
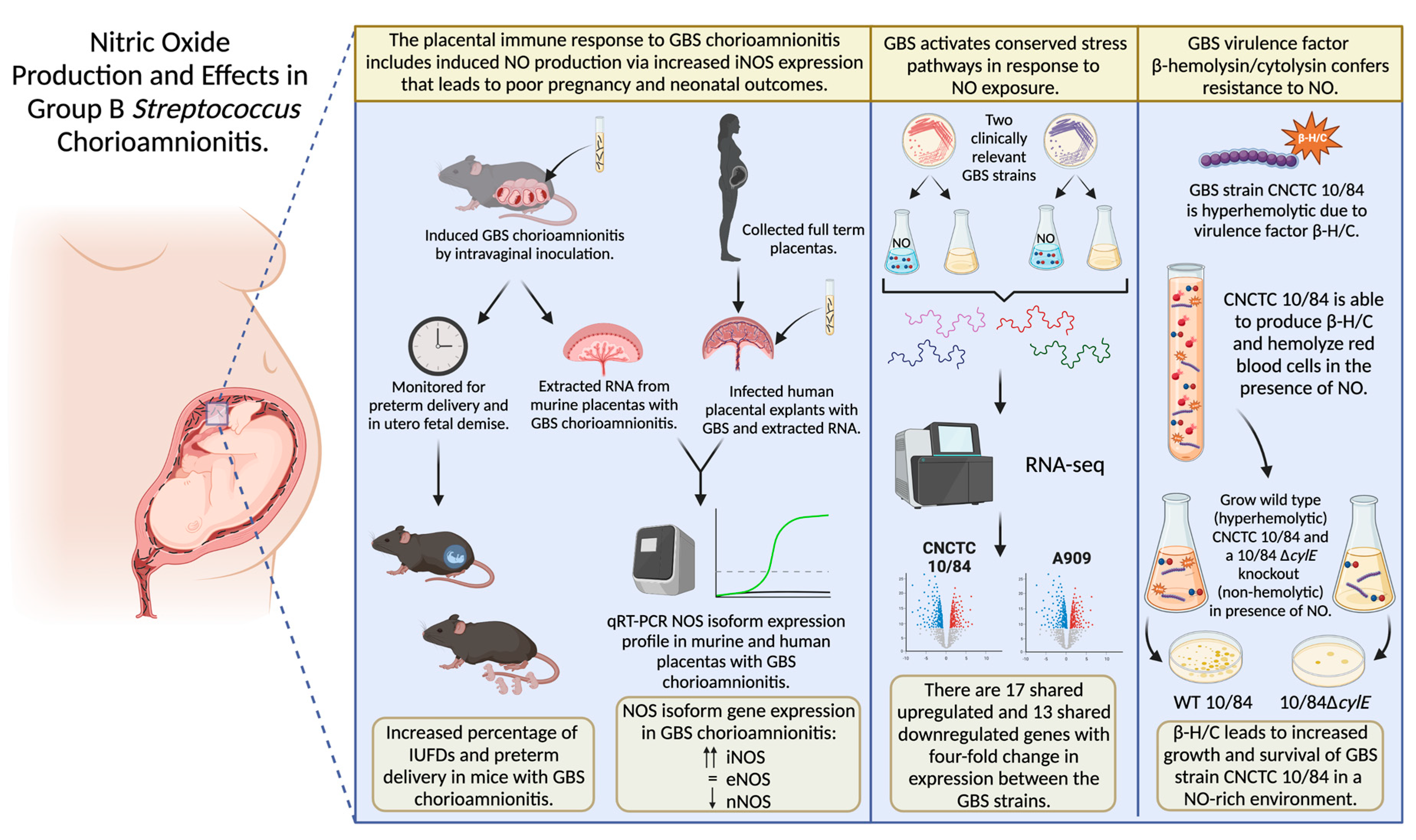
Nitric Oxide Production and Effects in Group B Streptococcus Chorioamnionitis
 ,
,
Abstract
:
1. Introduction
2. Methods
2.1. Ethics Statement
2.2. Statistical Analyses
2.3. Bacterial Strains and Growth Conditions
2.4. Murine GBS Chorioamnionitis Model
2.5. Human Placental Collection
2.6. Human Placental GBS Infection and RNA Purification
2.7. Quantification of Placental Gene Expression
2.8. Histological Preparation and Staining of Murine Placenta
2.9. Growth Curve Analysis
2.10. Hemolysis Assays
2.11. GBS Survival Assay
2.12. Whole-Genome RNA-Seq Transcriptomic Analyses
2.13. RT-qPCR Validation of RNA-Seq Up- and Downregulated Genes
3. Results
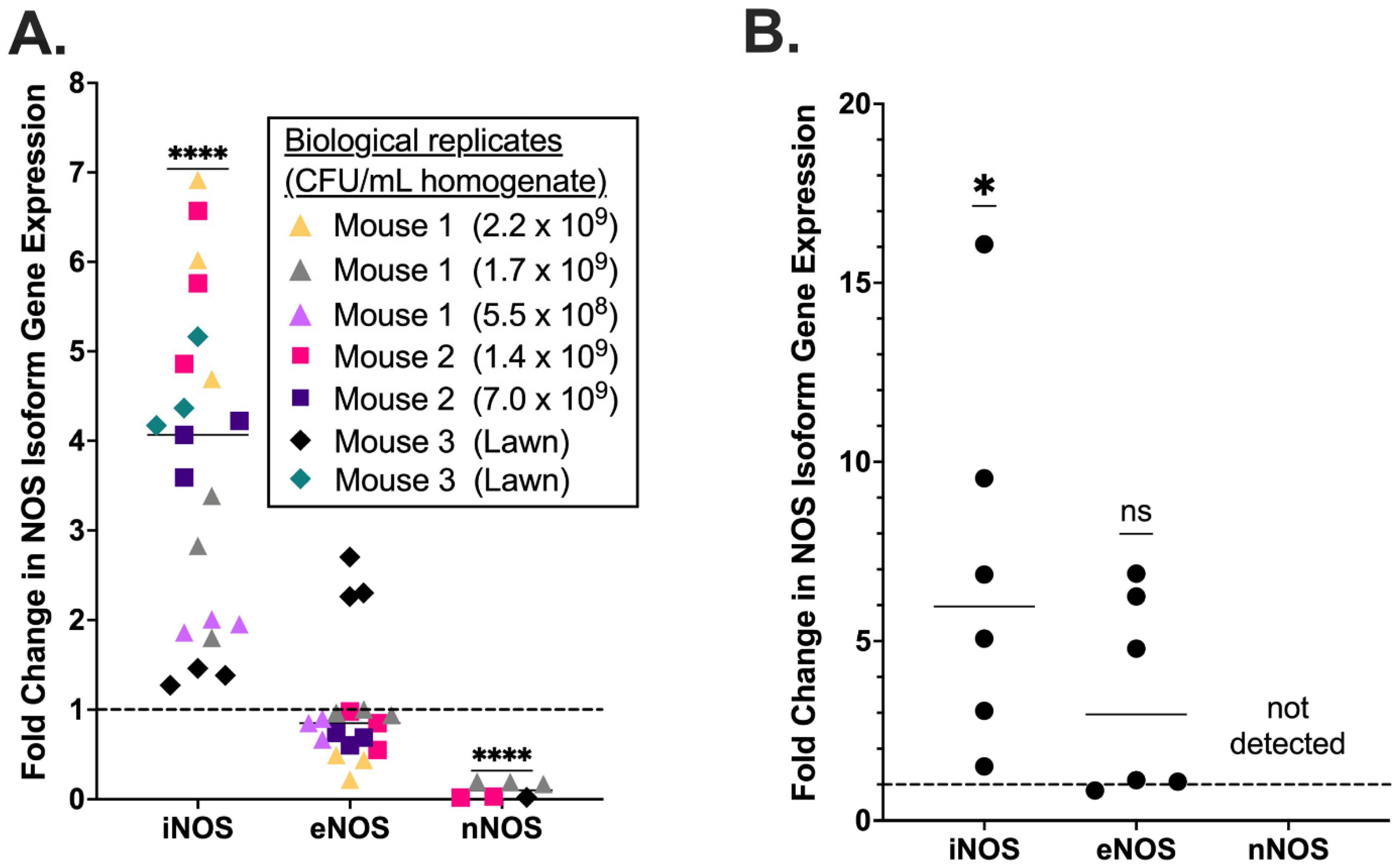
3.1. Placental iNOS Isoform Expression Is Increased in a Murine Model of GBS Chorioamnionitis
3.2. Ex Vivo Human Placental Samples Infected with GBS Show Increased iNOS Expression
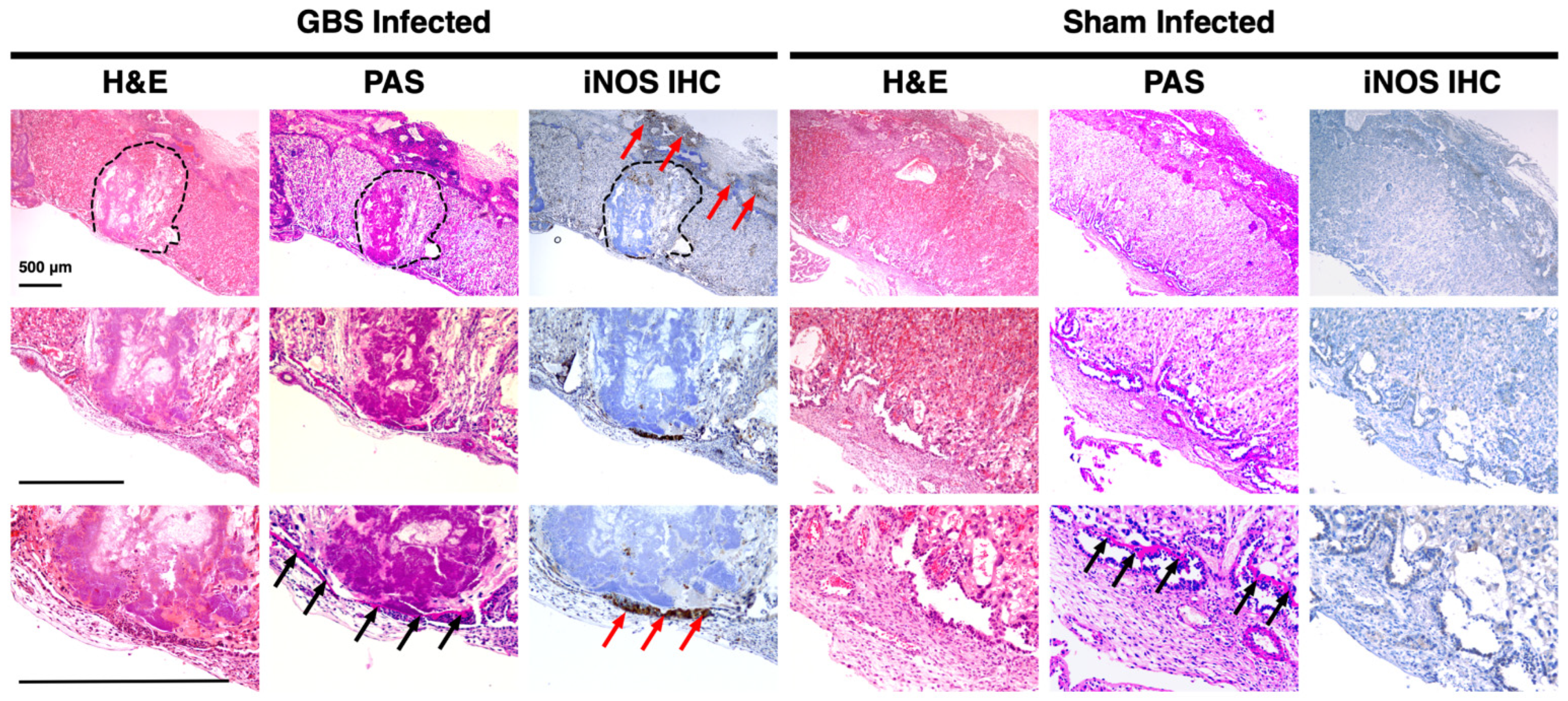
3.3. iNOS Isoform Expression Localizes to the Junctional Zone of GBS-Infected Murine Placentas
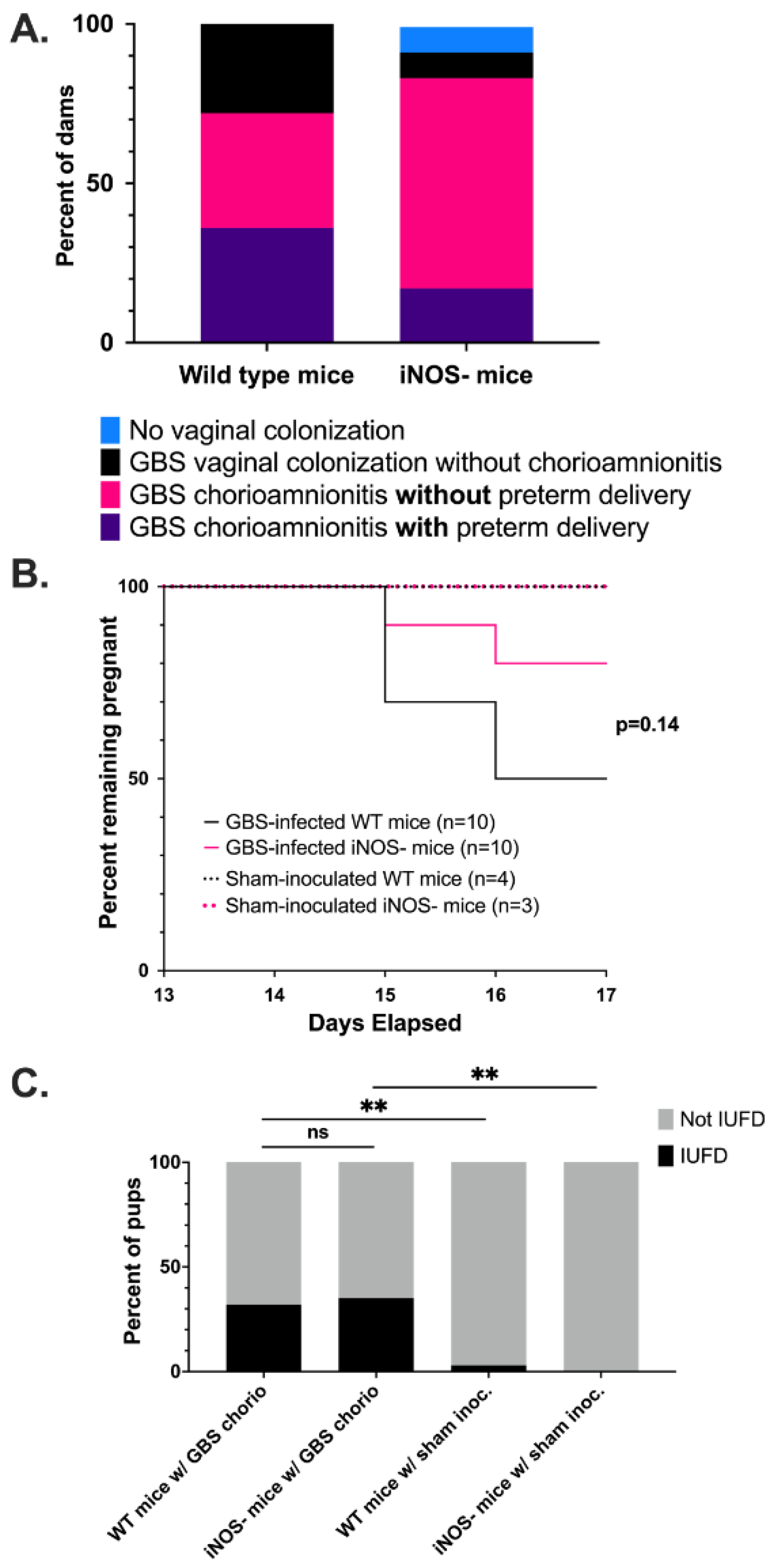
3.4. Wild Type Dams with GBS Chorioamnionitis Have Increased Rates of Preterm Delivery
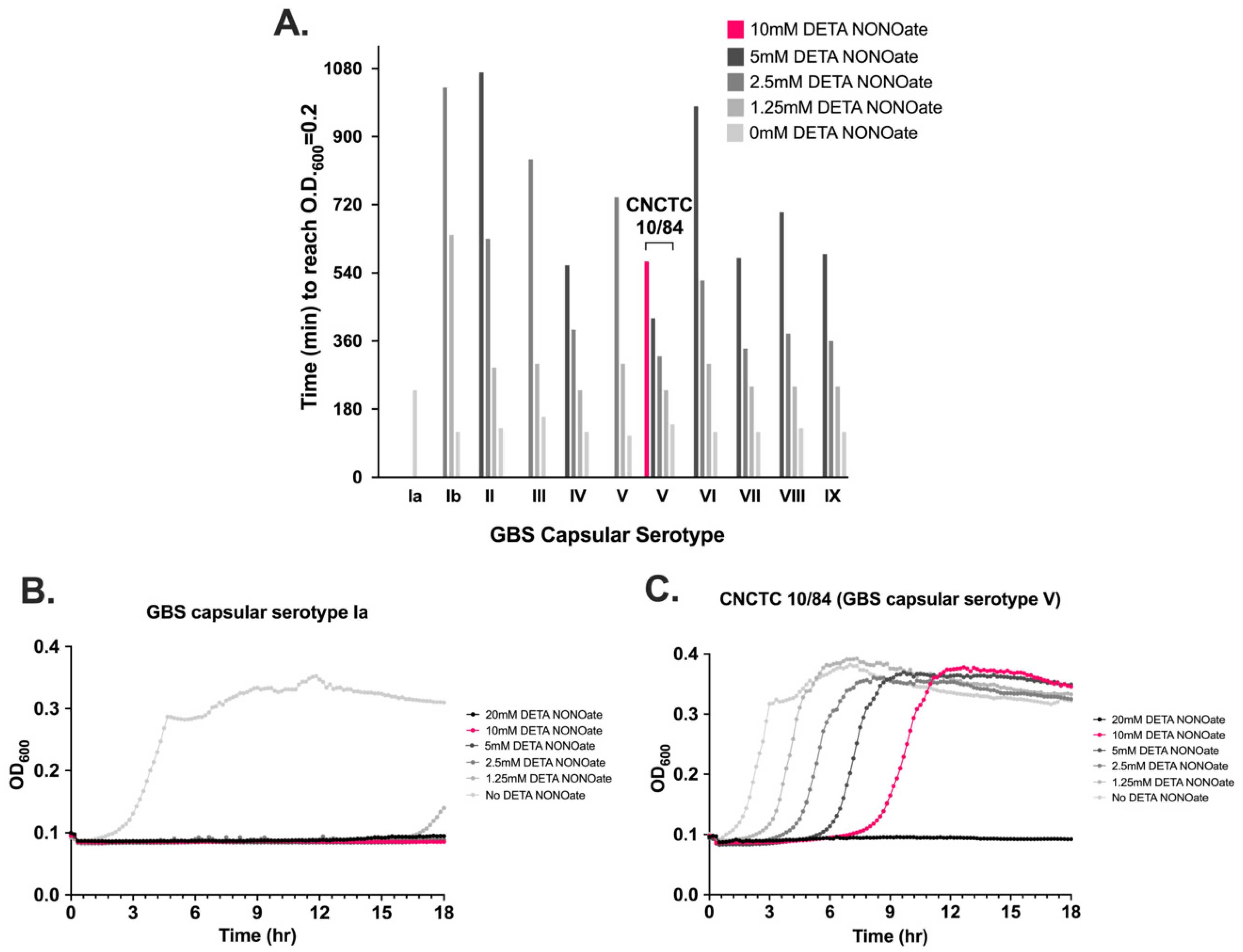
3.5. NO Exposure Results in Dose-Dependent GBS Growth Inhibition That Varies by Serovar
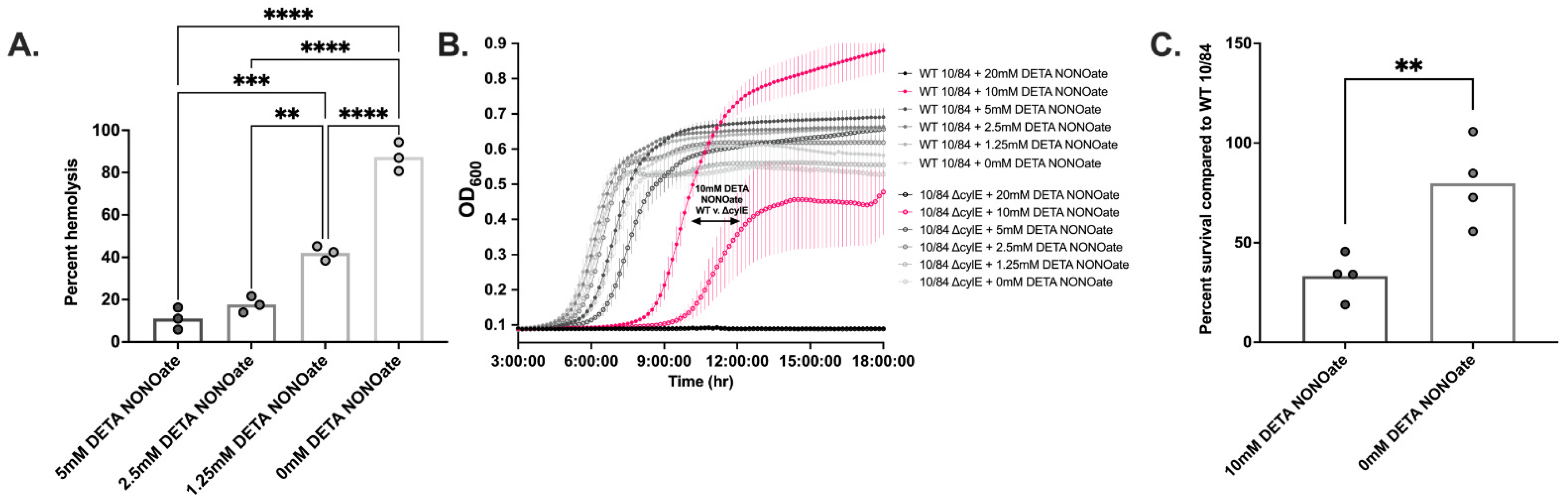
3.6. β-Hemolysin/Cytolysin Confers a Growth and Survival Advantage to GBS in a NO-Rich Environment
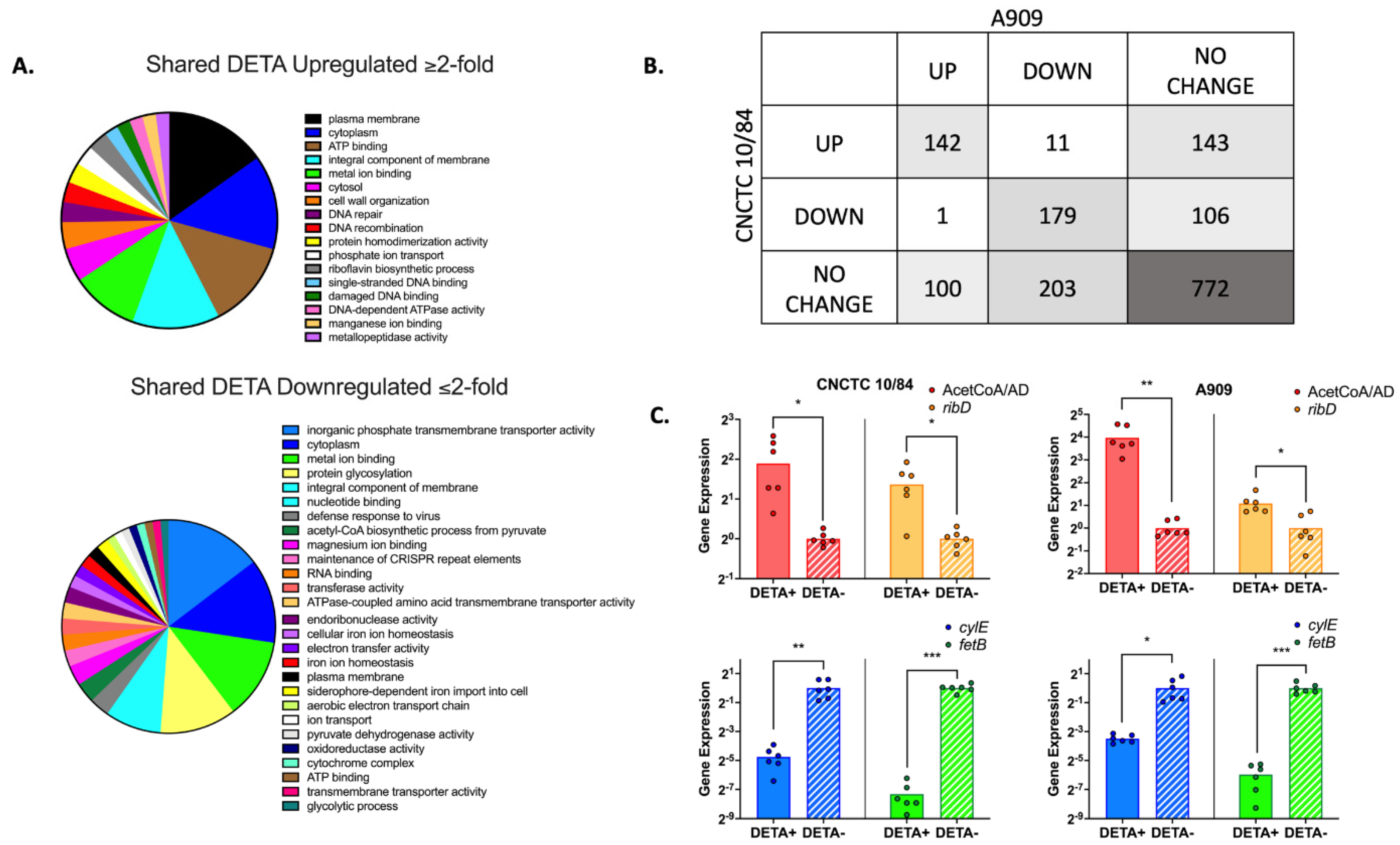
3.7. GBS Displays a Broad Transcriptomic Response to NO Exposure
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stoll, B.J. Early-Onset Neonatal Sepsis: A Continuing Problem in Need of Novel Prevention Strategies. Pediatrics 2016, 138, e20163038. [Google Scholar] [CrossRef] [PubMed]
- Nan, C.; Dangor, Z.; Cutland, C.; Edwards, M.; Madhi, S.; Cunnington, M. Maternal Group B Streptococcus-related Stillbirth: A Systematic Review. BJOG Int. J. Obstet. Gynaecol. 2015, 122, 1437–1445. [Google Scholar] [CrossRef]
- Hall, J.; Adams, N.H.; Bartlett, L.; Seale, A.C.; Lamagni, T.; Bianchi-Jassir, F.; Lawn, J.E.; Baker, C.J.; Cutland, C.; Heath, P.T.; et al. Maternal Disease With Group B Streptococcus and Serotype Distribution Worldwide: Systematic Review and Meta-Analyses. Clin. Infect. Dis. 2017, 65, S112–S124. [Google Scholar] [CrossRef]
- Bizzarro, M.J.; Shabanova, V.; Baltimore, R.S.; Dembry, L.-M.; Ehrenkranz, R.A.; Gallagher, P.G. Neonatal Sepsis 2004-2013: The Rise and Fall of Coagulase-Negative Staphylococci. J. Pediatr. 2015, 166, 1193–1199. [Google Scholar] [CrossRef]
- Baker, C.J. The Spectrum of Perinatal Group B Streptococcal Disease. Vaccine 2013, 31 (Suppl. 4), D3–D6. [Google Scholar] [CrossRef]
- Patras, K.A.; Nizet, V. Group B Streptococcal Maternal Colonization and Neonatal Disease: Molecular Mechanisms and Preventative Approaches. Front. Pediatr. 2018, 6, 27. [Google Scholar] [CrossRef]
- Reingold, A.; Watt, J.P. Group B Streptococcus Infections of Soft Tissue and Bone in California Adults, 1995–2012. Epidemiol. Infect. 2015, 143, 3343–3350. [Google Scholar] [CrossRef]
- Muñoz, P.; Llancaqueo, A.; Rodríguez-Créixems, M.; Peláez, T.; Martin, L.; Bouza, E. Group B Streptococcus Bacteremia in Nonpregnant Adults. Arch. Intern. Med. 1997, 157, 213–216. [Google Scholar] [CrossRef]
