
Molecular Epidemiology of G6PD Genotypes in Different Ethnic Groups Residing in Saharan and Sahelian Zones of Mauritania

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Characteristics of Participants
2.2. Prevalence and Type of G6PD Genotype and Gender
2.3. Prevalence and Type of G6PD Genotype and Ethnic Groups
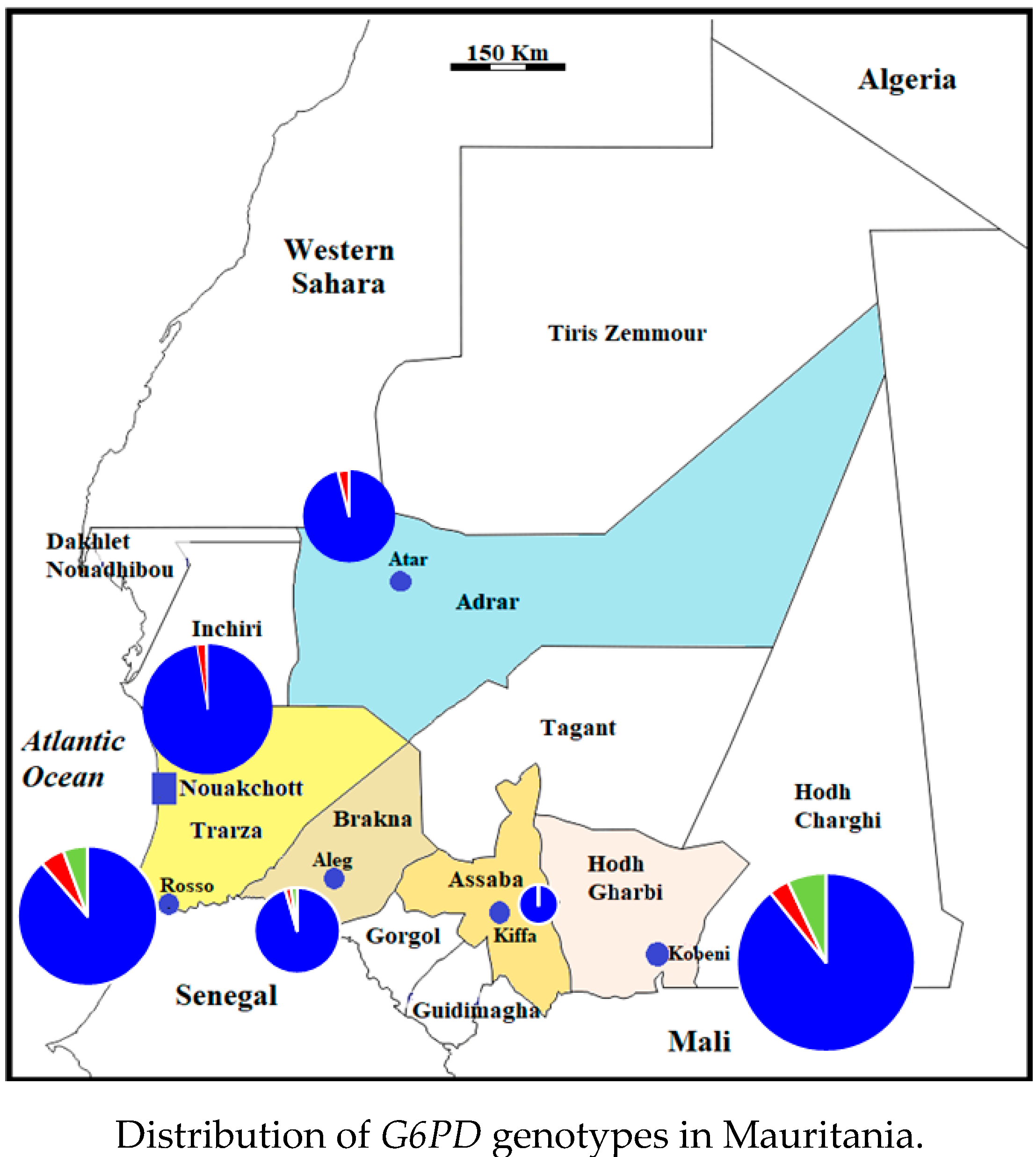
2.4. Distribution of G6PD Genotypes in Mauritania
3. Discussion
4. Materials and Methods
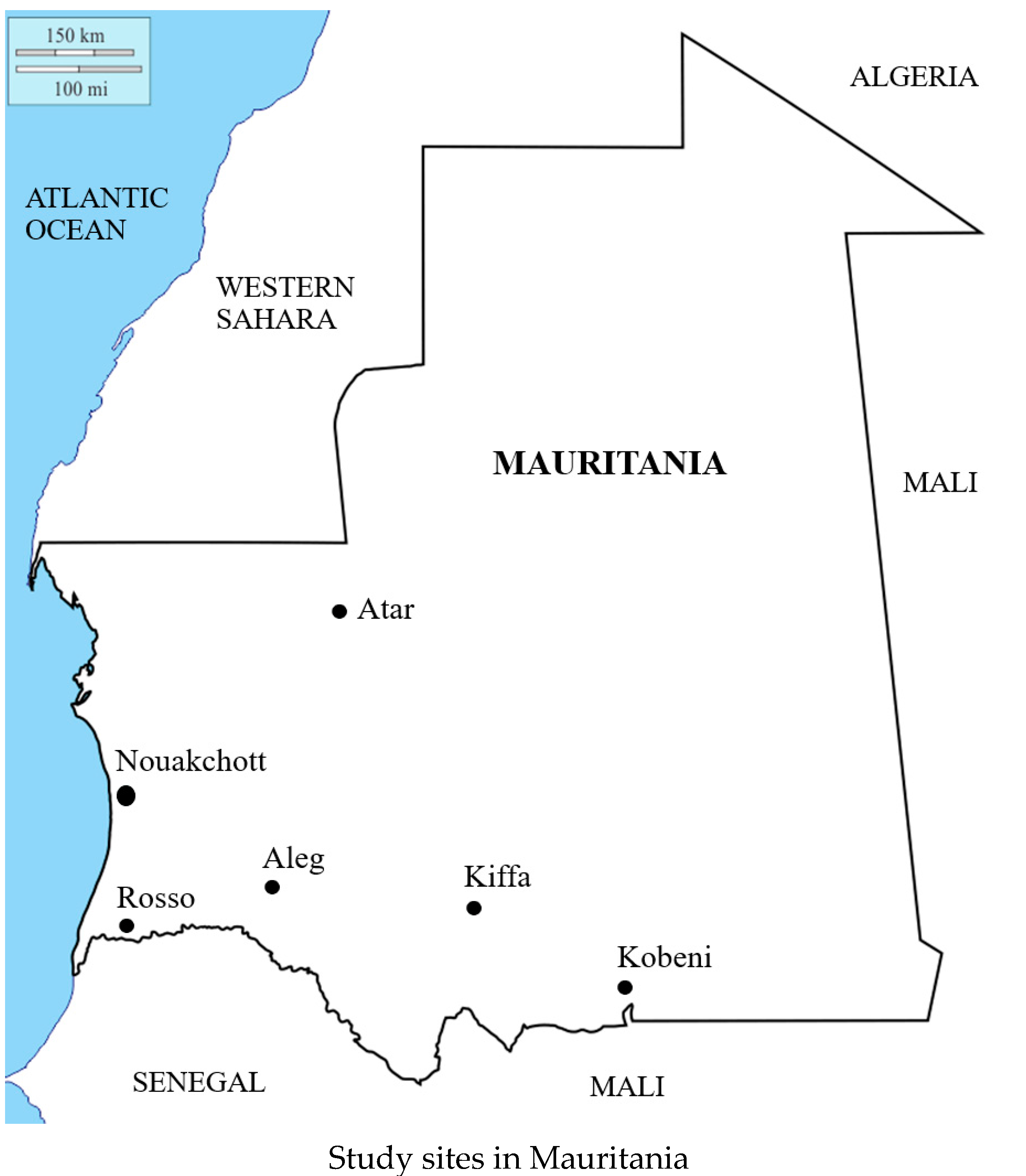
4.1. Study Sites
4.2. Collection of Blood Samples
4.3. Genotyping
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beutler, E. Glucose-6-phosphate dehydrogenase deficiency. Br. J. Haematol. 1970, 18, 117–121. [Google Scholar] [CrossRef]
- Vulliamy, T.; Mason, P.; Luzzatto, L. The molecular basis of glucose-6-phosphate dehydrogenase deficiency. Trends Genet. 1992, 8, 138–143. [Google Scholar] [CrossRef]
- Nkhoma, E.T.; Poole, C.; Vannappagari, V.; Hall, S.A.; Beutler, E. The global prevalence of glucose-6-phosphate dehydrogenase deficiency: A systematic review and meta-analysis. Blood Cells Mol. Dis. 2009, 42, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Howes, R.E.; Battle, K.E.; Satyagraha, A.W.; Baird, J.K.; Hay, S.I. G6PD deficiency: Global distribution, genetic variants and primaquine therapy. Adv. Parasitol. 2013, 81, 133–201. [Google Scholar] [CrossRef] [PubMed]
- Luzzatto, L.; Arese, P. Favism and glucose-6-phosphate dehydrogenase deficiency. N. Engl. J. Med. 2018, 378, 60–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippi, G.; Mattiuzzi, C. Updated worldwide epidemiology of inherited erythrocyte disorders. Acta Haematol. 