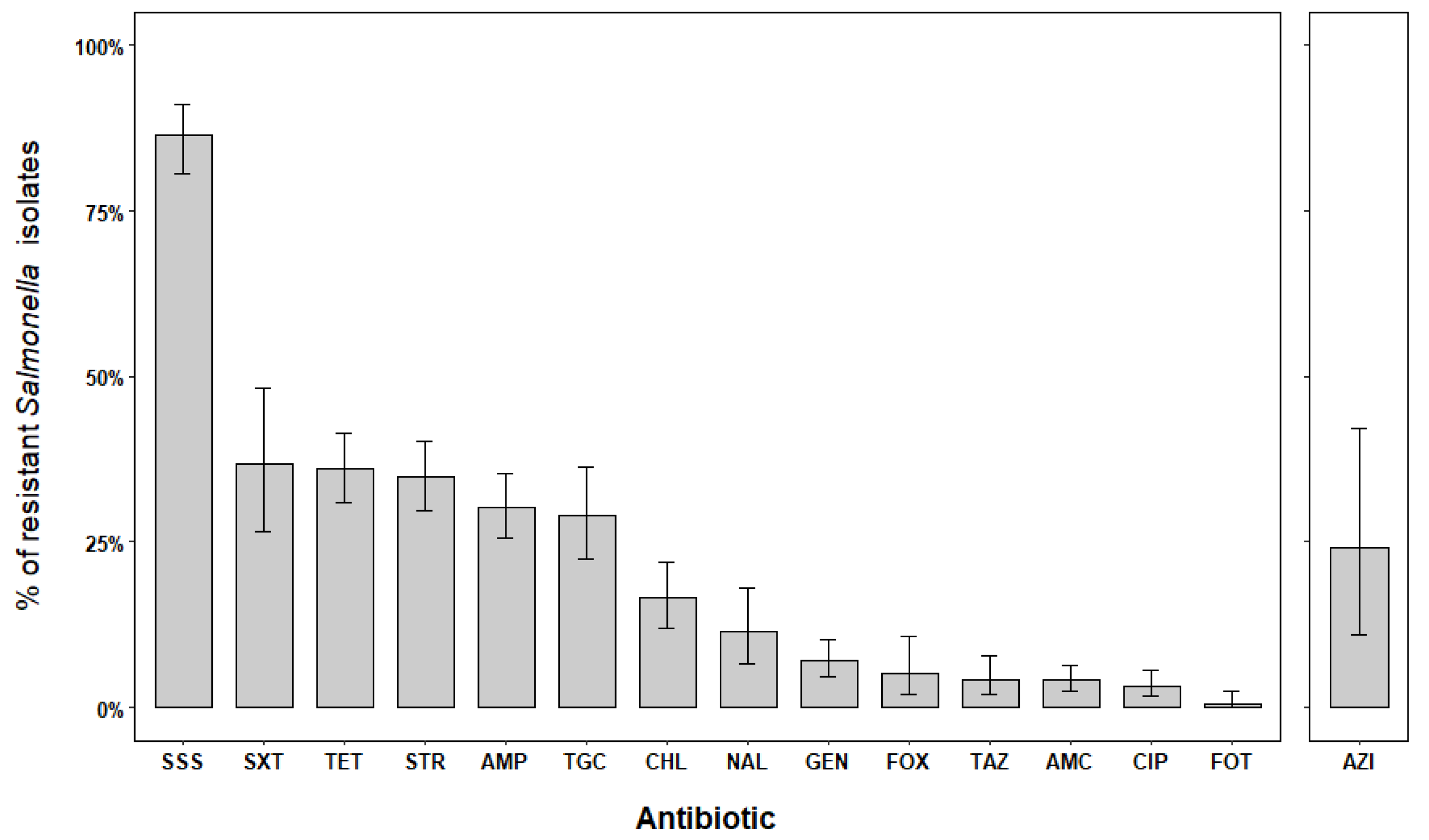
2.2. Antimicrobial Resistances in Salmonella Isolates
2.3. Multi Drug Resistance in Salmonella Isolates
3. Discussion
4. Materials and Methods
4.1. Strains
4.2. Antimicrobial Resistance Analyses
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, 6971. [Google Scholar] [CrossRef]
- Giannella, R.A. Chapter 21: Salmonella. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. Available online: https://www.ncbi.nlm.nih.gov/books/NBK8435/ (accessed on 25 November 2022).
- Gal-Mor, O.; Boyle, E.C.; Grassl, G.A. Same species, different diseases: How and why typhoidal and non-typhoidal Salmonella enterica serovars differ. Front. Microbiol. 2014, 5, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knodler, L.A.; Elfenbein, J.R. Salmonella enterica. Trends Microbiol. 2019, 27, 964–965. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. Salmonella (Non-Typhoidal). Available online: https://www.who.int/news-room/fact-sheets/detail/salmonella-(non-typhoidal) (accessed on 15 May 2022).
