
Alterations in Genes rib, scpB and Pilus Island Decrease the Prevalence of Predominant Serotype V, Not III and VI, of Streptococcus agalactiae from 2008 to 2012
Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Susceptibility Analysis
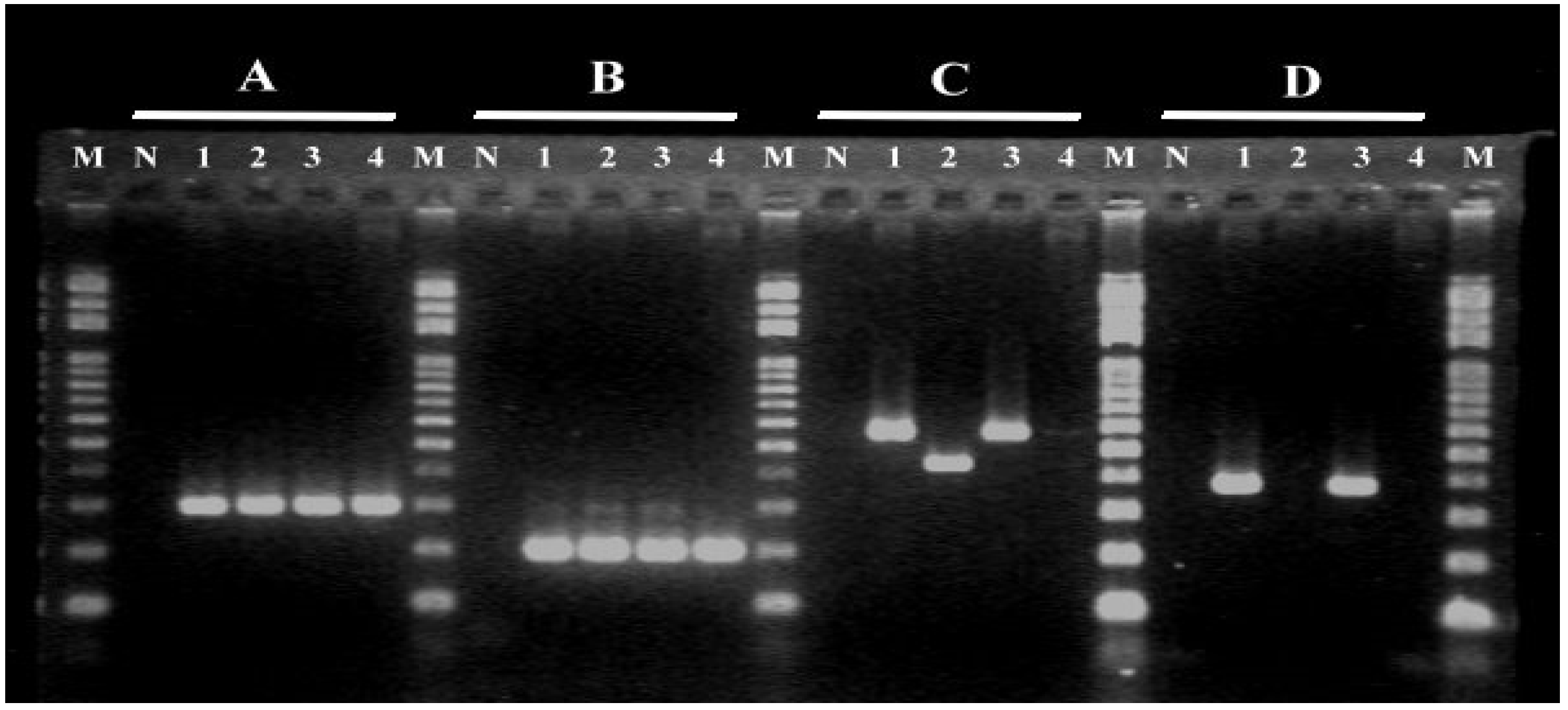
2.2. ST-17 and Pilus Island Gene Analysis
2.3. Virulence Gene Analysis
2.4. Amino Acid Comparison and Conserved Domain (CDD) Analysis of scpB Protein
2.5. Phylogenetic Analysis
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Antimicrobial Susceptibility
4.3. PCR Identification of ST17 Strains
4.4. Pilus Island Genes, Virulence Genes and Sequencing
4.5. Amino Acid Comparison and Conserved Domain Analysis of scpB Protein
4.6. Phylogenetic Analysis of scpB Gene
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.-H.; Lu, C.-C.; Chiu, C.-H.; Wang, M.-H.; Yang, T.-H.; Chu, C. Genetically diverse serotypes III and VI substitute major clonal disseminated serotypes Ib and V as prevalent serotypes of Streptococcus agalactiae from 2007 to 2012. J. Microbiol. Immunol. Infect. 2016, 49, 672–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobadilla, F.J.; Novosak, M.G.; Cortese, I.J.; Delgado, O.D.; Laczeski, M.E. Prevalence, serotypes and virulence genes of Streptococcus agalactiae isolated from pregnant women with 35–37 weeks of gestation. BMC Infect. Dis. 2021, 21, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Teatero, S.; Ferrieri, P.; Martin, I.; Demczuk, W.; McGeer, A.; Fittipaldi, N. Serotype distribution, population structure, and antimicrobial resistance of group B Streptococcus strains recovered from colonized pregnant women. J. Clin. Microbiol. 2017, 55, 412–422. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Li, S.; Li, L.; Wang, X.; Yao, Z.; Ye, X. Alarming regional differences in prevalence and antimicrobial susceptibility of group B streptococci in pregnant women: A systematic review and meta-analysis. J. Glob. Antimicrob. Resist. 2016, 7, 169–177. [Google Scholar] [CrossRef]
- Watkins, L.K.F.; McGee, L.; Schrag, S.J.; Beall, B.; Jain, J.H.; Pondo, T.; Farley, M.M.; Harrison, L.H.; Zansky, S.M.; Baumbach, J. Epidemiology of invasive group B streptococcal infections among nonpregnant adults in the United States, 2008-2016. JAMA Intern. Med. 2019, 179, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Poyart, C.; Tazi, A.; Réglier-Poupet, H.; Billoët, A.; Tavares, N.; Raymond, J.; Trieu-Cuot, P. Multiplex PCR assay for rapid and accurate capsular typing of group B streptococci. J. Clin. Microbiol. 