- Watkins, L.K.F.; McGee, L.; Schrag, S.J.; Beall, B.; Jain, J.H.; Pondo, T.; Farley, M.M.; Harrison, L.H.; Zansky, S.M.; Baumbach, J.; et al. Epidemiology of Invasive Group B Streptococcal Infections Among Nonpregnant Adults in the United States, 2008. JAMA Intern. Med. 2019, 179, 479–488. [Google Scholar] [CrossRef]
- Phares, C.R. Epidemiology of Invasive Group B Streptococcal Disease in the United States, 1999. JAMA 2008, 299, 2056–2065. [Google Scholar] [CrossRef]
- Skoff, T.H.; Farley, M.M.; Petit, S.; Craig, A.S.; Schaffner, W.; Gershman, K.; Harrison, L.H.; Lynfield, R.; Boetani, J.M.; Zansky, S.; et al. Increasing Burden of Invasive Group B Streptococcal Disease in Nonpregnant Adults, 1990. Clin. Infect. Dis. 2009, 49, 85–92. [Google Scholar] [CrossRef]
- Schrag, S.J.; Zywicki, S.; Farley, M.M.; Reingold, A.L.; Harrison, L.H.; Lefkowitz, L.B.; Hadler, J.L.; Danila, R.; Cieslak, P.R.; Schuchat, A. Group B Streptococcal Disease in the Era of Intrapartum Antibiotic Prophylaxis. N. Engl. J. Med. 2000, 342, 15–20. [Google Scholar] [CrossRef]
- Kim, C.J.; Romero, R.; Chaemsaithong, P.; Chaiyasit, N.; Yoon, B.H.; Kim, Y.M. Acute Chorioamnionitis and Funisitis: Definition, Pathologic Features, and Clinical Significance. Am. J. Obstet. Gynecol. 2015, 213, S29–S52. [Google Scholar] [CrossRef] [PubMed]
- Soraisham, A.S.; Singhal, N.; McMillan, D.D.; Sauve, R.S.; Lee, S.K.; Network, C.N. A Multicenter Study on the Clinical Outcome of Chorioamnionitis in Preterm Infants. Am. J. Obstet. Gynecol. 2009, 200, 372.e1–372.e6. [Google Scholar] [CrossRef]
- Burke, C.; Chin, E.G. Chorioamnionitis at Term: Definition, Diagnosis, and Implications for Practice. J. Perinat. Neonatal Nurs. 2016, 30, 106–114. [Google Scholar] [CrossRef]
- Martinelli, P.; Sarno, L.; Maruotti, G.M.; Paludetto, R. Chorioamnionitis and Prematurity: A Critical Review. J. Matern.-Fetal Neonatal Med. 2012, 25, 21–23. [Google Scholar] [CrossRef]
- Pugni, L.; Pietrasanta, C.; Acaia, B.; Merlo, D.; Ronchi, A.; Ossola, M.W.; Bosari, S.; Mosca, F. Chorioamnionitis and Neonatal Outcome in Preterm Infants: A Clinical Overview. J. Matern.-Fetal Neonatal Med. 2016, 29, 1525–1529. [Google Scholar] [CrossRef]
- Vanderhoeven, J.P.; Bierle, C.J.; Kapur, R.P.; McAdams, R.M.; Beyer, R.P.; Bammler, T.K.; Farin, F.M.; Bansal, A.; Spencer, M.; Deng, M.; et al. Group B Streptococcal Infection of the Choriodecidua Induces Dysfunction of the Cytokeratin Network in Amniotic Epithelium: A Pathway to Membrane Weakening. PLoS Pathog. 2014, 10, e1003920. [Google Scholar] [CrossRef]
- Berardi, A.; Lugli, L.; Rossi, C.; Guidotti, I.; Lanari, M.; Creti, R.; Perrone, E.; Biasini, A.; Sandri, F.; Volta, A.; et al. Impact of Perinatal Practices for Early-Onset Group B Streptococcal Disease Prevention. Pediatr. Infect. Dis. J. 2013, 32, e265–e271. [Google Scholar] [CrossRef] [PubMed]
- Prevention of Group B Streptococcal Early-Onset Disease in Newborns: ACOG Committee Opinion, Number. Obstet. Gynecol. 2019, 134, e19–e40. [CrossRef]
- Ander, S.E.; Diamond, M.S.; Coyne, C.B. Immune Responses at the Maternal-Fetal Interface. Sci. Immunol. 2019, 4, aat6114. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Nowicki, B.; Yallampalli, C. Differential Expression of Uterine NO in Pregnant and Nonpregnant Rats with Intrauterine Bacterial Infection. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2001, 280, R1356–R1363. [Google Scholar] [CrossRef]