2020, 143, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Persico, M.G.; Viglietto, G.; Martini, G.; Toniolo, D.; Paonessa, G.; Moscatelli, C.; Dono, R.; Vulliamy, T.; Luzzatto, L.; D’Urso, M. Isolation of human glucose-6-phosphate dehydrogenase (G6PD) cDNA clones: Primary structure of the protein and unusual 5′ non-coding region. Nucleic Acids Res. 1986, 14, 2511–2522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minucci, A.; Moradkhani, K.; Hwang, M.J.; Zuppi, C.; Giardina, B.; Capoluongo, E. Glucose-6-phosphate dehydrogenase (G6PD) mutations database: Review of the “old” and update of the new mutations. Blood Cells Mol. Dis. 2012, 48, 154–165. [Google Scholar] [CrossRef]
- Gómez-Manzo, S.; Marcial-Quino, J.; Vanoye-Carlo, A.; Serrano-Posada, H.; Ortega-Cuellar, D.; González-Valdez, A.; Castillo-Rodríguez, R.A.; Hernández-Ochoa, B.; Sierra-Palacios, E.; Rodríguez-Bustamante, E.; et al. Glucose-6-phosphate dehydrogenase: Update and analysis of new mutations around the world. Int. J. Mol. Sci. 2016, 17, 2069. [Google Scholar] [CrossRef] [Green Version]
- Collins, W.E.; Jeffery, G.M. Plasmodium ovale: Parasite and disease. Clin. Microbiol. Rev. 2005, 18, 570–581. [Google Scholar] [CrossRef] [Green Version]
- White, N.J.; Imwong, M. Relapse. Adv. Parasitol. 2012, 80, 113–150. [Google Scholar] [CrossRef]
- WHO. Guidelines for the Treatment of Malaria, 3rd ed.; World Health Organization: Geneva, Switzerland, 2015; pp. 1–313. [Google Scholar]
- Lekweiry, K.M.; Salem, M.S.; Basco, L.K.; Briolant, S.; Hafid, J.; Boukhary, A.O. Malaria in Mauritania: Retrospective and prospective overview. Malar. J. 2015, 14, 100. [Google Scholar] [CrossRef] [Green Version]
- Ould Ahmedou Salem, M.S.; Mint Lekweiry, K.; Mint Deida, J.; Ould Emouh, A.; Ould Weddady, M.; Ould Mohamed Salem Boukhary, A.; Basco, L.K. Increasing prevalence of Plasmodium vivax among febrile patients in Nouakchott, Mauritania. Am. J. Trop. Med. Hyg. 2015, 92, 537–540. [Google Scholar] [CrossRef] [Green Version]