2007, 45, 1985–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, N.; Bohnsack, J.F.; Takahashi, S.; Oliver, K.A.; Chan, M.-S.; Kunst, F.; Glaser, P.; Rusniok, C.; Crook, D.W.; Harding, R.M. Multilocus sequence typing system for group B streptococcus. J. Clin. Microbiol. 2003, 41, 2530–2536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Cao, X.; Li, S.; Ou, Q.; Lin, D.; Yao, Z.; Chen, S.; Wen, G.; Ye, X. Phenotypic and molecular characterization of Streptococcus agalactiae colonized in Chinese pregnant women: Predominance of ST19/III and ST17/III. Res. Microbiol. 2018, 169, 101–107. [Google Scholar] [CrossRef]
- Lamy, M.-C.; Dramsi, S.; Billoët, A.; Réglier-Poupet, H.; Tazi, A.; Raymond, J.; Guérin, F.; Couvé, E.; Kunst, F.; Glaser, P. Rapid detection of the “highly virulent” group B Streptococcus ST-17 clone. Microbes Infect. 2006, 8, 1714–1722. [Google Scholar] [CrossRef]
- Springman, A.C.; Lacher, D.W.; Waymire, E.A.; Wengert, S.L.; Singh, P.; Zadoks, R.N.; Davies, H.; Manning, S.D. Pilus distribution among lineages of group b streptococcus: An evolutionary and clinical perspective. BMC Microbiol. 2014, 14, 1–11. [Google Scholar] [CrossRef]
- Lazzarin, M.; Mu, R.; Fabbrini, M.; Ghezzo, C.; Rinaudo, C.D.; Doran, K.S.; Margarit, I. Contribution of pilus type 2b to invasive disease caused by a Streptococcus agalactiae ST-17 strain. BMC Microbiol. 2017, 17, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudzana, R.; Mavenyengwa, R.T.; Gudza-Mugabe, M. Analysis of virulence factors and antibiotic resistance genes in group B streptococcus from clinical samples. BMC Infect. Dis. 2021, 21, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shome, B.R.; Bhuvana, M.; Mitra, S.D.; Krithiga, N.; Shome, R.; Velu, D.; Banerjee, A.; Barbuddhe, S.B.; Prabhudas, K.; Rahman, H. Molecular characterization of Streptococcus agalactiae and Streptococcus uberis isolates from bovine milk. Trop. Anim. Health Prod. 2012, 44, 1981–1992. [Google Scholar] [CrossRef] [PubMed]
- Koroleva, I.V.; Efstratiou, A.; Suvorov, A.N. Structural heterogeneity of the streptococcal C5a peptidase gene in Streptococcus pyogenes. J. Bacteriol. 2002, 184, 6384–6386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuzeville, S.; Puymege, A.; Madec, J.-Y.; Haenni, M.; Payot, S. Characterization of a new CAMP factor carried by an integrative and conjugative element in Streptococcus agalactiae and spreading in Streptococci. PLoS ONE 2012, 7, e48918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fröhlicher, S.; Reichen-Fahrni, G.; Müller, A.M.; Surbek, D.; Droz, S.C.; Spellerberg, B.; Sendi, P. Serotype distribution and antimicrobial susceptibility of group B streptococci in pregnant women: Results from a Swiss tertiary centre. Swiss Med. Wkly. 2014, 144, w13935. [Google Scholar] [CrossRef] [Green Version]