- Myatt, L.; Eis, A.L.; Brockman, D.E.; Greer, I.A.; Lyall, F. Endothelial Nitric Oxide Synthase in Placental Villous Tissue from Normal, Pre-Eclamptic and Intrauterine Growth Restricted Pregnancies. Hum. Reprod. 1997, 12, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Ghabour, M.S.; Eis, A.L.W.; Brockman, D.E.; Pollock, J.S.; Myatt, L. Immunohistochemical Characterization of Placental Nitric Oxide Synthase Expression in Preeclampsia. Am. J. Obstet. Gynecol. 1995, 173, 687–694. [Google Scholar] [CrossRef]
- Thomson, A.J.; Telfer, J.F.; Kohnen, G.; Young, A.; Cameron, I.T.; Greer, I.A.; Norman, J.E. Nitric Oxide Synthase Activity and Localization Do Not Change in Uterus and Placenta during Human Parturition. Hum. Reprod. 1997, 12, 2546–2552. [Google Scholar] [CrossRef]
- Tsai, W.C.; Strieter, R.M.; Zisman, D.A.; Wilkowski, J.M.; Bucknell, K.A.; Chen, G.H.; Standiford, T.J. Nitric Oxide Is Required for Effective Innate Immunity against Klebsiella Pneumoniae. Infect. Immun. 1997, 65, 1870–1875. [Google Scholar] [CrossRef] [PubMed]
- Chakravortty, D.; Hensel, M. Inducible Nitric Oxide Synthase and Control of Intracellular Bacterial Pathogens. Microbes Infect. 2003, 5, 621–627. [Google Scholar] [CrossRef]
- Nakatsuka, M.; Asagiri, K.; Kimura, Y.; Kamada, Y.; Tada, K.; Kudo, T. Generation of Peroxynitrite and Apoptosis in Placenta of Patients with Chorioamnionitis: Possible Implications in Placental Abruption. Hum. Reprod. 1999, 14, 1101–1106. [Google Scholar] [CrossRef]
- Dramsi, S.; Caliot, E.; Bonne, I.; Guadagnini, S.; Prévost, M.; Kojadinovic, M.; Lalioui, L.; Poyart, C.; Trieu-Cuot, P. Assembly and Role of Pili in Group B Streptococci. Mol. Microbiol. 2006, 60, 1401–1413. [Google Scholar] [CrossRef]
- Santi, I.; Scarselli, M.; Mariani, M.; Pezzicoli, A.; Masignani, V.; Taddei, A.; Grandi, G.; Telford, J.L.; Soriani, M. BibA: A Novel Immunogenic Bacterial Adhesin Contributing to Group B Streptococcus Survival in Human Blood. Mol. Microbiol. 2007, 63, 754–767. [Google Scholar] [CrossRef]
- Buscetta, M.; Papasergi, S.; Firon, A.; Pietrocola, G.; Biondo, C.; Mancuso, G.; Midiri, A.; Romeo, L.; Teti, G.; Speziale, P.; et al. FbsC, a Novel Fibrinogen-Binding Protein, Promotes Streptococcus Agalactiae-Host Cell Interactions. J. Biol. Chem. 2014, 289, 21003–21015. [Google Scholar] [CrossRef] [PubMed]
- Lalioui, L.; Pellegrini, E.; Dramsi, S.; Baptista, M.; Bourgeois, N.; Doucet-Populaire, F.; Rusniok, C.; Zouine, M.; Glaser, P.; Kunst, F.; et al. The SrtA Sortase of Streptococcus Agalactiae Is Required for Cell Wall Anchoring of Proteins Containing the LPXTG Motif, for Adhesion to Epithelial Cells, and for Colonization of the Mouse Intestine. Infect. Immun. 2005, 73, 3342–3350. [Google Scholar] [CrossRef]
- Brokaw, A.; Furuta, A.; Dacanay, M.; Rajagopal, L.; Waldorf, K.M.A. Bacterial and Host Determinants of Group B Streptococcal Vaginal Colonization and Ascending Infection in Pregnancy. Front. Cell Infect. Microbiol. 2021, 11, 720789. [Google Scholar] [CrossRef]
- Lindahl, G.; Stålhammar-Carlemalm, M.; Areschoug, T. Surface Proteins of Streptococcus Agalactiae and Related Proteins in Other Bacterial Pathogens. Clin. Microbiol. Rev. 2005, 18, 102–127. [Google Scholar] [CrossRef]
- Vornhagen, J.; Quach, P.; Boldenow, E.; Merillat, S.; Whidbey, C.; Ngo, L.Y.; Waldorf, K.M.A.; Rajagopal, L. Bacterial Hyaluronidase Promotes Ascending GBS Infection and Preterm Birth. mBio 2016, 7, e00781-16. [Google Scholar] [CrossRef] [PubMed]