- Florez-Cuadrado, D.; Moreno, M.A.; Ugarte-Ruíz, M.; Domínguez, L. Chapter 5: Antimicrobial resistance in the food chain in the European Union. In Advances in Food and Nutrition Research; Rodríguez-Lázaro, D., Ed.; Academic Press: Cambridge, MA, USA, 2018; Volume 86, pp. 115–136. [Google Scholar]
- Nair, V.T.D.; Venkitanarayanan, K.; Kollanoor Johny, A. Antibiotic-resistant Salmonella in the food supply and the potential role of antibiotic alternatives for control. Foods 2018, 7, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Kaye, K.S.; Engemann, J.J.; Fraimow, H.S.; Abrutyn, E. Pathogens resistant to antimicrobial agents: Epidemiology, molecular mechanisms, and clinical management. Infect. Dis. Clin. N. Am. 2004, 18, 467–511. [Google Scholar] [CrossRef]
- Wegener, H.C. Antibiotic resistance—Linking human and animal health. In Institute of Medicine (US). Improving Food Safety Through a One Health Approach: Workshop Summary; National Academies Press (US): Washington, DC, USA, 2012; p. A15. Available online: https://www.ncbi.nlm.nih.gov/books/NBK114485/ (accessed on 25 November 2022).
- Plaza-Rodríguez, C.; Alt, K.; Grobbel, M.; Hammerl, J.A.; Irrgang, A.; Szabo, I.; Stingl, K.; Schuh, E.; Wiehle, L.; Pfefferkorn, B.; et al. Wildlife as sentinels of antimicrobial resistance in Germany? Front. Vet. Sci. 2021, 7, 627821. [Google Scholar] [CrossRef]
- Fredriksson-Ahomaa, M. Wild boar: A reservoir of foodborne zoonoses. Foodborne Pathog. Dis. 2019, 16, 153–165. [Google Scholar] [CrossRef]
- ECDC; EFSA; EMA; OECD. Joint Report by ECDC, EFSA, EMA, and OECD on Antimicrobial Resistance in the EU/EEA and a One Health Response 2022. Available online: https://www.ecdc.europa.eu/en/publications-data/antimicrobial-resistance-eueea-one-health-response (accessed on 15 May 2022).
- Consolidated Text: Directive 2003/99/EC of the European Parliament and of the Council of 17 November 2003 on the Monitoring of Zoonoses and Zoonotic Agents, Amending Council Decision 90/424/EEC and Repealing Council Directive 92/117/EEC. Eurlex Website. Available online: https://eur-lex.europa.eu/legal-content/en/ALL/?uri=CELEX%3A02003L0099-20130701 (accessed on 2 June 2022).
- Mancin, M.; Barco, L.; Losasso, C.; Belluco, S.; Cibin, V.; Mazzucato, M.; Bilei, S.; Carullo, M.R.; Decastelli, L.; Di Giannatale, E.; et al. Salmonella serovar distribution from non-human sources in Italy; results from the IT-Enter-Vet network. Vet. Rec. 2018, 183, 69. [Google Scholar] [CrossRef]
- Sacchini, L.; Ricci, L.; Zilli, K.; Romantini, R.; Di Donato, G.; Neri, D.; Persiani, T.; Di Giannatale, E. Characterization of Salmonella Typhimurium and monophasic Salmonella Typhimurium isolated in Abruzzo and Molise regions, Italy, from 2012 to 2017. Vet. Ital. 2021, 57, 297–304. [Google Scholar] [CrossRef]
- Alessiani, A.; Goffredo, E.; Mancini, M.; Occhiochiuso, G.; Faleo, S.; Didonna, A.; Fischetto, R.; Suglia, F.; De Vito, D.; Stallone, A.; et al. Evaluation of antimicrobial resistance in Salmonella strains isolated from food, animal and human samples between 2017 and 2021 in southern Italy. Microorganisms 2022, 10, 812. [Google Scholar] [CrossRef]
- Graziani, C.; Busani, L.; Dionisi, A.M.; Caprioli, A.; Ivarsson, S.; Hedenström, I.; Luzzi, I. Virulotyping of Salmonella enterica serovar Napoli strains isolated in Italy from human and nonhuman sources. Foodborne Pathog. Dis. 2011, 8, 997–1003. [Google Scholar] [CrossRef]
- Graziani, C.; Luzzi, I.; Owczarek, S.; Dionisi, A.M.; Busani, L. Salmonella enterica serovar Napoli infection in Italy from 2000 to 2013: Spatial and spatio-temporal analysis of cases distribution and the effect of human and animal density on the risk of infection. PLoS ONE 2015, 10, e0142419. [Google Scholar] [CrossRef] [Green Version]
- Sabbatucci, M.; Dionisi, A.M.; Pezzotti, P.; Lucarelli, C.; Barco, L.; Mancin, M.; Luzzi, I. Molecular and epidemiologic analysis of re-emergent Salmonella enterica serovar Napoli, Italy, 2011–2015. Emerg. Infect. Dis. 2018, 24, 562–565. [Google Scholar] [CrossRef] [Green Version]