- Uh, Y.; Kim, H.Y.; Jang, I.H.; Hwang, G.Y.; Yoon, K.J. Correlation of serotypes and genotypes of macrolide-resistant Streptococcus agalactiae. Yonsei Med. J. 2005, 46, 480–483. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Ma, L.; Zhu, L.; Zhou, X.-H.; Xu, L.-J.; Guo, C.; Meng, J.-H.; Zhang, X.-H.; Liu, Q.-H.; Huang, R. Molecular characterization of pathogenic group B streptococcus from a tertiary hospital in Shanxi, China: High incidence of sequence type 10 strains in infants/pregnant women. J. Microbiol. Immunol. Infect. 2021, 54, 1094–1100. [Google Scholar] [CrossRef]
- do Nascimento, C.S.; Dos Santos, N.F.; Ferreira, R.C.; Taddei, C.R. Streptococcus agalactiae in pregnant women in Brazil: Prevalence, serotypes, and antibiotic resistance. Braz. J. Microbiol. 2019, 50, 943–952. [Google Scholar] [CrossRef]
- Nabavinia, M.; Khalili, M.B.; Sadeh, M.; Eslami, G.; Vakili, M.; Azartoos, N.; Mojibiyan, M. Distribution of Pilus island and antibiotic resistance genes in Streptococcus agalactiae obtained from vagina of pregnant women in Yazd, Iran. Iran, J. Microbiol. 2020, 12, 411. [Google Scholar] [CrossRef]
- Gajic, I.; Plainvert, C.; Kekic, D.; Dmytruk, N.; Mijac, V.; Tazi, A.; Glaser, P.; Ranin, L.; Poyart, C.; Opavski, N. Molecular epidemiology of invasive and non-invasive group B Streptococcus circulating in Serbia. Int. J. Med. Microbiol. 2019, 309, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Poyart, C.; Réglier-Poupet, H.; Tazi, A.; Billoët, A.; Dmytruk, N.; Bidet, P.; Bingen, E.; Raymond, J.; Trieu-Cuot, P. Invasive group B streptococcal infections in infants, France. Emerging Infect. Dis. 2008, 14, 1647. [Google Scholar]
- Li, J.; Ji, W.; Gao, K.; Zhou, H.; Zhang, L.; Mu, X.; Yuan, C.; Guan, X.; Deng, Q.; Zhang, L. Molecular characteristics of group B Streptococcus isolates from infants in southern mainland China. BMC Infect. Dis. 2019, 19, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rinaudo, C.D.; Rosini, R.; Galeotti, C.L.; Berti, F.; Necchi, F.; Reguzzi, V.; Ghezzo, C.; Telford, J.L.; Grandi, G.; Maione, D. Specific involvement of pilus type 2a in biofilm formation in group B Streptococcus. PLoS ONE 2010, 5, e9216. [Google Scholar] [CrossRef]