- Vornhagen, J.; Armistead, B.; Santana-Ufret, V.; Gendrin, C.; Merillat, S.; Coleman, M.; Quach, P.; Boldenow, E.; Alishetti, V.; Leonhard-Melief, C.; et al. Group B Streptococcus Exploits Vaginal Epithelial Exfoliation for Ascending Infection. J. Clin. Investig. 2018, 128, 1985–1999. [Google Scholar] [CrossRef] [PubMed]
- Whidbey, C.; Harrell, M.I.; Burnside, K.; Ngo, L.; Becraft, A.K.; Iyer, L.M.; Aravind, L.; Hitti, J.; Waldorf, K.M.A.; Rajagopal, L. A Hemolytic Pigment of Group B Streptococcus Allows Bacterial Penetration of Human Placenta. J. Exp. Med. 2013, 210, 1265–1281. [Google Scholar] [CrossRef]
- Nizet, V.; Gibson, R.L.; Chi, E.Y.; Framson, P.E.; Hulse, M.; Rubens, C.E. Group B Streptococcal Beta-Hemolysin Expression Is Associated with Injury of Lung Epithelial Cells. Infect. Immun. 1996, 64, 3818–3826. [Google Scholar] [CrossRef]
- Doran, K.S.; Liu, G.Y.; Nizet, V. Group B Streptococcal Beta-Hemolysin/Cytolysin Activates Neutrophil Signaling Pathways in Brain Endothelium and Contributes to Development of Meningitis. J. Clin. Investig. 2003, 112, 736–744. [Google Scholar] [CrossRef]
- Doran, K.S.; Chang, J.C.W.; Benoit, V.M.; Eckmann, L.; Nizet, V. Group B Streptococcal Beta-Hemolysin/Cytolysin Promotes Invasion of Human Lung Epithelial Cells and the Release of Interleukin. J. Infect. Dis. 2002, 185, 196–203. [Google Scholar] [CrossRef]
- Randis, T.M.; Gelber, S.E.; Hooven, T.A.; Abellar, R.G.; Akabas, L.H.; Lewis, E.L.; Walker, L.B.; Byland, L.M.; Nizet, V.; Ratner, A.J. Group B Streptococcus β-Hemolysin/Cytolysin Breaches Maternal-Fetal Barriers to Cause Preterm Birth and Intrauterine Fetal Demise in Vivo. J. Infect. Dis. 2014, 210, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Y.; Doran, K.S.; Lawrence, T.; Turkson, N.; Puliti, M.; Tissi, L.; Nizet, V. Sword and Shield: Linked Group B Streptococcal Beta-Hemolysin/Cytolysin and Carotenoid Pigment Function to Subvert Host Phagocyte Defense. Proc. Natl. Acad. Sci. USA 2004, 101, 14491–14496. [Google Scholar] [CrossRef] [PubMed]
- Hensler, M.E.; Liu, G.Y.; Sobczak, S.; Benirschke, K.; Nizet, V.; Heldt, G.P. Virulence Role of Group B Streptococcus β-Hemolysin/Cytolysin in a Neonatal Rabbit Model of Early-Onset Pulmonary Infection. J. Infect. Dis. 2005, 191, 1287–1291. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, H.W. Nontypable Group B Streptococci Isolated from Human Sources. J. Clin. Microbiol. 1977, 6, 183–184. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Active Bacterial Core Surveillance (ABCs) Report; Emerging Infections Program Network Group B Streptococcus: Atlanta, GA, USA, 2016; Volume 2016. [Google Scholar]
- Pritzlaff, C.A.; Chang, J.C.; Kuo, S.P.; Tamura, G.S.; Rubens, C.E.; Nizet, V. Genetic Basis for the Β-haemolytic/Cytolytic Activity of Group B Streptococcus. Mol. Microbiol. 2001, 39, 236–248. [Google Scholar] [CrossRef]
- Laubach, V.E.; Shesely, E.G.; Smithies, O.; Sherman, P.A. Mice Lacking Inducible Nitric Oxide Synthase Are Not Resistant to Lipopolysaccharide-Induced Death. Proc. Natl. Acad. Sci. USA 1995, 92, 10688–10692. [Google Scholar] [CrossRef]
- Drewlo, S.; Levytska, K.; Kingdom, J. Revisiting the Housekeeping Genes of Human Placental Development and Insufficiency Syndromes. Placenta 2012, 33, 952–954. [Google Scholar] [CrossRef]
- Hrabie, J.A.; Keefer, L.K. Chemistry of the Nitric Oxide-Releasing Diazeniumdiolate (“Nitrosohydroxylamine”) Functional Group and Its Oxygen-Substituted Derivatives. Chem. Rev. 2002, 102, 1135–1154. [Google Scholar] [CrossRef]