- Razzuoli, E.; Listorti, V.; Martini, I.; Migone, L.; Decastelli, L.; Mignone, W.; Berio, E.; Battistini, R.; Ercolini, C.; Serracca, L.; et al. Prevalence and antimicrobial resistances of Salmonella spp. isolated from wild boars in Liguria region, Italy. Pathogens 2021, 10, 568. [Google Scholar] [CrossRef]
- Proroga, Y.T.R.; Capuano, F.; Carullo, M.R.; La Tela, I.; Capparelli, R.; Barco, L.; Pasquale, V. Occurrence and antimicrobial resistance of Salmonella strains from food of animal origin in southern Italy. Folia Microbiol. 2016, 61, 21–27. [Google Scholar] [CrossRef]
- Mellou, K.; Gkova, M.; Panagiotidou, E.; Tzani, M.; Sideroglou, T.; Mandilara, G. Diversity and resistance profiles of human non-typhoidal Salmonella spp. in Greece, 2003–2020. Antibiotics 2021, 10, 983. [Google Scholar] [CrossRef]
- McMillan, E.A.; Jackson, C.R.; Frye, J.G. Transferable plasmids of Salmonella enterica associated with antibiotic resistance genes. Front. Microbiol. 2020, 11, 562181. [Google Scholar] [CrossRef]
- Di Marcantonio, L.; Romantini, R.; Marotta, F.; Chiaverini, A.; Zilli, K.; Abass, A.; Di Giannatale, E.; Garofolo, G.; Janowicz, A. The current landscape of antibiotic resistance of Salmonella Infantis in Italy: The expansion of extended-spectrum beta-lactamase producers on a local scale. Front. Microbiol. 2022, 13, 812481. [Google Scholar] [CrossRef]
- Chironna, M.; Tafuri, S.; Gallone, M.S.; Sallustio, A.; Martinelli, D.; Prato, R.; Germinario, C. Outbreak of Salmonella infantis gastroenteritis among people who had eaten at a hash house in southern Italy. Public Health 2014, 128, 438–443. [Google Scholar] [CrossRef]
- Franco, A.; Leekitcharoenphon, P.; Feltrin, F.; Alba, P.; Cordaro, G.; Iurescia, M.; Tolli, R.; D’Incau, M.; Staffolani, M.; Di Giannatale, E.; et al. Emergence of a clonal lineage of multidrug-resistant ESBL-producing Salmonella Infantis transmitted from broilers and broiler meat to humans in Italy between 2011 and 2014. PLoS ONE 2016, 10, e0144802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sévellec, Y.; Granier, S.A.; Le Hello, S.; Weill, F.X.; Guillier, L.; Mistou, M.Y.; Cadel-Six, S. Source attribution study of sporadic Salmonella Derby cases in France. Front. Microbiol. 2020, 11, 889. [Google Scholar] [CrossRef] [PubMed]
- García-Fernández, A.; Gallina, S.; Owczarek, S.; Dionisi, A.M.; Benedetti, I.; Decastelli, L.; Luzzi, I. Emergence of ciprofloxacin-resistant Salmonella enterica serovar Typhi in Italy. PLoS ONE 2015, 10, e0132065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haley, B.J.; Cole, D.J.; Lipp, E.K. Distribution, diversity, and seasonality of waterborne salmonellae in a rural watershed. Appl. Environ. Microbiol. 2009, 75, 1248–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saporito, L.; Colomba, C.; Scarlata, F.; Li Vecchi, V.; Mammina, C.; Titone, L. Clinical and microbiological features of Salmonella gastroenteritis in children. Inf. Med. 2007, 1, 24–29. [Google Scholar]
- Botti, V.; Navillod, F.V.; Domenis, L.; Orusa, R.; Pepe, E.; Robetto, S.; Guidetti, C. Salmonella spp. and antibiotic-resistant strains in wild mammals and birds in north-western Italy from 2002 to 2010. Vet. Ital. 2013, 49, 187–294. [Google Scholar]
- Chiari, M.; Zanoni, M.; Tagliabue, S.; Lavazza, A.; Alborali, L.G. Salmonella serotypes in wild boars (Sus scrofa) hunted in northern Italy. Acta Vet. Scand. 2013, 55, 42. [Google Scholar] [CrossRef] [Green Version]
- Bonardi, S.; Bolzoni, L.; Zanoni, R.G.; Morganti, M.; Corradi, M.; Gilioli, S.; Pongolini, S. Limited Exchange of Salmonella Among Domestic Pigs and Wild Boars in Italy. Ecohealth 2019, 16, 420–428. [Google Scholar] [CrossRef]
- Baudart, J.; Lemarchand, K.; Brisabois, A.; Lebaron, P. Diversity of Salmonella strains isolated from the aquatic environment as determined by serotyping and amplification of the ribosomal DNA spacer regions. Appl. Environ. Microbiol. 2000, 66, 1544–1552. [Google Scholar] [CrossRef] [Green Version]
- European Centre for Disease Prevention and Control, ECDC. Increase in Salmonella Stourbridge Infections in Germany during 2016; ECDC: Stockholm, Sweden, 2016; Available online: https://www.ecdc.europa.eu/en/publications-data/increase-salmonella-stourbridge-infections-germany-during-2016-16-december-2016 (accessed on 25 November 2022).