- Gizachew, M.; Tiruneh, M.; Moges, F.; Adefris, M.; Tigabu, Z.; Tessema, B. Molecular characterization of Streptococcus agalactiae isolated from pregnant women and newborns at the University of Gondar Comprehensive Specialized Hospital, Northwest Ethiopia. BMC Infect. Dis. 2020, 20, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, G.; Stålhammar-Carlemalm, M.; Areschoug, T. Surface proteins of Streptococcus agalactiae and related proteins in other bacterial pathogens. Clin. Microbiol. Rev. 2005, 18, 102–127. [Google Scholar] [CrossRef] [Green Version]
- Usein, C.-R.; Grigore, L.; Georgescu, R.; Cristea, V.; Bãltoiu, M.; Strãuţ, M. Molecular characterization of adult-colonizing Streptococcus agalactiae from an area-based surveillance study in Romania. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 2301–2310. [Google Scholar] [CrossRef]
- Dobrut, A.; Brzychczy-Włoch, M. Immunogenic Proteins of Group B Streptococcus—Potential Antigens in Immunodiagnostic Assay for GBS Detection. Pathogens 2021, 11, 43. [Google Scholar] [CrossRef]
- Kaczorek, E.; Małaczewska, J.; Wójcik, R.; Siwicki, A.K. Biofilm production and other virulence factors in Streptococcus spp. isolated from clinical cases of bovine mastitis in Poland. BMC Vet. Res. 2017, 13, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Tulyaprawat, O.; Pharkjaksu, S.; Shrestha, R.K.; Ngamskulrungroj, P. Emergence of multi-drug resistance and its association with uncommon serotypes of Streptococcus agalactiae isolated from non-neonatal patients in Thailand. Front. Microbiol. 2021, 12, 719353. [Google Scholar] [CrossRef]
- Springman, A.C.; Lacher, D.W.; Wu, G.; Milton, N.; Whittam, T.S.; Davies, H.D.; Manning, S.D. Selection, recombination, and virulence gene diversity among group B streptococcal genotypes. J. Bacteriol. 2009, 191, 5419–5427. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Insitute. Performance standards for antimicrobial susceptibility tests, 27th ed.; Clinical and Laboratory Standards Insitute: Wayne, PA, USA, 2017. [Google Scholar]
- Martins, E.R.; Melo-Cristino, J.; Ramirez, M. Evidence for rare capsular switching in Streptococcus agalactiae. J. Bacteriol. 2010, 192, 1361–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohnsack, J.F.; Takahashi, S.; Hammitt, L.; Miller, D.V.; Aly, A.A.; Adderson, E.E. Genetic polymorphisms of group B Streptococcus scpB alter functional activity of a cell-associated peptidase that inactivates C5a. Infect. Immun. 2000, 68, 5018–5025. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Antimicrobial Resistances | Antimicrobial Agents | Serotype III (%) | Serotype V (%) | Serotype VI (%) | Total (%) | Sum (%) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2008 | 2012 | 2008 | 2012 | 2008 | 2012 | 2008 | 2012 | |||
