
Diagnostic Measures of Disease Progression in Cattle Following Natural Infection with Bovine Leukemia Virus

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. ELISA
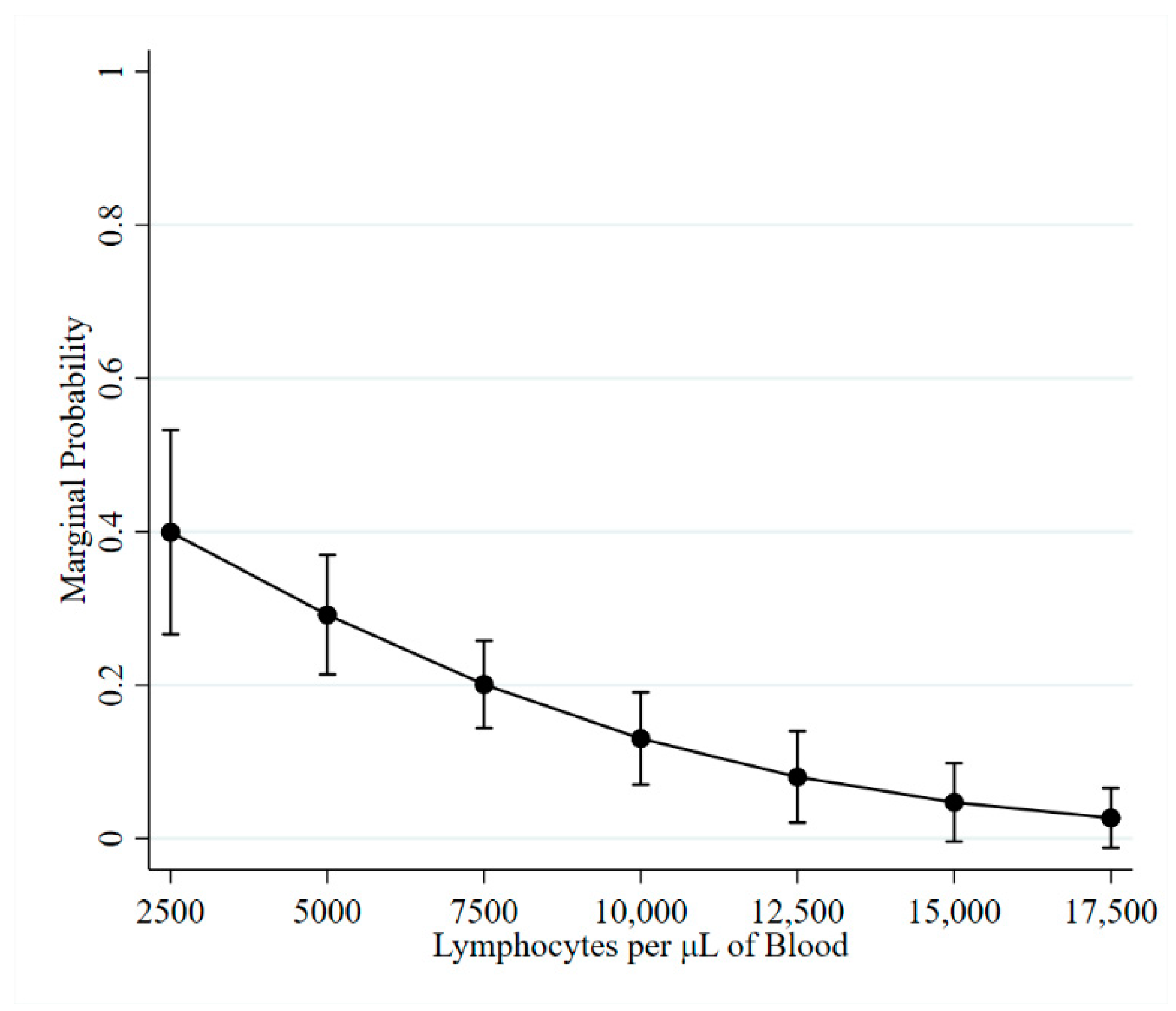
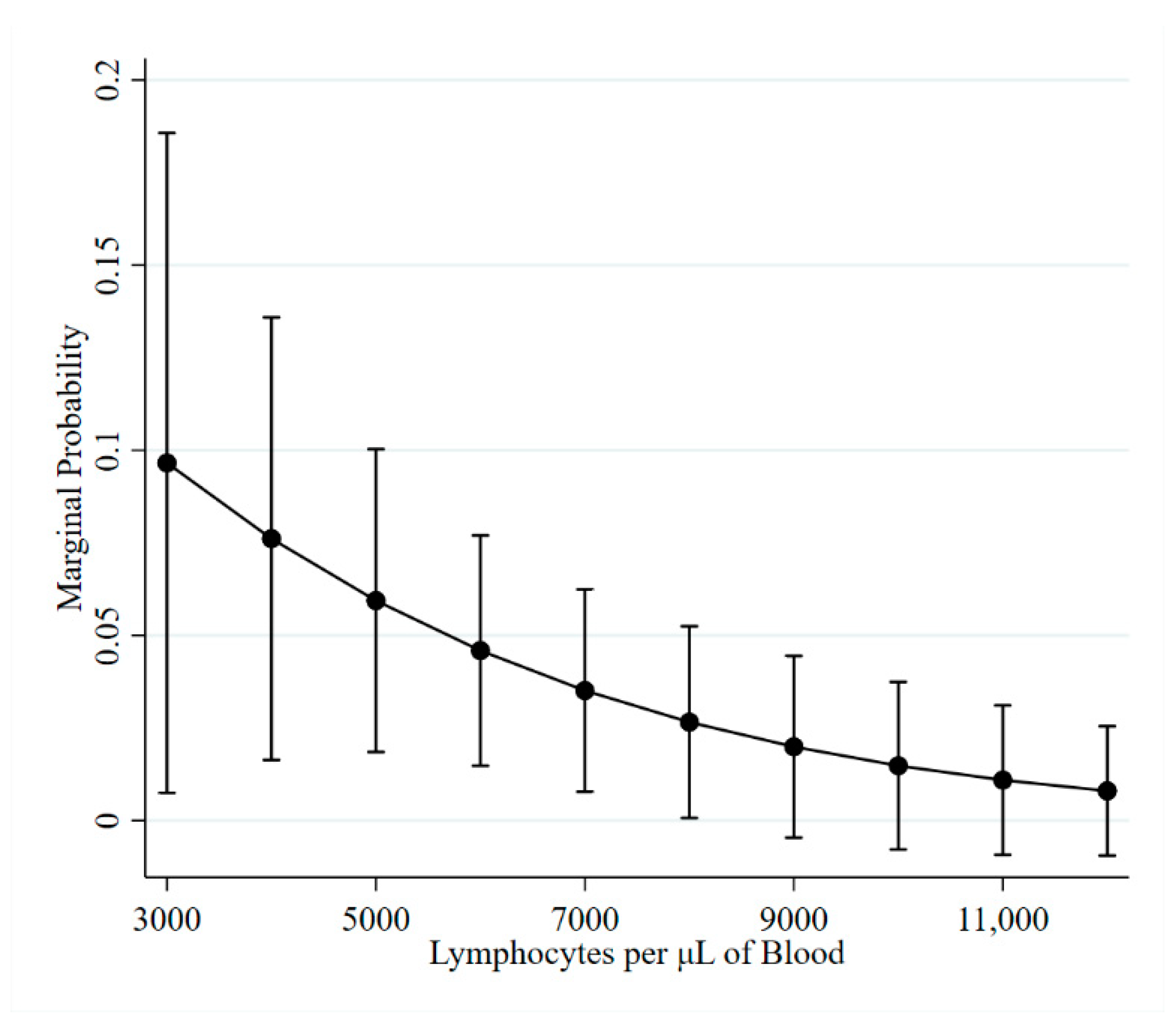
2.2. Lymphocyte Counts
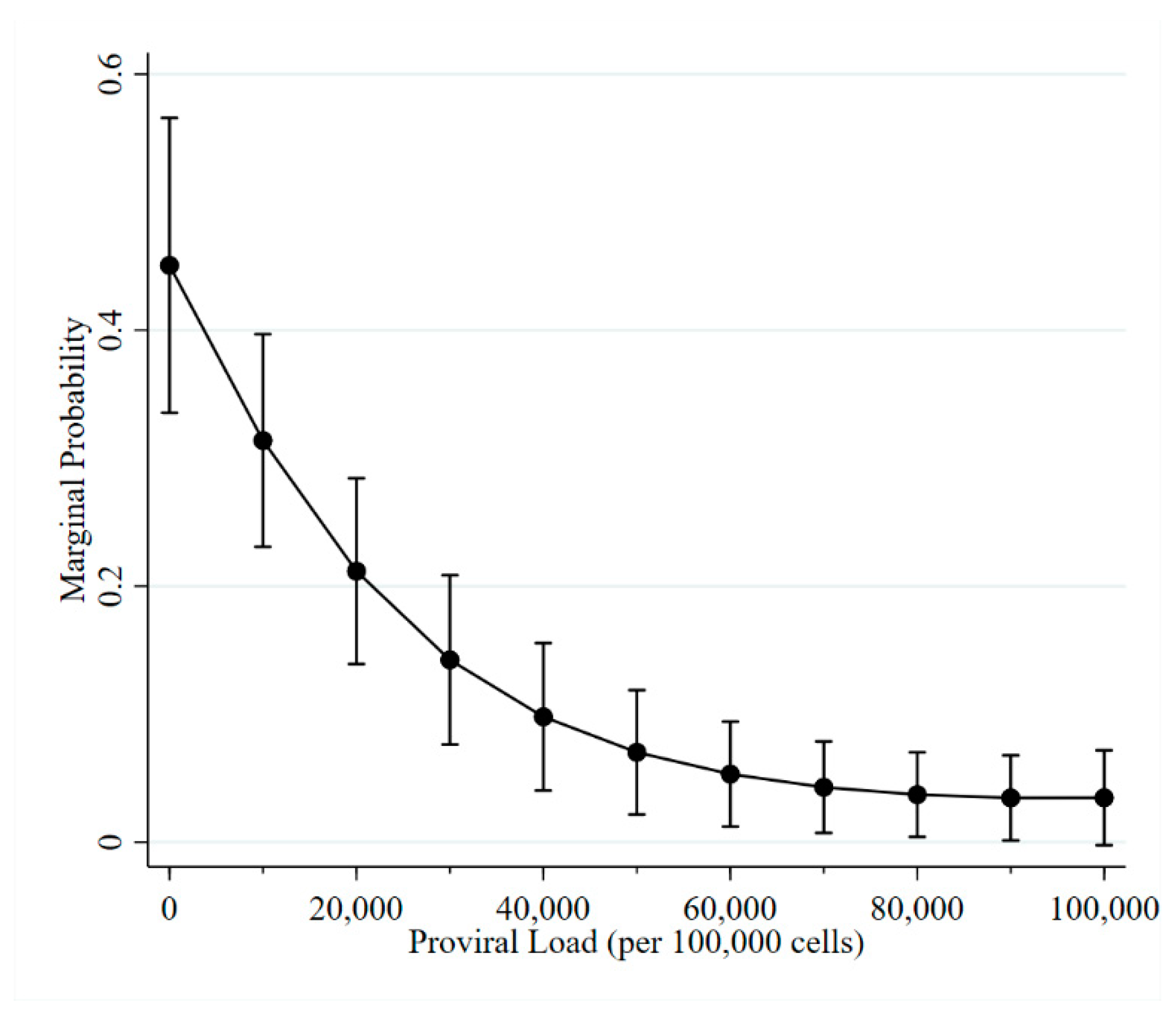
2.3. Proviral Load
3. Discussion
4. Materials and Methods
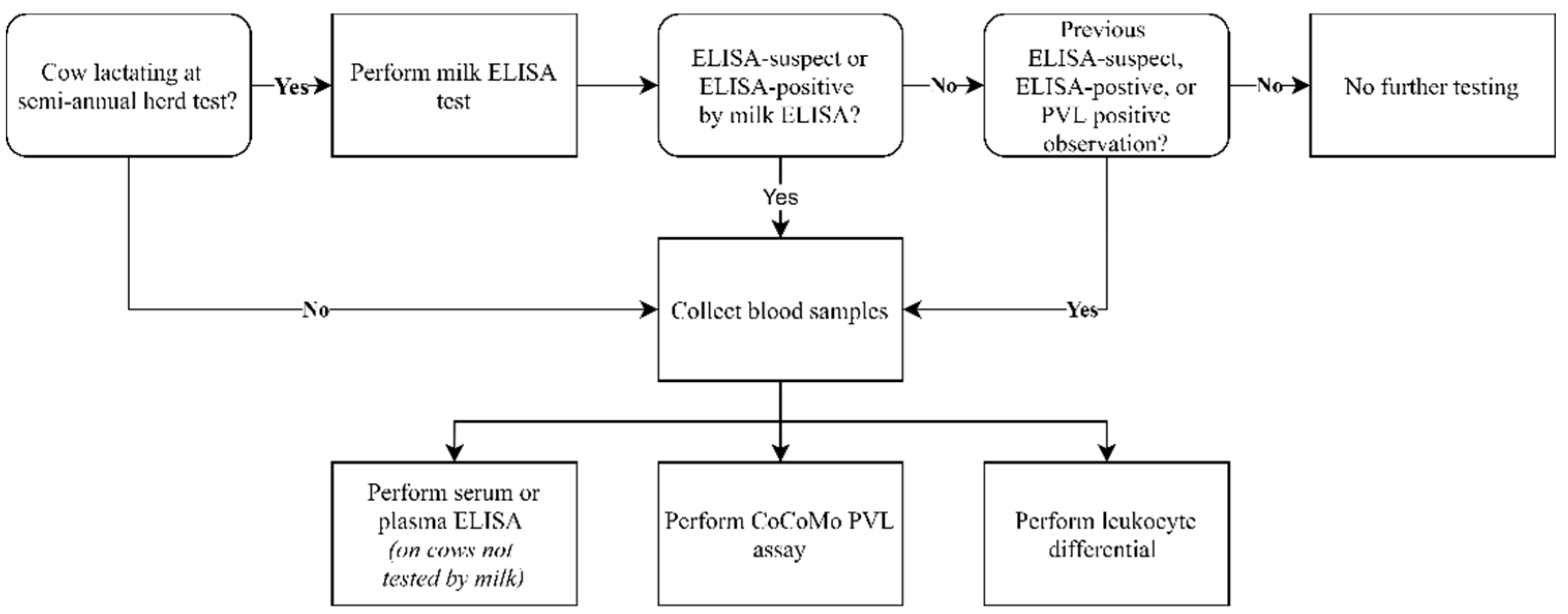
4.1. Study Design
4.2. Sample Analysis
4.3. Data Analysis
4.3.1. Descriptive Analysis