- Bennett, S.; Littrell, K.; Hill, T.; Mahovic, M.; Behravesh, C. Multistate foodborne disease outbreaks associated with raw tomatoes, United States, 1990-2010: A recurring public health problem. Epidemiol. Infect. 2015, 143, 1352–1359. [Google Scholar] [CrossRef]
- Robinson, E.; Travanut, M.; Fabre, L.; Larréché, S.; Ramelli, L.; Pascal, L.; Guinard, A.; Vincent, N.; Calba, C.; Meurice, L.; et al. Outbreak of Salmonella Newport associated with internationally distributed raw goats’ milk cheese, France, 2018. Epidemiol. Infect. 2020, 148, e180. [Google Scholar] [CrossRef]
- Mughini-Gras, L.; Pijnacker, R.; Duijster, J.; Heck, M.; Wit, B.; Veldman, K.; Franz, E. Changing epidemiology of invasive non-typhoid Salmonella infection: A nationwide population-based registry study. Clin. Microbiol. Infect. 2020, 26, 941.e9–941.e14. [Google Scholar] [CrossRef]
- Antimicrobial Resistance in Europe. Resistance of Salmonella, E. coli and Campylobacter in Food, Animals and Humans, Country by Country in 2019 and 2020. Available online: https://multimedia.efsa.europa.eu/dataviz-2020/index.htm (accessed on 23 November 2022).
- Crump, J.A.; Sjölund-Karlsson, M.; Gordon, M.A.; Parry, C.M. Epidemiology, clinical presentation, laboratory diagnosis, antimicrobial resistance, and antimicrobial management of invasive Salmonella infections. Clin. Microbiol. Rev. 2015, 28, 901–937. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Hu, D.; Zhang, Q.; Liao, X.P.; Liu, Y.H.; Sun, J. Efflux pump overexpression contributes to tigecycline heteroresistance in Salmonella enterica serovar Typhimurium. Front. Cell Infect. Microbiol. 2017, 7, 37. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Aziz, N.K.; Tartor, Y.H.; Gharieb, R.M.A.; Erfan, A.M.; Khalifa, E.; Said, M.A.; Ammar, A.M.; Samir, M. Extensive drug-resistant Salmonella enterica isolated from poultry and humans: Prevalence and molecular determinants behind the co-resistance to ciprofloxacin and tigecycline. Front. Microbiol. 2021, 12, 738784. [Google Scholar] [CrossRef]
- Uddin, M.B.; Hossain, S.; Hasan, M.; Alam, M.N.; Debnath, M.; Begum, R.; Roy, S.; Harun-Al-Rashid, A.; Chowdhury, M.; Rahman, M.M.; et al. Multidrug antimicrobial resistance and molecular detection of mcr-1 gene in Salmonella species isolated from chicken. Animals 2021, 11, 206. [Google Scholar] [CrossRef]
- Su, L.H.; Chiu, C.H.; Chu, C.; Ou, J.T. Antimicrobial resistance in nontyphoid Salmonella serotypes: A global challenge. Clin. Infect. Dis. 2004, 39, 546–551. [Google Scholar] [CrossRef] [Green Version]
- Britto, C.D.; Wong, V.K.; Dougan, G.; Pollard, A.J. A systematic review of antimicrobial resistance in Salmonella enterica serovar Typhi, the etiological agent of typhoid. PLoS Negl. Trop. Dis. 2018, 12, e0006779. [Google Scholar] [CrossRef] [Green Version]
- Song, Q.; Xu, Z.; Gao, H.; Zhang, D. Overview of the development of quinolone resistance in Salmonella species in China, 2005–2016. Infect. Drug Resist. 2018, 11, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Saito, S.; Koori, Y.; Ohsaki, Y.; Osaka, S.; Oana, K.; Nagano, Y.; Arakawa, Y.; Nagano, N. Third-generation cephalosporin-resistant non-typhoidal Salmonella isolated from human feces in Japan. Jpn. J. Infect. Dis. 2017, 70, 301–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, L.; Hopkins, K.L.; Meunier, D.; De Pinna, E.; Fitzgerald-Hughes, D.; Humphreys, H.; Woodford, N. Resistance to third-generation cephalosporins in human non-typhoidal Salmonella enterica isolates from England and Wales, 2010–2012. J. Antimicrob. Chemother. 2014, 69, 977–981. [Google Scholar] [CrossRef] [PubMed]
- Marchello, C.S.; Carr, S.D.; Crump, J.A. A Systematic review on antimicrobial resistance among Salmonella Typhi worldwide. Am. J. Trop. Med. Hyg. 2020, 103, 2518–2527. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, M.; Simon, S.; Meinen, A.; Trost, E.; Banerji, S.; Pfeifer, Y.; Flieger, A. Third generation cephalosporin resistance in clinical non-typhoidal Salmonella enterica in Germany and emergence of blaCTX-M-harbouring pESI plasmids. Microb. Genom. 2021, 7, 000698. [Google Scholar] [CrossRef] [PubMed]
- Falgenhauer, L.; Ghosh, H.; Guerra, B.; Yao, Y.; Fritzenwanker, M.; Fischer, J.; Helmuth, R.; Imirzalioglu, C.; Chakraborty, T. Comparative genome analysis of IncHI2 VIM-1 carbapenemase-encoding plasmids of Escherichia coli and Salmonella enterica isolated from a livestock farm in Germany. Vet. Microbiol. 2017, 200, 114–117. [Google Scholar] [CrossRef]
- Gao, Y.; Wen, J.; Wang, S.; Xu, X.; Zhan, Z.; Chen, Z.; Bai, J.; Qu, X.; Zhang, H.; Zhang, J. Plasmid-encoded blaNDM-5 gene that confers high-level carbapenem resistance in Salmonella Typhimurium of pork origin. Infect. Drug Resist. 2020, 13, 1485. [Google Scholar] [CrossRef]
- Wang, Z.; He, J.; Li, Q.; Tang, Y.; Wang, J.; Pan, Z.; Chen, X.; Jiao, X. First detection of NDM-5-positive Salmonella enterica serovar Typhimurium isolated from retail pork in China. Microb. Drug Resist. 2020, 26, 434–437. [Google Scholar] [CrossRef]
- Bogdanić, N.; Močibob, L.; Vidović, T.; Soldo, A.; Begovać, J. Azithromycin consumption during the COVID-19 pandemic in Croatia, 2020. PLoS ONE 2022, 17, e0263437. [Google Scholar] [CrossRef]
- Echeverría-Esnal, D.; Martin-Ontiyuelo, C.; Navarrete-Rouco, M.E.; De-Antonio Cuscó, M.; Ferrández, O.; Horcajada, J.P.; Grau, S. Azithromycin in the treatment of COVID-19: A review. Expert Rev. Anti-Infect. Ther. 2021, 19, 147–163. [Google Scholar] [CrossRef]
- The Medicines Utilisation Monitoring Centre. National Report on Antibiotics Use in Italy; Italian Medicines Agency: Rome, Italy, 2022.