- Grosser, M.R.; Paluscio, E.; Thurlow, L.R.; Dillon, M.M.; Cooper, V.S.; Kawula, T.H.; Richardson, A.R. Genetic Requirements for Staphylococcus Aureus Nitric Oxide Resistance and Virulence. PLoS Pathog. 2018, 14, e1006907. [Google Scholar] [CrossRef]
- Hooven, T.A.; Catomeris, A.J.; Bonakdar, M.; Tallon, L.J.; Santana-Cruz, I.; Ott, S.; Daugherty, S.C.; Tettelin, H.; Ratner, A.J. The Streptococcus Agalactiae Stringent Response Enhances Virulence and Persistence in Human Blood. Infect. Immun. 2017, 86, e00612-17. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Baerends, R.J.S.; Smits, W.K.; de Jong, A.; Hamoen, L.W.; Kok, J.; Kuipers, O.P. Genome2D: A Visualization Tool for the Rapid Analysis of Bacterial Transcriptome Data. Genome Biol. 2004, 5, R37. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, I. MBGD: Microbial Genome Database for Comparative Analysis. Nucleic Acids Res. 2003, 31, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, I.; Mihara, M.; Nishide, H.; Chiba, H.; Kato, M. MBGD Update 2018: Microbial Genome Database Based on Hierarchical Orthology Relations Covering Closely Related and Distantly Related Comparisons. Nucleic Acids Res. 2018, 47, D382–D389. [Google Scholar] [CrossRef]
- Tranguch, S.; Huet-Hudson, Y. Decreased Viability of Nitric Oxide Synthase Double Knockout Mice. Mol. Reprod. Dev. 2003, 65, 175–179. [Google Scholar] [CrossRef]
- Sagar, A.; Klemm, C.; Hartjes, L.; Mauerer, S.; van Zandbergen, G.; Spellerberg, B. The β-Hemolysin and Intracellular Survival of Streptococcus Agalactiae in Human Macrophages. PLoS ONE 2013, 8, e60160. [Google Scholar] [CrossRef]
- Nizet, V.; Kim, K.S.; Stins, M.; Jonas, M.; Chi, E.Y.; Nguyen, D.; Rubens, C.E. Invasion of Brain Microvascular Endothelial Cells by Group B Streptococci. Infect. Immun. 1997, 65, 5074–5081. [Google Scholar] [CrossRef]
- Bebien, M.; Hensler, M.E.; Davanture, S.; Hsu, L.-C.; Karin, M.; Park, J.M.; Alexopoulou, L.; Liu, G.Y.; Nizet, V.; Lawrence, T. The Pore-Forming Toxin β Hemolysin/Cytolysin Triggers P38 MAPK-Dependent IL-10 Production in Macrophages and Inhibits Innate Immunity. PLoS Pathog. 2012, 8, e1002812. [Google Scholar] [CrossRef]
- Whidbey, C.; Vornhagen, J.; Gendrin, C.; Samson, J.M.; Doering, K.; Ngo, L.; Gundlach, J.; Elovitz, M.; Liggitt, D.; Duncan, J.; et al. A Bacterial Lipid Toxin Induces Pore Formation, Pyroptosis, and Infection-Associated Fetal Injury and Preterm Birth. PLoS Pathog. 2014, 1, 41. [Google Scholar]
- Whidbey, C.; Vornhagen, J.; Gendrin, C.; Boldenow, E.; Samson, J.M.; Doering, K.; Ngo, L.; Ezekwe, E.A.D.; Gundlach, J.H.; Elovitz, M.A.; et al. A Streptococcal Lipid Toxin Induces Membrane Permeabilization and Pyroptosis Leading to Fetal Injury. EMBO Mol. Med. 2015, 7, 488–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa-Fraile, M.; Rodríguez-Granger, J.; Haidour-Benamin, A.; Cuerva, J.M.; Sampedro, A. Granadaene: Proposed Structure of the Group B Streptococcus Polyenic Pigment. Appl. Environ. Microbiol. 2006, 72, 6367–6370. [Google Scholar] [CrossRef] [PubMed]
- Armistead, B.; Herrero-Foncubierta, P.; Coleman, M.; Quach, P.; Whidbey, C.; Justicia, J.; Tapia, R.; Casares, R.; Millán, A.; Haidour, A.; et al. Lipid Analogs Reveal Features Critical for Hemolysis and Diminish Granadaene Mediated Group B Streptococcus Infection. Nat. Commun. 2020, 11, 1502. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Beres, S.B.; Yerramilli, P.; Pruitt, L.; Cantu, C.C.; Olsen, R.J.; Musser, J.M. Genetic Basis Underlying the Hyperhemolytic Phenotype of Streptococcus Agalactiae Strain CNCTC10/84. J. Bacteriol. 2020, 202, e00504-20. [Google Scholar] [CrossRef]