4.3.2. Analytical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Buehring, G.C.; Kramme, P.M.; Schultz, R.D. Evidence for bovine leukemia virus in mammary epithelial cells of infected cows. Lab. Investig. 1994, 71, 359–365. [Google Scholar] [CrossRef]
- Panei, C.J.; Takeshima, S.; Omori, T.; Nunoya, T.; Davis, W.C.; Ishizaki, H.; Matoba, K.; Aida, Y. Estimation of bovine leukemia virus (BLV) proviral load harbored by lymphocyte subpopulations in BLV-infected cattle at the subclinical stage of enzootic bovine leucosis using BLV-CoCoMo-qPCR. BMC Vet. Res. 2013, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, H.; Xie, B.; Oyamada, T.; Hiraga, A.; Yoshikawa, T. Detection of bovine leukemia viruses (BLV) in mammary tissues of BLV antibody-positive cows affected by subclinical mastitis. J. Vet. Med. Sci. 1997, 59, 301–302. [Google Scholar] [CrossRef] [Green Version]
- Bauermann, F.V.; Ridpath, J.F.; Dargatz, D.A. Bovine leukemia virus seroprevalence among cattle presented for slaughter in the United States. J. Vet. Diagn. Investig. 2017, 29, 704–706. [Google Scholar] [CrossRef] [Green Version]
- Ladronka, R.M.; Ainsworth, S.; Wilkins, M.J.; Norby, B.; Byrem, T.M.; Bartlett, P.C. Prevalence of Bovine Leukemia Virus Antibodies in US Dairy Cattle. Vet. Med. Int. 2018, 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- USDA. Bovine Leukosis Virus (BLV) on U.S. Dairy Operations. Available online: https://www.aphis.usda.gov/animal_health/nahms/dairy/downloads/dairy07/Dairy07_is_BLV.pdf (accessed on 21 December 2018).