- Proroga, Y.; Capuano, F.; Capparelli, R.; Bilei, S.; Bernardo, M.; Cocco, M.P.; Campagnuolo, R.; Pasquale, V. Characterization of non-typhoidal Salmonella enterica strains of human origin in central and southern Italy. Ital. J. Food Saf. 2018, 7, 6888. [Google Scholar] [CrossRef]
- La Tela, I.; Peruzy, M.F.; D’Alessio, N.; Di Nocera, F.; Casalinuovo, F.; Carullo, M.R.; Cardinale, D.; Cristiano, D.; Capuano, F. Serotyping and evaluation of antimicrobial resistance of Salmonella strains detected in wildlife and natural environments in southern Italy. Antibiotics 2021, 10, 353. [Google Scholar] [CrossRef]
- Lee, H.Y.; Su, L.H.; Tsai, M.H.; Kim, S.W.; Chang, H.H.; Jung, S.I.; Park, K.H.; Perera, J.; Carlos, C.; Tan, B.H.; et al. High rate of reduced susceptibility to ciprofloxacin and ceftriaxone among nontyphoid Salmonella clinical isolates in Asia. Antimicrob. Agents Chemother. 2009, 53, 2696–2699. [Google Scholar] [CrossRef]
- Sriyapai, P.; Pulsrikarn, C.; Chansiri, K.; Nyamniyom, A.; Sriyapai, T. Molecular characterization of cephalosporin and fluoroquinolone resistant Salmonella Choleraesuis isolated from patients with systemic salmonellosis in Thailand. Antibiotics 2021, 10, 844. [Google Scholar] [CrossRef]
- Balbin, M.M.; Hull, D.; Guest, C.; Nichols, L.; Dunn, R.; Hull, D.; Thakur, S. Antimicrobial resistance and virulence factors profile of Salmonella spp. and Escherichia coli isolated from different environments exposed to anthropogenic activity. J. Glob. Antimicrob. Resist. 2020, 22, 578–583. [Google Scholar] [CrossRef]
- Larsson, D.G.J.; Flach, C.F. Antibiotic resistance in the environment. Nat. Rev. Microbiol. 2022, 20, 257–269. [Google Scholar] [CrossRef]
- Italian Ministry of Health. National Plan to Combat Antimicrobial Resistance (PNCAR), 2017–2020. Available online: https://www.salute.gov.it/imgs/C_17_pubblicazioni_2660_allegato.pdf (accessed on 15 May 2022).
- Listorti, V.; Battistini, R.; Ercolini, C.; Tramuta, C.; Razzuoli, E.; Vencia, W.; Decastelli, L.; Gallina, S.; Masotti, C.; Serracca, L. In vitro susceptibility of multidrug resistant strains of Salmonella to Essential Oils. Nat. Prod. Commun. 2020, 15, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Zottola, T.; Montagnaro, S.; Magnapera, C.; Sasso, S.; De Martino, L.; Bragagnolo, A.; D’Amici, L.; Condoleo, R.; Pisanelli, G.; Iovane, G.; et al. Prevalence and antimicrobial susceptibility of Salmonella in European wild boar (Sus scrofa); Latium Region—Italy. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 161–168. [Google Scholar] [CrossRef]
- Cilia, G.; Turchi, B.; Fratini, F.; Bilei, S.; Bossù, T.; De Marchis, M.L.; Cerri, D.; Pacini, M.I.; Bertelloni, F. Prevalence, virulence and antimicrobial susceptibility of Salmonella spp.; Yersinia enterocolitica and Listeria monocytogenes in European Wild boar (Sus scrofa) hunted in Tuscany (central Italy). Pathogens 2021, 10, 93. [Google Scholar] [CrossRef]
- Torres, R.T.; Joana Fernandes, J.; Carvalho, J.; Cunha, M.V.; Caetano, T.; Mendo, S.; Serrano, E.; Fonseca, C. Wild boar as a reservoir of antimicrobial resistance. Sci. Total Environ. 2020, 717, 135001. [Google Scholar] [CrossRef]
- Clinical Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- U.S. Food and Drug Administration. Antibacterial Susceptibility Test Interpretive Criteria. Available online: https://www.fda.gov/drugs/development-resources/antibacterial-susceptibility-test-interpretive-criteria (accessed on 2 June 2022).