- Madrid, L.; Seale, A.C.; Kohli-Lynch, M.; Edmond, K.M.; Lawn, J.E.; Heath, P.T.; Madhi, S.A.; Baker, C.J.; Bartlett, L.; Cutland, C.; et al. Infant Group B Streptococcal Disease Incidence and Serotypes Worldwide: Systematic Review and Meta-Analyses. Clin. Infect. Dis. 2017, 65, S160–S172. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-Scale Gene Function Analysis with the PANTHER Classification System. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef]
- Seale, A.C.; Bianchi-Jassir, F.; Russell, N.J.; Kohli-Lynch, M.; Tann, C.J.; Hall, J.; Madrid, L.; Blencowe, H.; Cousens, S.; Baker, C.J.; et al. Estimates of the Burden of Group B Streptococcal Disease Worldwide for Pregnant Women, Stillbirths, and Children. Clin. Infect. Dis. 2017, 65, S200–S219. [Google Scholar] [CrossRef]
- Krause, B.J.; Hanson, M.A.; Casanello, P. Role of Nitric Oxide in Placental Vascular Development and Function. Placenta 2011, 32, 797–805. [Google Scholar] [CrossRef]
- Maltepe, E.; Bakardjiev, A.I.; Fisher, S.J. The Placenta: Transcriptional, Epigenetic, and Physiological Integration during Development. J. Clin. Investig. 2010, 120, 1016–1025. [Google Scholar] [CrossRef]
- Coan, P.M.; Conroy, N.; Burton, G.J.; Ferguson-Smith, A.C. Origin and Characteristics of Glycogen Cells in the Developing Murine Placenta. Dev. Dynam. 2006, 235, 3280–3294. [Google Scholar] [CrossRef]
- Croy, A.; Yamada, A.T.; DeMayo, F.J.; Adamson, S.L. The Guide to Investigation of Mouse Pregnancy; Elsevier Science: Amsterdam, The Netherlands, 2014; ISBN 9780123947949. [Google Scholar]
- Megli, C.J.; Coyne, C.B. Infections at the Maternal–Fetal Interface: An Overview of Pathogenesis and Defence. Nat. Rev. Microbiol. 2022, 20, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Doster, R.; Rogers, L.; Aronoff, D.; Gaddy, J. Macrophages Produce Extracellular Traps in Response to Streptococcus Agalactiae Infection. Open Forum. Infect. Dis. 2015, 2, 867. [Google Scholar] [CrossRef]
- Houser, B.L. Decidual Macrophages and Their Roles at the Maternal-Fetal Interface. Yale J. Biol. Med. 2012, 85, 105–118. [Google Scholar]
- Thomas, J.R.; Appios, A.; Zhao, X.; Dutkiewicz, R.; Donde, M.; Lee, C.Y.C.; Naidu, P.; Lee, C.; Cerveira, J.; Liu, B.; et al. Phenotypic and Functional Characterization of First-Trimester Human Placental Macrophages, Hofbauer Cells. J. Exp. Med. 2020, 218, e20200891. [Google Scholar] [CrossRef]
- Ning, F.; Liu, H.; Lash, G.E. The Role of Decidual Macrophages during Normal and Pathological Pregnancy. Am. J. Reprod. Immunol. 2016, 75, 298–309. [Google Scholar] [CrossRef]
- Houser, B.L.; Tilburgs, T.; Hill, J.; Nicotra, M.L.; Strominger, J.L. Two Unique Human Decidual Macrophage Populations. J. Immunol. 2011, 186, 2633–2642. [Google Scholar] [CrossRef]
- Czikk, M.J.; McCarthy, F.P.; Murphy, K.E. Chorioamnionitis: From Pathogenesis to Treatment. Clin. Microbiol. Infect. 2011, 17, 1304–1311. [Google Scholar] [CrossRef]
- Romero, R.; Gomez-Lopez, N.; Winters, A.D.; Jung, E.; Shaman, M.; Bieda, J.; Panaitescu, B.; Pacora, P.; Erez, O.; Greenberg, J.M.; et al. Evidence That Intra-Amniotic Infections Are Often the Result of an Ascending Invasion—A Molecular Microbiological Study. J. Perinat. Med. 2019, 47, 915–931. [Google Scholar] [CrossRef]