- USDA. High Prevalence of BLV in U.S. Dairy Herds. Available online: https://www.aphis.usda.gov/animal_health/nahms/dairy/downloads/dairy96/Dairy96_is_BLV.pdf (accessed on 31 August 2018).
- USDA APHIS. Bovine Leukosis Virus in US Beef Cattle. Available online: https://www.aphis.usda.gov/animal_health/nahms/beefcowcalf/downloads/beef97/Beef97_is_BLV.pdf (accessed on 22 December 2018).
- Acaite, J.; Tamosiunas, V.; Lukauskas, K.; Milius, J.; Pieskus, J. The eradication experience of enzootic bovine leukosis from Lithuania. Prev. Vet. Med. 2007, 82, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Nuotio, L.; Rusanen, H.; Sihvonen, L.; Neuvonen, E. Eradication of enzootic bovine leukosis from Finland. Prev. Vet. Med. 2003, 59, 43–49. [Google Scholar] [CrossRef]
- Ferrer, J.F.; Piper, C.E. Role of Colostrum and Milk in the Natural Transmission of the Bovine Leukemia Virus. Cancer Res. 1981, 41, 4906–4909. [Google Scholar]
- Lucas, M.H.; Roberts, D.H.; Banks, J. Shedding of Bovine Leukosis Virus in Nasal Secretions of Infected Animals. Vet. Rec. 1993, 132, 276–278. [Google Scholar] [CrossRef]
- Miller, J.M.; Van Der Maaten, M.J. Infectivity tests of secretions and excretions from cattle infected with bovine leukemia virus. J. Natl. Cancer Inst. 1979, 62, 425–428. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Kitamura-Muramatsu, Y.; Saito, S.; Ishizaki, H.; Nakano, M.; Haga, S.; Matoba, K.; Ohno, A.; Murakami, H.; Takeshima, S.; et al. Detection of the BLV provirus from nasal secretion and saliva samples using BLV-CoCoMo-qPCR-2: Comparison with blood samples from the same cattle. Virus Res. 2015, 210, 248–254. [Google Scholar] [CrossRef]
- Hopkins, S.G.; Digiacomo, R.F. Natural Transmission of Bovine Leukemia Virus in Dairy and Beef Cattle. Vet. Clin. N. Am. Food Anim. Pract. 1997, 13, 107–128. [Google Scholar] [CrossRef]
- Rodríguez, S.M.; Florins, A.; Gillet, N.; de Brogniez, A.; Sánchez-Alcaraz, M.T.; Boxus, M.; Boulanger, F.; Gutiérrez, G.; Trono, K.; Alvarez, I.; et al. Preventive and therapeutic strategies for bovine leukemia virus: Lessons for HTLV. Viruses 2011, 3, 1210–1248. [Google Scholar] [CrossRef]
- Ruggiero, V.J.; Bartlett, P.C. Single-use hypodermic needles and obstetric sleeves failed to reduce bovine leukemia virus transmission in three dairy herds. Bov. Pract. 2019, 53, 128–133. [Google Scholar]
- Sprecher, D.J.; Pelzer, K.D.; Lessard, P. Possible Effect of Altered Management-Practices on Seroprevalence of Bovine Leukemia Virus in Heifers of a Dairy Herd with History of High Prevalence of Infection. J. Am. Vet. Med. Assoc. 1991, 199, 584–588. [Google Scholar] [PubMed]
- Gutiérrez, G.; Alvarez, I.; Politzki, R.; Lomónaco, M.; Dus Santos, M.J.; Rondelli, F.; Fondevila, N.; Trono, K. Natural progression of Bovine Leukemia Virus infection in Argentinean dairy cattle. Vet. Microbiol. 2011, 151, 255–263. [Google Scholar] [CrossRef]
- Molloy, J.B.; Dimmock, C.K.; Eaves, F.W.; Bruyeres, A.G.; Cowley, J.A.; Ward, W.H. Control of bovine leukaemia virus transmission by selective culling of infected cattle on the basis of viral antigen expression in lymphocyte cultures. Vet. Microbiol. 1994, 39, 323–333. [Google Scholar] [CrossRef]
