

Tsetse Flies Infected with Trypanosomes in Three Active Human African Trypanosomiasis Foci of the Republic of Congo
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
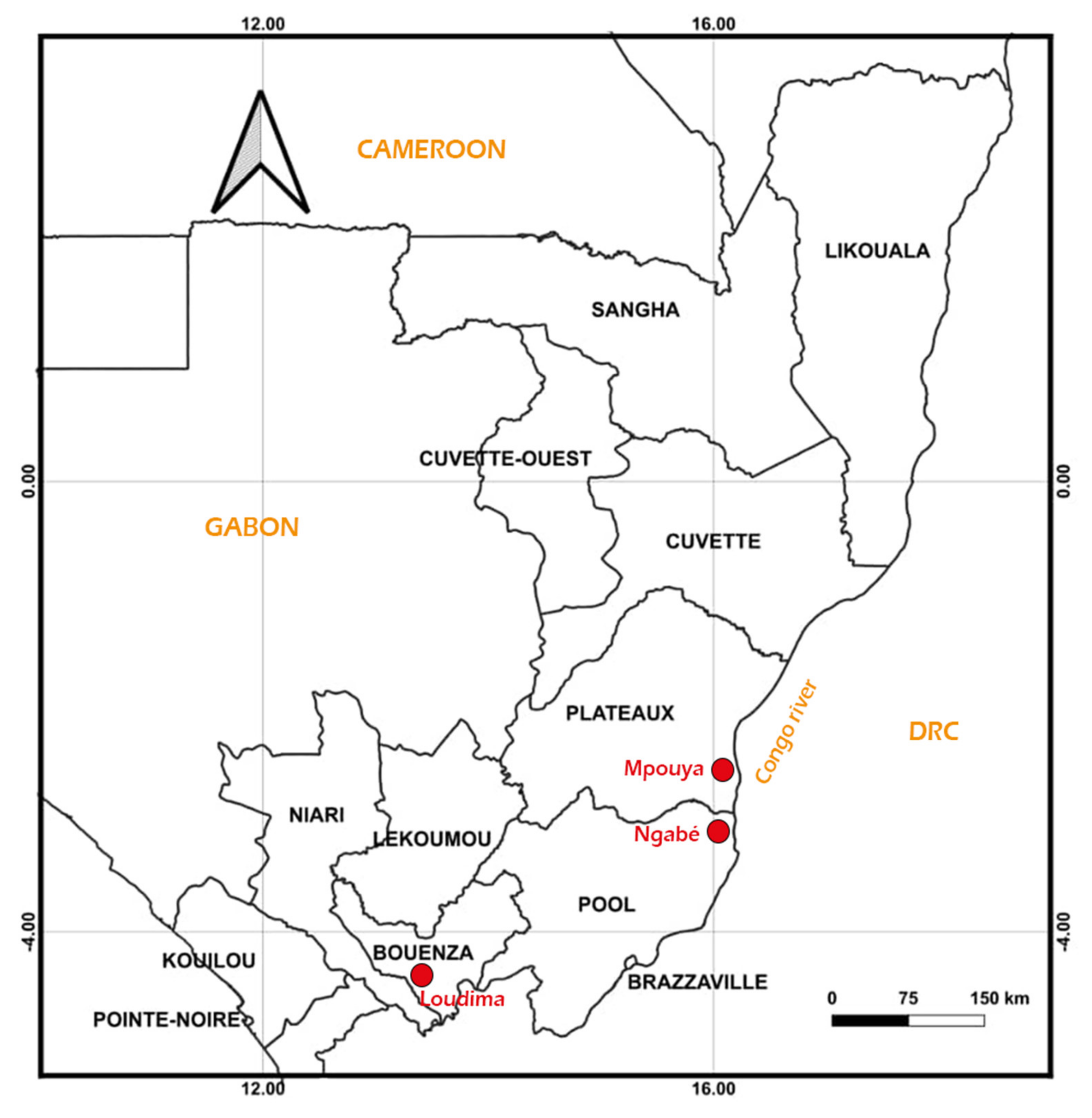
2.1. Study Sites
- − Mpouya (2°36′57″ S, 16°12′43″ E) is located in the Plateau administrative division about 300 km north of Brazzaville. The focus is composed of several villages bordering the Congo River. Its vegetation is essentially a savannah with a few tall trees, and the population lives on fishing and farming.