- StataCorp. Stata Statistical Software: Release 17; StataCorp LLC: College Station, TX, USA, 2021. [Google Scholar]



{kind=link}
{kind=link}
| Serotype/Subtype Identified | Human (n = 264) | Environment (n = 193) | Wild Boar (n = 60) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | % | (95% CI) | n | % | (95% CI) | n | % | (95% CI) | |
| S. enterica subsp. enterica (non-typed) | 5 | 1.9 | (0.6–4.4) | 35 | 18.1 | (13.0–24.3) | 10 | 16.7 | (8.3–28.5) |
| 6,7:c:1,5 | 3 | 1.1 | (0.2–3.3) | 2 | 1.0 | (0.1–3.7) | − | − | |
| S. Bovismorbificans | − | − | − | − | 3 | 5.0 | (1.0–13.9) | ||
| S. Brandenburg | 5 | 1.9 | (0.6–4.4) | 1 | 0.5 | (0.01–3.0) | − | − | |
| S. Bredeney | 3 | 1.1 | (0.2–3.3) | 1 | 0.5 | (0.01–3.0) | − | − | |
| S. Carno | 1 | 0.4 | (0.01–2.1) | − | − | − | − | ||
| S. Cerro | − | − | 1 | 0.5 | (0.01–3.0) | − | − | ||
| S. Choleraesuis var. Kunzendorf | − | − | − | − | 3 | 5.0 | (1.0–13.9) | ||
| S. Coeln | 5 | 1.9 | (0.6–4.4) | 8 | 4.1 | (1.8–8.0) | − | − | |
| S. Derby | 3 | 1.1 | (0.2–3.3) | − | − | − | − | ||
| S. Drogana | 1 | 0.4 | (0.01–2.1) | − | − | − | − | ||
| S. Eingedi | − | − | 1 | 0.5 | (0.01–3.0) | − | − | ||
| S. Enteritidis | 12 | 4.5 | (2.4–7.8) | − | − | 2 | 3.3 | (0.4–11.5) | |
| S. Fann | − | − | 1 | 0.5 | (0.01–3.0) | − | − | ||
| S. Give | 3 | 1.1 | (0.2–3.3) | − | − | − | − | ||
| S. Goldcoast | 3 | 1.1 | (0.2–3.3) | 1 | 0.5 | (0.01–3.0) | |||
| S. Hadar | 1 | 0.4 | (0.01–2.1) | − | − | − | − | ||
| S. Ilugun | − | − | 2 | 1.0 | (0.1–3.7) | − | − | ||
| S. Infantis | 8 | 3.0 | (1.3–5.9) | − | − | − | |||
| S. Kapemba | 2 | 0.8 | (0.1–2.7) | 1 | 0.5 | (0.01–3.0) | − | − | |
| S. Kentucky | 1 | 0.4 | (0.01–2.1) | − | − | − | − | ||
| S. Kortrijk | 1 | 0.4 | (0.01–2.1) | − | − | − | − | ||
| S. Kottbus | − | − | 9 | 4.7 | (2.2–8.7) | 3 | 5.0 | (1.0–13.9) | |
| S. Litchfield | 1 | 0.4 | (0.01–2.1) | − | − | − | − | ||
| S. Livingstone | 1 | 0.4 | (0.01–2.1) | − | − | 5 | 8.3 | (2.8–18.4) | |
| S. London | 3 | 1.1 | (0.2–3.3) | 1 | 0.5 | (0.01–3.0) | − | − | |
| S. Mbandaka | 1 | 0.4 | (0.01–2.1) | − | − | − | − | ||
| S. Muenster | 1 | 0.4 | (0.01–2.1) | − | − | 3 | 5.0 | (1.0–13.9) | |
| S. Napoli | 20 | 7.6 | (4.7–11.5) | 34 | 17.6 | (12.5–23.7) | 4 | 6.7 | (1.8–16.2) |
| S. Newport | 1 | 0.4 | (0.01–2.1) | 1 | 0.5 | (0.01–3.0) | 6 | 10.0 | (3.8–20.5) |
| S. Othmarschen | 2 | 0.8 | (0.1–2.7) | − | − | − | − | ||
| S. Panama | 1 | 0.4 | (0.01–2.1) | − | − | − | − | ||
| S. Patience | 1 | 0.4 | (0.01–2.1) | − | − | − | − | ||
| S. Presov | − | − | 1 | 0.5 | (0.01–3.0) | − | − | ||
| S. Rissen | 11 | 4.2 | (2.1–7.3) | 2 | 1.0 | (0.1–3.7) | − | − | |
| S. Senftenberg | 1 | 0.4 | (0.01–2.1) | − | − | − | − | ||
| S. Stoneferry | − | − | − | − | 2 | 3.3 | (0.4–11.5) | ||
| S. Stourbridge | 1 | 0.4 | (0.01–2.1) | 19 | 9.8 | (6.0–15.0) | 2 | 3.3 | (0.4–11.5) |
| S. Thompson | 3 | 1.1 | (0.2–3.3) | 8 | 4.1 | (1.8–8.0) | 1 | 1.7 | (0.04–8.9) |
| S. Typhi | 2 | 0.8 | (0.1–2.7) | − | − | − | |||
| S. Typhimurium | 29 | 11.0 | (7.5–15.4) | 1 | 0.5 | (0.01–3.0) | − | − | |