- Ruggiero, V.J.; Norby, B.; Benitez, O.J.; Hutchinson, H.; Sporer, K.R.B.; Droscha, C.; Swenson, C.L.; Bartlett, P.C. Controlling bovine leukemia virus in dairy herds by identifying and removing cows with the highest proviral load and lymphocyte counts. J. Dairy Sci. 2019, 102, 9165–9175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, I.; Lévy, D. Pathobiology of bovine leukemia virus. Vet. Res. BioMed Cent. 1994, 25, 521–536. [Google Scholar]
- EFSA AHAW Panel (EFSA Panel on Animal Health and Welfare). Scientific opinion on enzootic bovine leukosis. EFSA J. 2015, 13, 4188. [Google Scholar] [CrossRef] [Green Version]
- Juliarena, M.A.; Gutierrez, S.E.; Ceriani, C. Determination of proviral load in bovine leukemia virus-infected cattle with and without lymphocytosis. Am. J. Vet. Res. 2007, 68, 1220–1225. [Google Scholar] [CrossRef]
- Nieto Farias, M.V.; de Souza, F.N.; Lendez, P.A.; Martínez-Cuesta, L.; Santos, K.R.; Libera, A.M.M.P.D.; Ceriani, M.C.; Dolcini, G.L. Lymphocyte proliferation and apoptosis of lymphocyte subpopulations in bovine leukemia virus-infected dairy cows with high and low proviral load. Vet. Immunol. Immunopathol. 2018, 206, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Juliarena, M.A.; Barrios, C.N.; Carolina Ceriani, M.; Esteban, E.N. Hot topic: Bovine leukemia virus (BLV)-infected cows with low proviral load are not a source of infection for BLV-free cattle. J. Dairy Sci. 2016, 99, 4586–4589. [Google Scholar] [CrossRef] [Green Version]
- Mekata, H.; Yamamoto, M.; Hayashi, T.; Kirino, Y.; Sekiguchi, S.; Konnai, S.; Horii, Y.; Norimine, J. Cattle with a low bovine leukemia virus proviral load are rarely an infectious source. Jpn. J. Vet. Res. 2018, 66, 157–163. [Google Scholar] [CrossRef]
- Bendixen, H.J. Bovine enzootic leukosis. Adv. Vet. Sci. 1965, 10, 129–204. [Google Scholar] [PubMed]
- Alvarez, I.; Gutiérrez, G.; Gammella, M.; Martínez, C.; Politzki, R.; González, C.; Caviglia, L.; Carignano, H.; Fondevila, N.; Poli, M.; et al. Evaluation of total white blood cell count as a marker for proviral load of bovine leukemia virus in dairy cattle from herds with a high seroprevalence of antibodies against bovine leukemia virus. Am. J. Vet. Res. 2013, 74, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Ohno, A.; Takeshima, S.-N.; Matsumoto, Y.; Aida, Y. Risk factors associated with increased bovine leukemia virus proviral load in infected cattle in Japan from 2012 to 2014. Virus Res. 2015, 210, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Lendez, P.A.; Passucci, J.A.; Poli, M.A.; Gutierrez, S.E.; Dolcini, G.L.; Ceriani, M.C. Association of TNF-α gene promoter region polymorphisms in bovine leukemia virus (BLV)-infected cattle with different proviral loads. Arch. Virol. 2015, 160, 2001–2007. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, H.C.; Norby, B.; Droscha, C.J.; Sordillo, L.M.; Coussens, P.M.; Bartlett, P.C. Bovine leukemia virus detection and dynamics following experimental inoculation. Res. Vet. Sci. 2020, 133, 269–275. [Google Scholar] [CrossRef]