| 0 | - | 8 (47.1) | 8 (44.4) | 21 (36.2) | 7 (46.7) | 10 (58.8) | 14 (70) | 39 (42.4) | 29 (54.7) | 68 (46.9) |
| 1 | AZM | 1 (1.7) | 1 (1.1) | 0 | 1 (0.7) | |||||
| CC | 1 (5.9) | 2 (11.1) | 1 (1.1) | 2 (3.8) | 3 (2.1) | |||||
| 2 | E AZM | 3 (17.6) | 1 (5.6) | 1 (5) | 3 (3.3) | 2 (3.8) | 5 (3.4) | |||
| AZM CC | 1 (1.7) | 1 (1.1) | 0 | 1 (0.7) | ||||||
| 3 | E AZM CC | 5 (29.4) | 7 (38.9) | 35 (60.3) | 8 (53.3) | 7 (41.2) | 5 (25) | 47 (51.1) | 20 (37.7) | 67 (46.2) |
| Total | 17 (18.5) | 18 (34) | 58 (63) | 15 (28.3) | 17 (18.5) | 20 (37.7) | 92 | 53 | 145 | |
| Total (N (%)) | ST-17 (PCR/MLST) | PI gene (N (%)) | Serotype III (N (%)) | Serotype V (N (%)) | Serotype VI (N (%)) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| PI-1 | PI-2a | PI-2b | 2008 | 2012 | 2008 | 2012 | 2008 | 2012 | ||
| 10 (6.9) | +/+ | + | + | 6 (35.3) | 4 (22.2) | |||||
| 2 (1.4) | + | 2 (11.1) | ||||||||
| 103 (71) | −/− | + | + | 11 (64.7) | 10 (55.6) | 43 (74.1) | 11 (73.3) | 16 (94.1) | 12 (60) | |
| 24 (16.6) | + | 1 (5.6) | 10 (17.2) | 4 (26.7) | 1 (5.9) | 8 (40) | ||||
| 6 (4.1) | + | 1 (5.6) | 5 (8.6) | |||||||
| 145 | 12 (8.3) | 113 (77.9) | 127 (87.6) | 18 (12.4) | 17 | 18 | 58 | 15 | 17 | 20 |
| Total N (%) | Virulence gene (N (%)) | Serotype III (N (%)) | Serotype V (N (%)) | Serotype VI (N (%)) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| pavA | cfb | scpB-1 | scpB-2 | rib | 2008 | 2012 | 2008 | 2012 | 2008 | 2012 | |
| 43 (29.7) | + | + | + | + | 15 (88.2) | 16 (88.9) | 1 (1.7) | 3 (20) | 2 (11.8) | 6 (30) | |
| 7 (4.8) | + | + | + | + | 2 (3.4) | 5 (33.3) | |||||
| 44 (30.3) | + | + | + | 2 (11.8) | 2 (11.1) | 9 (15.5) | 2 (13.3) | 15 (88.2) | 14 (70) | ||
| 47 (32.4) | + | + | + | 42 (72.4) | 5 (33.3) | ||||||
| 4 (2.8) | + | + | 4 (6.9) | ||||||||
| 145 | 145 (100) | 145 (100) | 87 (60) | 54 (37.2) | 50 (34.5) | 17 | 18 | 58 | 15 | 17 | 20 |
| Country | Taiwan (this study) | China | China | USA | Brazil | Switzerland | Iran | South Korea | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | 2008 + 2012 | 2015 | 2017–2019 | 2008–2016 | 2008–2018 | 2009–2010 | 2015–2016 | 1990–2002 | ||||||||
| Serotype | All | III | V | VI | All | All | All | All | III | V | VI | All | All | III | V | VI |
| Azithromycin | 51 | 45.7 | 61.6 | 35.1 | ||||||||||||
| Clindamycin | 49 | 42.9 | 60.3 | 32.4 | 66.4 | 66.7 | 43.2 | 2–16.7 | 15 | 24.7 | 0 | 21.1 | 35 | 29 | 85 | 5 |
| Erythromycin | 49.7 | 45.7 | 58.9 | 35.1 | 72.1 | 70.3 | 54.8 | 4–14 | 15 | 25.8 | 0 | 14 | 30 | 23 | 85 | 21 |