| S. Typhimurium 1,4,[5],12:i:- | 123 | 46.6 | (40.5–52.8) | 5 | 2.6 | (0.8–5.9) | 1 | 1.7 | (0.04–8.9) |
| S. Veneziana | 1 | 0.4 | (0.01–2.1) | 41 | 21.2 | (15.7–27.7) | − | − | |
| S. Virchow | 1 | 0.4 | (0.01–2.1) | − | − | − | − | ||
| S. Wohlen | 1 | 0.4 | (0.01–2.1) | 1 | 0.5 | (0.01–3.0) | − | − | |
| S. enterica subsp. diarizonae | 1 | 0.4 | (0.01–2.1) | 11 | 5.7 | (2.9–10.0) | 9 | 15.0 | (7.1–26.6) |
| S. enterica subsp. houtenae | − | − | 1 | 0.5 | (0.01–3.0) | 2 | 3.3 | (0.4–11.5) | |
| S. enterica subsp. salamae | 2 | 0.8 | (0.1–2.7) | 4 | 2.1 | (0.6–5.2) | − | − | |
| No. of Isolates (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| SOURCE | AMR | AMC | AMP | AZI a | FOT | FOX | TAZ | CIP | CHL |
| HUMAN | S | 149/198 (75.3) | 113/228 (49.6) | − | 221/229 (96.5) | 110/118 (93.2) | 203/215 (94.4) | 197/228 (86.4) | 189/230 (82.2) |
| I | 34/198 (17.2) *** | 4/228 (1.8) | 23/23 (100) *** | 7/229 (3.1) | 2/118 (1.7) | 3/203 (1.4) | 21/228 (9.2) ** | 3/230 (1.3) | |
| R | 15/198 (7.6) * | 111/228 (48.7) *** | − | 1/229 (0.4) | 6/118 (5.1) | 9/215 (4.2) | 10/228 (4.4) | 38/230 (16.5) | |
| ENVIRONMENT | S | 124/128 (96.8) | 140/145 (96.5) | − | 141/145 (97.2) | 64/65 (98.5) | 125/127 (98.4) | 141/145 (97.2) | 145/145 (100) |
| I | 2/128 (1.6) *** | 3/145 (2.1) | 2/10 (20.0) *** | 4/141 (2.8) | 1/65 (1.5) | 2/127 (1.6) | 2/145 (1.4) ** | − | |
| R | 2/128 (1.6) * | 2/145 (1.4) *** | 8/10 (80.0) | − | − | − | 2/145 (1.4) | − | |
| WILD BOAR | S | − | 30/30 (100) | − | 39/39 (100) | − | 39/39 (100) | 39/39 (100) | 3/3 (100) |
| R | − | − | − | − | − | − | − | ||
| No. of Isolates (%) | |||||||||
| SOURCE | AMR | GEN | MER a | NAL | STR | SSS | TET | TGC b | SXT b |
| HUMAN | S | 173/230 (81.6) | 32/33 (97.0) | 100/139 (71.9) | 84/208 (40.4) | 5/117 (4.3) | 81/206 (39.3) | 47/99 (47.5) | 7/21 (33.3) |
| I | 28/212 (13.2) | 1/33 (3.0) | 23/139 (16.5) | 22/208 (10.6) | 3/117 (2.6) | 11/206 (5.3) | 5/99 (5.1) | − | |
| R | 11/212 (5.2) | − | 16/139 (11.5) | 102/208 (49.0) *** | 109/117 (93.2) *** | 114/206 (55.3) *** | 47/99 (47.5) * | 14/21 (66.7) | |
| ENVIRONMENT | S | 102/144 (70.8) | 19/19 (100) | 59/65 (90.8) | 91/130 (70.0) | 13/66 (19.7) | 114/137 (83.2) | 72/80 (90.0) | 10/24 (41.7) |
| I | 28/144 (19.4) | − | 6/65 (9.2) | 23/130 (17.7) | 4/66 (6.1) | 13/137 (9.5) | 3/80 (3.8) | − | |
| R | 14/144 (9.7) | − | − | 16/130 (12.3) *** | 49/66 (74.2) | 10/137 (7.3) *** | 5/80 (6.2) * | 14/24 (58.3) | |
| WILD BOAR | S | 39/39 (100) | 35/35 (100) | 4/4 (100) | − | − | 4/4 (100) | 35/35 (100) | 36/39 (92.3) |
| R | − | − | − | − | − | − | 3/39 (7.7) | ||
| Serotype/Subspecies Involved | Source | No. of Strains | Cephalosporines and/or Quinolones | Antimicrobial Resistance Profile |
|---|---|---|---|---|
| 6,7:c:1,5 | human | 1 | Nalidixic acid | AMP-STR-SSS-NAL-SXT |
| S. Typhimurium 1,4,[5],12:i:- | human | 1 | Nalidixic acid | SSS-NAL |
| S. Typhimurium 1,4,[5],12:i:- | human | 1 | Ceftazidime | AMP-TAZ-STR-TET |
| S. Typhimurium 1,4,[5],12:i:- | human | 1 | Ceftazidime | AMP-TAZ-STR-SSS-TET |