- − Ngabé (3°12′52″ S, 16°10′1″ E) is situated about 200 km north of Brazzaville in the Pool region. This focus is also composed of several villages bordering the Congo River. The vegetation is savannah and forest galleries along the edges of rivers. The population mainly practices farming, hunting and fishing for a living.
- − Loudima (4°6′45″ S, 13°3′30″ E) is located in the Bouenza division about 300 km south of Brazzaville. The vegetation is dominated by grass fields, and the main economic activity of the population is farming (Figure 1).
2.2. Tsetse Flies Sampling
2.3. DNA Extraction
2.4. Molecular Identification of Tsetse Fly Species
2.5. Molecular Identification of Different Trypanosome Species
2.6. Identification of Trypanosoma Congolense Forest and Trypanosoma Congolense Savannah
2.7. Data Analysis
3. Results
3.1. Entomological Surveys
3.2. Molecular Identification of Trypanosomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Büscher, P.; Cecchi, G.; Jamonneau, V.; Priotto, G. Human african trypanosomiasis. Lancet 2017, 390, 2397–2409. [Google Scholar] [CrossRef]
- World Health Organization. Accelerating Work to Overcome the Global Impact of Neglected Tropical Diseases: A Roadmap for Implementation; World Health Organization: Geneva, Switzerland, 2012. [Google Scholar]
- Aksoy, S. Control of tsetse flies and trypanosomes using molecular genetics. Vet. Parasitol. 2003, 115, 125–145. [Google Scholar] [CrossRef]
- Jamonneau, V.; Ravel, S.; Koffi, M.; Kaba, D.; Zeze, D.; Ndri, L.; Sane, B.; Coulibaly, B.; Cuny, G.; Solano, P. Mixed infections of trypanosomes in tsetse and pigs and their epidemiological significance in a sleeping sickness focus of Cote d’Ivoire. Parasitology 2004, 129, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Roditi, I.; Lehane, M.J. Interactions between trypanosomes and tsetse flies. Curr. Opin. Microbiol. 2008, 11, 345–351. [Google Scholar] [CrossRef]
- Franco, J.R.; Cecchi, G.; Paone, M.; Diarra, A.; Grout, L.; Kadima Ebeja, A.; Simarro, P.P.; Zhao, W.; Argaw, D. The elimination of human African trypanosomiasis: Achievements in relation to WHO road map targets for 2020. PLoS Negl. Trop. Dis. 2022, 16, e0010047. [Google Scholar] [CrossRef]
- Elimination of human African trypanosomiasis as a public health problem validated in Equatorial Guinea. Available online: https://www.who.int/news/item/20-06-2022-elimination-of-human-african-trypanosomiasis-as-a-public-health-problem-validated-in-equatorial-guinea (accessed on 20 June 2022).