- Forletti, A.; Lützelschwab, C.M.; Cepeda, R.; Esteban, E.N.; Gutiérrez, S.E. Early events following bovine leukaemia virus infection in calves with different alleles of the major histocompatibility complex DRB3 gene. Vet. Res. 2020, 51, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Juliarena, M.A.; Poli, M.; Sala, L.; Ceriani, C.; Gutierrez, S.; Dolcini, G.; Rodríguez, E.M.; Mariño, B.; Rodríguez-Dubra, C.; Esteban, E.N. Association of BLV infection profiles with alleles of the BoLA-DRB3.2 gene. Anim. Genet. 2008, 39, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Gillet, N.A.; Gutiérrez, G.; Rodriguez, S.M.; de Brogniez, A.; Renotte, N.; Alvarez, I.; Trono, K.; Willems, L.; Rodríguez, S.M.; de Brogniez, A.; et al. Massive Depletion of Bovine Leukemia Virus Proviral Clones Located in Genomic Transciptionally Active Sites during Primary Infection. PLoS Pathog. 2013, 9, e1003687. [Google Scholar] [CrossRef] [PubMed]
- Jimba, M.; Takeshima, S.-N.; Murakami, H.; Kohara, J.; Kobayashi, N.; Matsuhashi, T.; Ohmori, T.; Nunoya, T.; Aida, Y. BLV-CoCoMo-qPCR: A useful tool for evaluating bovine leukemia virus infection status. BMC Vet. Res. 2012, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, G.; Alvarez, I.; Merlini, R.; Rondelli, F.; Trono, K. Dynamics of perinatal bovine leukemia virus infection. BMC Vet. Res. 2014, 10, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konishi, M.; Ishizaki, H.; Kameyama, K.; Murakami, K.; Yamamoto, T. The effectiveness of colostral antibodies for preventing bovine leukemia virus (BLV) infection in vitro. BMC Vet. Res. 2018, 14, 1–9. [Google Scholar] [CrossRef]
- Taxis, T.M.; Dejong, T.N.; Swenson, C.L.; Sporer, K.R.B.; Droscha, C.; Niles, D.; Bartlett, P.C. Reducing bovine leukemia virus prevalence on a large midwestern dairy farm by using lymphocyte counts, ELISA antibody testing, and proviral load. Bov. Pract. 2020, 54, 136–144. [Google Scholar]
- Nekouei, O.A.; Sanchez, J.; Keefe, G.P. Carryover of bovine leukemia virus antibodies in samples from shared milk meters. J. Dairy Sci. 2015, 98, 5274–5279. [Google Scholar] [CrossRef]
- Norby, B.; Bartlett, P.; Byrem, T.; Erskine, R. Effect of infection with bovine leukemia virus on milk production in Michigan dairy cows. J. Dairy Sci. 2016, 99, 2043–2052. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, P.C.; Norby, B.; Byrem, T.M.; Parmelee, A.; Ledergerber, J.T.; Erskine, R.J. Bovine leukemia virus and cow longevity in Michigan dairy herds. J. Dairy Sci. 2013, 96, 1591–1597. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, R.M.; Song, Z.; Poon, H.; Heeney, J.L.; Taylor, J.A.; Jefferson, B.; Vernau, W.; Valli, V.E. Proviral detection and serology in bovine leukemia virus-exposed normal cattle and cattle with lymphoma. Can. J. Vet. Res. 1992, 56, 339–348. [Google Scholar] [PubMed]
- Chen, Y.C.; Chang, C.C.; Hsu, W.L.; Chuang, S.T. Dairy cattle with bovine leukaemia virus RNA show significantly increased leukocyte counts. Vet. J. 2020, 257, 105449. [Google Scholar] [CrossRef]
- George, J.W.; Snipes, J.; Lane, V.M. Comparison of bovine hematology reference intervals from 1957 to 2006. Vet. Clin. Pathol. 2010, 39, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Florins, A.; Boxus, M.; Vandermeers, F.; Verlaeten, O.; Bouzar, A.B.; Defoiche, J.; Hubaux, R.; Burny, A.; Kettmann, R.; Willems, L. Emphasis on cell turnover in two hosts infected by bovine leukemia virus: A rationale for host susceptibility to disease. Vet. Immunol. Immunopathol. 2008, 125, 1–7. [Google Scholar] [CrossRef]