- Lupo, A.; Ruppen, C.; Hemphill, A.; Spellerberg, B.; Sendi, P. Phenotypic and Molecular Characterization of Hyperpigmented Group B Streptococci. Int. J. Med. Microbiol. 2014, 304, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Hooven, T.A.; Randis, T.M.; Daugherty, S.C.; Narechania, A.; Planet, P.J.; Tettelin, H.; Ratner, A.J. Complete Genome Sequence of Streptococcus Agalactiae CNCTC 10/84, a Hypervirulent Sequence Type 26 Strain. Genome Announc. 2014, 2, e01338-14. [Google Scholar] [CrossRef]
- Pony, P.; Rapisarda, C.; Terradot, L.; Marza, E.; Fronzes, R. Filamentation of the Bacterial Bi-Functional Alcohol/Aldehyde Dehydrogenase AdhE Is Essential for Substrate Channeling and Enzymatic Regulation. Nat. Commun. 2020, 11, 1426. [Google Scholar] [CrossRef]
- Imlay, J.A. Where in the World Do Bacteria Experience Oxidative Stress? Environ. Microbiol. 2019, 21, 521–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedraz, L.; Blanco-Cabra, N.; Torrents, E. Gradual Adaptation of Facultative Anaerobic Pathogens to Microaerobic and Anaerobic Conditions. FASEB J. 2020, 34, 2912–2928. [Google Scholar] [CrossRef]
- Richardson, A.R.; Libby, S.J.; Fang, F.C. A Nitric Oxide–Inducible Lactate Dehydrogenase Enables Staphylococcus Aureus to Resist Innate Immunity. Science 2008, 319, 1672–1676. [Google Scholar] [CrossRef]
- Luong, T.T.; Kim, E.-H.; Bak, J.P.; Nguyen, C.T.; Choi, S.; Briles, D.E.; Pyo, S.; Rhee, D.-K. Ethanol-Induced Alcohol Dehydrogenase E (AdhE) Potentiates Pneumolysin in Streptococcus Pneumoniae. Infect. Immun. 2015, 83, 108–119. [Google Scholar] [CrossRef]
- Kim, H.; Bhunia, A.K. Secreted Listeria Adhesion Protein (Lap) Influences Lap-Mediated Listeria Monocytogenes Paracellular Translocation through Epithelial Barrier. Gut Pathog. 2013, 5, 16. [Google Scholar] [CrossRef]
- Lamy, M.-C.; Zouine, M.; Fert, J.; Vergassola, M.; Couve, E.; Pellegrini, E.; Glaser, P.; Kunst, F.; Msadek, T.; Trieu-Cuot, P.; et al. CovS/CovR of Group B Streptococcus: A Two-Component Global Regulatory System Involved in Virulence. Mol. Microbiol. 2004, 54, 1250–1268. [Google Scholar] [CrossRef]
- Poole, R.K. Flavohaemoglobin: The Pre-Eminent Nitric Oxide–Detoxifying Machine of Microorganisms. F1000Research 2020, 9, F1000 Faculty Rev-7. [Google Scholar] [CrossRef]
- Membrillo-Hernández, J.; Coopamah, M.D.; Anjum, M.F.; Stevanin, T.M.; Kelly, A.; Hughes, M.N.; Poole, R.K. The Flavohemoglobin of Escherichia Coli Confers Resistance to a Nitrosating Agent, a “Nitric Oxide Releaser,” and Paraquat and Is Essential for Transcriptional Responses to Oxidative Stress*. J. Biol. Chem. 1999, 274, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Ring, A.; Braun, J.S.; Nizet, V.; Stremmel, W.; Shenep, J.L. Group B Streptococcal β-Hemolysin Induces Nitric Oxide Production in Murine Macrophages. J. Infect. Dis. 2000, 182, 150–157. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, J.; Zheng, X.; Chen, L.; He, M.; Ma, S.; Lin, Y.; Lin, X.; Chen, C. Identification of Group B Streptococcus Serotypes and Genotypes in Late Pregnant Women and Neonates That Are Associated With Neonatal Early-Onset Infection in a South China Population. Front. Pediatr. 2020, 8, 265. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A909 (Serotype Ia) Gene Locus | 10/84 (Serotype V) Gene Locus | PANTHER Consensus Gene Annotation | A909 LFC | 10/84 LFC | Mean LFC |
|---|---|---|---|---|---|
| SAK_RS00425 | W903_RS00440 | alcohol dehydrogenase | 4.43 | 3.32 | 3.87 |
| SAK_RS00805 | W903_RS00840 | disulfide oxidoreductase | 3.56 | 2.84 | 3.20 |