- PNLTHA. Rapport Annuel Sur la Maladie du Sommeil; MinSanté: Brazzaville, Congo, 2019. [Google Scholar]
- PNLTHA. Rapport Annuel Sur la Maladie du Sommeil; MinSanté: Brazzaville, Congo, 2021. [Google Scholar]
- Hao, Z.; Kasumba, I.; Lehane, M.J.; Gibson, W.C.; Kwon, J.; Aksoy, S. Tsetse immune responses and trypanosome transmission: Implications for the development of tsetse-based strategies to reduce trypanosomiasis. Proc. Natl. Acad. Sci. USA 2001, 98, 12648–12653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouteux, J.P.; Noireau, F. Un nouvel écran-piège pour la lutte anti-tsétsé: Description et essais dans un foyer congolais de trypanosomiase humaine. Entomol. Exp. Appl. 1986, 41, 291–297. [Google Scholar] [CrossRef]
- Brunhes, J.; Cuisance, D.; Geoffroy, B.; Hervy, J.-P.; Lebbe, J. Logiciel D’identification Glossine Expert. Manuel Illustré D’utilisation. Les Glossines ou Mouches Tsé-Tsé; ORSTOM: Paris, France, 1994. [Google Scholar]
- Navajas, M.; Lagnel, J.; Gutierrez, J.; Boursot, P. Species-wide homogeneity of nuclear ribosomal ITS2 sequences in the spider mite Tetranychus urticae contrasts with extensive mitochondrial COI polymorphism. Heredity 1998, 80, 742–752. [Google Scholar] [CrossRef]
- Dyer, N.; Lawton, S.; Ravel, S.; Choi, K.; Lehane, M.; Robinson, A.; Okedi, L.; Hall, M.; Solano, P.; Donnelly, M. Molecular phylogenetics of tsetse flies (Diptera: Glossinidae) based on mitochondrial (COI, 16S, ND2) and nuclear ribosomal DNA sequences, with an emphasis on the palpalis group. Mol. Phylogenetics Evol. 2008, 49, 227–239. [Google Scholar] [CrossRef]
- Ravel, S.; Mediannikov, O.; Bossard, G.; Desquesnes, M.; Cuny, G.; Davoust, B. A study on African animal trypanosomosis in four areas of Senegal. Folia Parasitol. 2015, 62, 044. [Google Scholar] [CrossRef]
- Simo, G.; Fongho, P.; Farikou, O.; Ndjeuto-Tchouli, P.I.N.; Tchouomene-Labou, J.; Njiokou, F.; Asonganyi, T. Trypanosome infection rates in tsetse flies in the “silent” sleeping sickness focus of Bafia in the Centre Region in Cameroon. Parasites Vectors 2015, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masiga, D.K.; Smyth, A.J.; Hayes, P.; Bromidge, T.J.; Gibson, W.C. Sensitive detection of trypanosomes in tsetse flies by DNA amplification. Int. J. Parasitol. 1992, 22, 909–918. [Google Scholar] [CrossRef]
- Majiwa, P.; Thatthi, R.; Moloo, S.; Nyeko, J.; Otieno, L.; Maloo, S. Detection of trypanosome infections in the saliva of tsetse flies and buffy-coat samples from antigenaemic but aparasitaemic cattle. Parasitology 1994, 108, 313–322. [Google Scholar] [CrossRef]
- Schoonjans, F.; Zalata, A.; Depuydt, C.; Comhaire, F. MedCalc: A new computer program for medical statistics. Comput. Methods Programs Biomed. 1995, 48, 257–262. [Google Scholar] [CrossRef]
- PNLTHA. Rapport Annuel D’activités; MinSanté: Brazzaville, Congo, 2015. [Google Scholar]
- Maillot, L. Glossines d’Afrique Centrale. 2. Espèces rares ou peu répandues, mais pouvant jouer un rôle comme vecteur. Rev. D’élevage Et De Médecine Vétérinaire Des Pays Trop 1961, 14, 439–443. [Google Scholar] [CrossRef] [Green Version]
- Maillot, L. Les variétés de Glossina palpalis en Afrique Equatoriale Française. Bull. De La Société De Pathol. Exot. 1953, 46, 1066–1080. [Google Scholar]
- Taufflieb, R. Glossines et Élevage Dans la Région du Niari; 1964. Available online: https://horizon.documentation.ird.fr/exl-doc/pleins_textes/divers11-10/10310.pdf (accessed on 20 June 2022).
- Munang’andu, H.M.; Siamudaala, V.; Munyeme, M.; Nalubamba, K.S. A review of ecological factors associated with the epidemiology of wildlife trypanosomiasis in the Luangwa and Zambezi valley ecosystems of Zambia. Interdiscip. Perspect. Infect. Dis. 2012, 2012, 1–13. [Google Scholar] [CrossRef]
- Loudima. Available online: https://www.city-facts.com/loudima (accessed on 8 May 2020).