| Reference | [8] | [18] | [5] | [19] | [16] | [20] | [17] | |||||||||
| Gene | Primers | Sequences (5′ to 3′) | Product Size (bp) | Reference |
|---|---|---|---|---|
| ST-17 | ST-17S | ATACAAATTCTGCTGACTACCG | 210 | [9] |
| ST-17AS | TTAAATCCTTCCTGACCATTCC | |||
| adhP | adhP-F | GTTGGTCATGGTGAAGCACT | 672 | [7] |
| adhP-R | ACTGTACCTCCAGCACGAAC | |||
| pheS | pheS-F | GATTAAGGAGTAGTGGCACG | 723 | |
| pheS-R | TTGAGATCGCCCATTGAAAT | |||
| atr | atr-F | CGATTCTCTCAGCTTTGTTA | 627 | |
| atr-R | AAGAAATCTCTTGTGCGGAT | |||
| glnA | glnA-F | CCGGCTACAGATGAACAATT | 589 | |
| glnA-R | CTGATAATTGCCATTCCACG | |||
| sdhA | sdhA-F | AGAGCAAGCTAATAGCCAAC | 646 | |
| sdhA-R | ATATCAGCAGCAACAAGTGC | |||
| glcK | glcK-F | CTCGGAGGAACGACCATTAA | 607 | |
| glcK-R | CTTGTAACAGTATCACCGTT | |||
| tkt | tkt-F | CCAGGCTTTGATTTAGTTGA | 859 | |
| tkt-R | AATAGCTTGTTGGCTTGAAA | |||
| PI-1 | PI1-UP | GGTCGTCGATGCTCTGGATTC | 881 | [33] |
| PI1-DN | GTTGCCCAGTAACAGCTTCTCC | |||
| PI-2a | PI2a-UP | CTATGACACTAATGGTAGAAC | 575 | |
| PI2a-DN | CACCTGCAATAGACATCATAG | |||
| PI-2b | PI2b-UP | ACACGACTATGCCTCCTCATG | 721 | |
| PI2b-DN | TCTCCTACTGGAATAATGACAG | |||
| PI1_all | PI1_all-UP | ACCTATGTTGCTGATTCGGCTGAAAATG | 684 * | |
| PI1_all-DN | TACGGACACTTTCTAGTGCCTTTGGATC | |||
| pavA | pavA-F | TTCCCATGATTTCAACAACAAG | 288 | [30] |
| pavA-R | AACCTTTTGACCATGAATTGGTA | |||
| cfb | cfb-F | ATGGGATTTGGGATAACTAAGCTAG | 193 | |
| cfb-R | AGCGTGTATTCCAGATTTCCTTAT | |||
| scpB | scpB-F | AGTTGCTTCTTACAGCCCAGA | 567 | |
| scpB-R | GGCGCAGACATACTAGTTCCA | |||
| rib | rib-F | CAGGAAGTGCTGTTACGTTAAAC | 369 | |
| rib-R | CGTCCCATTTAGGGTCTTCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, I.-A.; Su, Y.; Wang, Y.-H.; Chu, C. Alterations in Genes rib, scpB and Pilus Island Decrease the Prevalence of Predominant Serotype V, Not III and VI, of Streptococcus agalactiae from 2008 to 2012. Pathogens 2022, 11, 1145. https://doi.org/10.3390/pathogens11101145
Tsai I-A, Su Y, Wang Y-H, Chu C. Alterations in Genes rib, scpB and Pilus Island Decrease the Prevalence of Predominant Serotype V, Not III and VI, of Streptococcus agalactiae from 2008 to 2012. Pathogens. 2022; 11(10):1145. https://doi.org/10.3390/pathogens11101145
Chicago/Turabian StyleTsai, I-An, Yaochi Su, Ying-Hsiang Wang, and Chishih Chu. 2022. "Alterations in Genes rib, scpB and Pilus Island Decrease the Prevalence of Predominant Serotype V, Not III and VI, of Streptococcus agalactiae from 2008 to 2012" Pathogens 11, no. 10: 1145. https://doi.org/10.3390/pathogens11101145