| SAK_RS10680 | W903_RS10090 | hypothetical protein | 3.48 | 2.60 | 3.04 |
| SAK_RS03085 | W903_RS03140 | hypothetical protein | 3.58 | 2.28 | 2.93 |
| SAK_RS10170 | W903_RS09695 | pyruvate formate-lyase-activating enzyme | 2.75 | 3.02 | 2.89 |
| SAK_RS10175 | W903_RS09700 | hypothetical protein | 2.43 | 2.94 | 2.69 |
| SAK_RS10190 | W903_RS09715 | anaerobic ribonucleoside-triphosphate reductase | 2.31 | 2.88 | 2.60 |
| SAK_RS10585 | W903_RS09995 | hypothetical protein | 2.08 | 2.95 | 2.51 |
| SAK_RS04360 | W903_RS04040 | cytosine deaminase | 2.47 | 2.53 | 2.50 |
| SAK_RS10180 | W903_RS09705 | oxidoreductases | 2.28 | 2.65 | 2.46 |
| SAK_RS04365 | W903_RS04045 | riboflavin synthase alpha chain | 2.58 | 2.16 | 2.37 |
| SAK_RS00720 | W903_RS00750 | GRPE protein | 2.39 | 2.33 | 2.36 |
| SAK_RS10400 | W903_RS09920 | response regulator of two-component system | 2.36 | 2.21 | 2.29 |
| SAK_RS04370 | W903_RS04050 | GTP cyclohydrolase II-related | 2.53 | 2.03 | 2.28 |
| SAK_RS10275 | W903_RS09800 | hypothetical protein | 2.30 | 2.09 | 2.19 |
| SAK_RS10645 | W903_RS10055 | hypothetical protein | 2.08 | 2.27 | 2.18 |
| SAK_RS10230 | W903_RS09755 | hypothetical protein | 2.00 | 2.13 | 2.07 |
| A909 (Serotype Ia) Gene Locus | 10/84 (Serotype V) Gene Locus | PANTHER Consensus Gene Annotation | A909 LFC | 10/84 LFC | Mean LFC |
|---|---|---|---|---|---|
| SAK_RS04090 | W903_RS03770 | inner membrane protein YBBM-related iron transporter (fetB) | −3.19 | −4.23 | −3.71 |
| SAK_RS04085 | W903_RS03765 | hypothetical protein | −3.02 | −4.18 | −3.60 |
| SAK_RS07535 | W903_RS07235 | membrane component of amino acid ABC transporter | −3.22 | −3.47 | −3.34 |
| SAK_RS07540 | W903_RS07240 | hypothetical protein | −3.22 | −3.30 | −3.26 |
| SAK_RS06865 | W903_RS06570 | 5′-nucleotidase-related | −3.16 | −3.34 | −3.25 |
| SAK_RS04495 | W903_RS04200 | hypothetical protein | −2.87 | −2.25 | −2.56 |
| SAK_RS04500 | W903_RS04205 | histidine transport system permease protein (hisM) | −2.60 | −2.37 | −2.49 |
| SAK_RS06330 | W903_RS06145 | hypothetical protein | −2.38 | −2.27 | −2.33 |
| SAK_RS04505 | W903_RS04210 | D-methionine-binding lipoprotein (metQ) | −1.93 | −2.59 | −2.26 |
| SAK_RS07995 | W903_RS07675 | hypothetical protein | −2.26 | −2.20 | −2.23 |
| SAK_RS06205 | W903_RS06010 | s1 RNA-binding domain-containing protein 1 | −2.26 | −2.15 | −2.21 |
| SAK_RS06325 | W903_RS06140 | hypothetical protein | −2.11 | −2.22 | −2.16 |
| SAK_RS08985 | W903_RS08565 | glutamine synthetase | −2.00 | −2.04 | −2.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keith, M.F.; Gopalakrishna, K.P.; Bhavana, V.H.; Hillebrand, G.H.; Elder, J.L.; Megli, C.J.; Sadovsky, Y.; Hooven, T.A. Nitric Oxide Production and Effects in Group B Streptococcus Chorioamnionitis. Pathogens 2022, 11, 1115. https://doi.org/10.3390/pathogens11101115
Keith MF, Gopalakrishna KP, Bhavana VH, Hillebrand GH, Elder JL, Megli CJ, Sadovsky Y, Hooven TA. Nitric Oxide Production and Effects in Group B Streptococcus Chorioamnionitis. Pathogens. 2022; 11(10):1115. https://doi.org/10.3390/pathogens11101115
Chicago/Turabian StyleKeith, Mary Frances, Kathyayini Parlakoti Gopalakrishna, Venkata Hemanjani Bhavana, Gideon Hayden Hillebrand, Jordan Lynn Elder, Christina Joann Megli, Yoel Sadovsky, and Thomas Alexander Hooven. 2022. "Nitric Oxide Production and Effects in Group B Streptococcus Chorioamnionitis" Pathogens 11, no. 10: 1115. https://doi.org/10.3390/pathogens11101115