- Rodhain, F.; Perez, C. Precis D’entomologie Medicale et Veterinaire. In Notions D’epidemiologie Des Maladies a Vecteurs; FAO: Rome, Italy, 1985. [Google Scholar]
- Gouteux, J.; Nkouka, E.; Noireau, F.; Frézil, J.-L.; Sinda, D. Les glossines de l’agglomération brazzavilloise Congo. I. Répartition et importance des gîtes. Rev. D’élevage Et De Médecine Vétérinaire Des Pays Trop. 1986, 39, 355–362. [Google Scholar]
- Maillot, L.; Ceccaldi, J. Enquête sur les glossines dans la vallée du M’Filou au niveau de Gamaba à proximité de Brazzaville (janvier-avril 1956). Bull.-Inst. D’etudes Centrafr. 1956, 12, 201–208. [Google Scholar]
- Simo, G.; Silatsa, B.; Flobert, N.; Lutumba, P.; Mansinsa, P.; Madinga, J.; Manzambi, E.; De Deken, R.; Asonganyi, T. Identification of different trypanosome species in the mid-guts of tsetse flies of the Malanga (Kimpese) sleeping sickness focus of the Democratic Republic of Congo. Parasites Vectors 2012, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Morlais, I.; Grebaut, P.; Bodo, J.; Djoha, S.; Cuny, G.; Herder, S. Detection and identification of trypanosomes by polymerase chain reaction in wild tsetse flies in Cameroon. Acta Trop. 1998, 70, 109–117. [Google Scholar] [CrossRef]
- Djohan, V.; Kaba, D.; Rayaissé, J.-B.; Dayo, G.-K.; Coulibaly, B.; Salou, E.; Dofini, F.; Kouadio, A.D.M.K.; Menan, H.; Solano, P. Detection and identification of pathogenic trypanosome species in tsetse flies along the Comoé River in Côte d’Ivoire. Parasite 2015, 22, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehane, M.; Msangi, A.; Whitaker, C.; Lehane, S. Grouping of trypanosome species in mixed infections in Glossina pallidipes. Parasitology 2000, 120, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Signaboubo, D.; Payne, V.K.; Moussa, I.M.A.; Hassane, H.M.; Berger, P.; Kelm, S.; Simo, G. Diversity of tsetse flies and trypanosome species circulating in the area of Lake Iro in southeastern Chad. Parasites Vectors 2021, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ooi, C.-P.; Schuster, S.; Cren-Travaillé, C.; Bertiaux, E.; Cosson, A.; Goyard, S.; Perrot, S.; Rotureau, B. The cyclical development of Trypanosoma vivax in the tsetse fly involves an asymmetric division. Front. Cell. Infect. Microbiol. 2016, 6, 115. [Google Scholar] [CrossRef] [Green Version]
- Isaac, C.; Ciosi, M.; Hamilton, A.; Scullion, K.M.; Dede, P.; Igbinosa, I.B.; Nmorsi, O.P.G.; Masiga, D.; Turner, C.M.R. Molecular identification of different trypanosome species and subspecies in tsetse flies of northern Nigeria. Parasites Vectors 2016, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Tchouomene-Labou, J.; Nana-Djeunga, H.; Simo, G.; Njitchouang, G.R.; Cuny, G.; Asonganyi, T.; Njiokou, F. Spatial and temporal variations relevant to tsetse control in the Bipindi focus of southern Cameroon. Parasites Vectors 2013, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Franco, J.R.; Cecchi, G.; Priotto, G.; Paone, M.; Diarra, A.; Grout, L.; Simarro, P.P.; Zhao, W.; Argaw, D. Monitoring the elimination of human African trypanosomiasis at continental and country level: Update to 2018. PLoS Negl. Trop. Dis. 2020, 14, e0008261. [Google Scholar] [CrossRef]