- Erskine, R.J.; Bartlett, P.C.; Byrem, T.M.; Render, C.L.; Febvay, C.; Houseman, J.T. Using a Herd Profile to Determine Age-Specific Prevalence of Bovine Leukemia Virus in Michigan Dairy Herds. Vet. Med. Int. 2012, 2012, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimba, M.; Takeshima, S.; Matoba, K.; Endoh, D.; Aida, Y. BLV-CoCoMo-qPCR: Quantitation of bovine leukemia virus proviral load using the CoCoMo algorithm. Retrovirology 2010, 7, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Number of Observations | ||||||
|---|---|---|---|---|---|---|
| 0 a | 1 | 2 | 3 | 4 | 5 | |
| Observations Overall | -- | 29.8% | 21.1% | 18.0% | 17.6% | 13.6% |
| (232/779) | (164/779) | (140/779) | (137/779) | (106/779) | ||
| ELISA Tests | 0.4% | 30.7% | 20.6% | 18.2% | 17.2% | 12.8% |
| (3/779) | (239/779) | (161/779) | (142/779) | (134/779) | (100/779) | |
| ELISA-SP b | 47.9% | 12.2% | 10.9% | 10.8% | 10.4% | 7.8% |
| (373/779) | (95/779) | (85/779) | (84/779) | (81/779) | (61/779) | |
| PVL Tests | 47.1% | 16.6% | 14.3% | 11.4% | 9.0% | 1.7% |
| (367/779) | (129/779) | (111/779) | (89/779) | (70/779) | (13/779) | |
| PVL-Positive | 51.2% | 16.8% | 13.0% | 10.4% | 7.1% | 1.5% |
| (399/779) | (131/779) | (101/779) | (81/779) | (55/779) | (12/779) | |
| Leukocyte Differentials | 52.5% | 16.2% | 11.3% | 11.2% | 8.9% | -- c |
| (409/779) | (126/779) | (88/779) | (87/779) | (69/779) | ||
| Combined ELISA, PVL, LC | 53.5% | 16.7% | 11.3% | 10.9% | 7.6% | -- c |
| (417/779) | (130/779) | (88/779) | (85/779) | (59/779) | ||
| ELISA New Infection |
|---|
| ELISA-suspect or ELISA-positive result |
| Concurrent or subsequent PVL-positive result(s) |
| Prior ELISA-negative result(s) |
| No Prior PVL-positive result(s) |
| ELISA False Negative |
| ELISA-negative result |
| Prior ELISA-suspect or ELISA-positive result Either concurrent, or prior and subsequent PVL-positive test(s) |
| ELISA False Positive |
| ELISA-suspect or ELISA-positive result |
| Concurrent PVL-negative result |
| Prior or subsequent ELISA-negative result(s) |
| Prior or subsequent PVL-negative result(s) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hutchinson, H.C.; Ruggiero, V.J.; Norby, B.; Sporer, K.R.B.; Bartlett, P.C. Diagnostic Measures of Disease Progression in Cattle Following Natural Infection with Bovine Leukemia Virus. Pathogens 2021, 10, 987. https://doi.org/10.3390/pathogens10080987
Hutchinson HC, Ruggiero VJ, Norby B, Sporer KRB, Bartlett PC. Diagnostic Measures of Disease Progression in Cattle Following Natural Infection with Bovine Leukemia Virus. Pathogens. 2021; 10(8):987. https://doi.org/10.3390/pathogens10080987
Chicago/Turabian StyleHutchinson, Holden C., Vickie J. Ruggiero, Bo Norby, Kelly R. B. Sporer, and Paul C. Bartlett. 2021. "Diagnostic Measures of Disease Progression in Cattle Following Natural Infection with Bovine Leukemia Virus" Pathogens 10, no. 8: 987. https://doi.org/10.3390/pathogens10080987