| S. Typhimurium 1,4,[5],12:i:- | human | 1 | Cefoxitin Ceftazidime | AMP-FOX-TAZ-STR-SSS |
| S. Typhimurium 1,4,[5],12:i:- | human | 1 | Cefoxitin Ceftazidime | AMP-FOX-TAZ-STR-SSS-TET |
| S. Typhimurium 1,4,[5],12:i:- | human | 1 | Cefoxitin | AMC-AMP-FOX-CHL-STR-SSS-TET |
| S. Coeln | human | 1 | Ceftazidime | TAZ-SSS |
| S. enterica subsp. enterica | Environm. | 1 | Ciprofloxacin | AZI-CIP-GEN-SXT |
| S. Enteritidis | human | 1 | Nalidixic acid | SSS-NAL |
| S. Enteritidis | human | 1 | Nalidixic acid | AMP-STR-SSS-NAL |
| S. Goldcoast | Environm. | 1 | Ciprofloxacin | AZI-CIP-TET-TGC-SXT |
| S. Hadar | human | 1 | Nalidixic acid | AMP-STR-SSS-TET-NAL |
| S. Infantis | human | 1 | Ciprofloxacin | CIP |
| S. Infantis | human | 3 | Ciprofloxacin Nalidixic acid | CIP-STR-SSS-TET-NAL-SXT |
| S. Infantis | human | 1 | Nalidixic acid | SSS-TET-NAL |
| S. Infantis | human | 1 | Nalidixic acid | AMP-SSS-TET-NAL |
| S. Kentucky | human | 2 | Ciprofloxacin Nalidixic acid | AMP-CIP-GEN-STR-SSS-TET-NAL |
| S. Napoli | human | 1 | Ceftazidime | AMP-TAZ-CHL-STR-SSS-TET |
| S. Othmarschen | human | 1 | Cefoxitin Ceftazidime Ciprofloxacin | AMP-FOX-TAZ-CIP-SSS |
| S. Rissen | human | 2 | Ciprofloxacin Nalidixic acid | AMP-CIP-TET-NAL |
| S. Senftenberg | human | 1 | Cefotaxime | FOT |
| S. Thompson | human | 1 | Ceftazidime | TAZ |
| S. Typhi | human | 1 | Ciprofloxacin Nalidixic acid | CIP-NAL |
| S. Typhi | human | 1 | Ciprofloxacin Nalidixic acid | AMP-CHL-CIP-STR-SSS-NAL-SXT |
| S. Typhimurium | human | 1 | Cefoxitin | FOX-SSS |
| S. Typhimurium | human | 2 | Ceftazidime Ciprofloxacin Nalidixic acid | AMP-TAZ-CIP-CHL-STR-SSS-TET-NAL- |
| S. Typhimurium | human | 1 | Ciprofloxacin | AMC-AMP-CIP-CHL-GEN-STR-TET-TGC |
| S. Typhimurium | human | 1 | Nalidixic acid | AMC-AMP-STR-SSS-TET-NAL |
| S. Virchow | human | 1 | Nalidixic acid | SSS-NAL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Listorti, V.; Garcia-Vozmediano, A.; Pitti, M.; Maurella, C.; Adriano, D.; Ercolini, C.; Dellepiane, M.; Guardone, L.; Razzuoli, E. Antimicrobial Resistance of Salmonella Strains Isolated from Human, Wild Boar, and Environmental Samples in 2018–2020 in the Northwest of Italy. Pathogens 2022, 11, 1446. https://doi.org/10.3390/pathogens11121446
Listorti V, Garcia-Vozmediano A, Pitti M, Maurella C, Adriano D, Ercolini C, Dellepiane M, Guardone L, Razzuoli E. Antimicrobial Resistance of Salmonella Strains Isolated from Human, Wild Boar, and Environmental Samples in 2018–2020 in the Northwest of Italy. Pathogens. 2022; 11(12):1446. https://doi.org/10.3390/pathogens11121446
Chicago/Turabian StyleListorti, Valeria, Aitor Garcia-Vozmediano, Monica Pitti, Cristiana Maurella, Daniela Adriano, Carlo Ercolini, Monica Dellepiane, Lisa Guardone, and Elisabetta Razzuoli. 2022. "Antimicrobial Resistance of Salmonella Strains Isolated from Human, Wild Boar, and Environmental Samples in 2018–2020 in the Northwest of Italy" Pathogens 11, no. 12: 1446. https://doi.org/10.3390/pathogens11121446