- Paguem, A.; Abanda, B.; Ndjonka, D.; Weber, J.S.; Ngomtcho, S.C.H.; Manchang, K.T.; Adoulmoumini, M.; Eisenbarth, A.; Renz, A.; Kelm, S. Widespread co-endemicity of Trypanosoma species infecting cattle in the Sudano-Sahelian and Guinea Savannah zones of Cameroon. BMC Vet. Res. 2019, 15, 1–15. [Google Scholar] [CrossRef]
- Magona, J.; Walubengo, J.; Kabi, F.; Odimim, J.; Ocaido, M. Mapping the distribution of tsetse and animal trypanosomosis in selected districts of Uganda to facilitate targeting control measures. Bull. Anim. Health Prod. Afr. 2017, 65, 459–469. [Google Scholar]
- Meyer, A.; Holt, H.R.; Selby, R.; Guitian, J. Past and ongoing tsetse and animal trypanosomiasis control operations in five African countries: A systematic review. PLoS Negl. Trop. Dis. 2016, 10, e0005247. [Google Scholar]


{kind=link}
| Foci | No. Tsetse fly | No of Teneral Flies (%) | No. Trap | ADT | No of Males (%) | No of Females (%) | Sex Ratio |
|---|---|---|---|---|---|---|---|
| Mpouya | 5 | 0 (0) | 38 | 0.043 | 0 (0) | 5 (100) | 0 |
| Ngabé | 179 | 5 (0.03) | 81 | 0.73 | 58 (32.40) | 121 (67.60) | 0.5 |
| Loudima | 1107 | 11 (0.01) | 132 | 2.79 | 462 (41.73) | 645 (58.27) | 0.7 |
| p-value | 0.001 |
| HAT Foci | Tsetse Fly Species | Total | ||
|---|---|---|---|---|
| G.p. palpalis | G.f. quazensis | G.f. congolensis | ||
| Mpouya | − | 5 | − | 5 |
| Ngabé | − | 178 | 1 | 179 |
| Loudima | 1107 | − | − | 1107 |
| Total | 1107 | 183 | 1 | 1291 |
| Foci | No. Tested | T. c. s. (%) | T. c. f. (%) | T. vivax (%) | T. b. (s.l.) (%) | Coinfection T.b + T.c.s | Coinfection T.b + T.c.f. | Unknown Species |
|---|---|---|---|---|---|---|---|---|
| Loudima | 1103 | 95 (8.61) | 21 (1.9) | 11 (1) | 82 (7.43) | 49 (85.96) | 8 (14.04) | 4 (0.36) |
| Mpouya | 5 | 0 | 0 | 0 | 1 (20) | 0 | 0 | 0 |
| Ngabé | 157 | 5 (3.18) | 1 (0.64) | 4 (2.55) | 0 | 0 | 0 | 0 |
| Total | 1265 | 100 (7.91) | 22 (1.74) | 15 (1.19) | 83 (6.56) | 49 (3.87) | 8 (0.63) | 4 (0.16) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bemba, I.; Lenga, A.; Awono-Ambene, H.P.; Antonio-Nkondjio, C. Tsetse Flies Infected with Trypanosomes in Three Active Human African Trypanosomiasis Foci of the Republic of Congo. Pathogens 2022, 11, 1275. https://doi.org/10.3390/pathogens11111275
Bemba I, Lenga A, Awono-Ambene HP, Antonio-Nkondjio C. Tsetse Flies Infected with Trypanosomes in Three Active Human African Trypanosomiasis Foci of the Republic of Congo. Pathogens. 2022; 11(11):1275. https://doi.org/10.3390/pathogens11111275
Chicago/Turabian StyleBemba, Irina, Arsene Lenga, Herman Parfait Awono-Ambene, and Christophe Antonio-Nkondjio. 2022. "Tsetse Flies Infected with Trypanosomes in Three Active Human African Trypanosomiasis Foci of the Republic of Congo" Pathogens 11, no. 11: 1275. https://doi.org/10.3390/pathogens11111275