- Mint Deida, J.; Ould Khalef, Y.; Mint Semane, E.; Ould Ahmedou Salem, M.S.; Bogreau, H.; Basco, L.; Ould Mohamed Salem Boukhary, A.; Tahar, R. Assessment of drug resistance associated genetic diversity in Mauritanian isolates of Plasmodium vivax reveals limited polymorphism. Malar. J. 2018, 17, 416. [Google Scholar] [CrossRef] [PubMed]
- Deida, J.; Tahar, R.; Khalef, Y.O.; Lekweiry, K.M.; Hmeyade, A.; Khairy, M.L.O.; Simard, F.; Bogreau, H.; Basco, L.; Boukhary, A.O.M.S. Oasis malaria, northern Mauritania. Emerg. Infect. Dis. 2019, 25, 273–280. [Google Scholar] [CrossRef]
- Mauritanian Ministry of Health. Guide Clinique et Thérapeutique à L’usage du Personnel des Centres de Santé de la Mauritanie, 2nd ed.; Agencia Española de Cooperación Internacional para el Desarrollo (AECID): Madrid, Spain, 2013; pp. 1–244. Available online: http://www.sante.gov.mr/?wpfb_dl=140 (accessed on 22 January 2021).
- Djigo, O.K.M.; Bollahi, M.A.; Hasni Ebou, M.; Ould Ahmedou Salem, M.S.; Tahar, R.; Bogreau, H.; Basco, L.; Ould Mohamed Salem Boukhary, A. Assessment of glucose-6-phosphate dehydrogenase activity using CareStart G6PD rapid diagnostic test and associated genetic variants in Plasmodium vivax malaria endemic setting in Mauritania. PLoS ONE 2019, 14, e0220977. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, G.S.; Lemine, S.M.; Cheibetta, S.; Mohamed, A. Neonatal screening for glucose-6-phosphate dehydrogenase (G6PD) deficiency in Mauritania. Pan Afr. Med. J. 2018, 30, 224. [Google Scholar] [CrossRef]
- WHO Working Group. Glucose-6-phosphate dehydrogenase deficiency. Bull. World Health Organ. 1989, 67, 601–611. [Google Scholar]
- Handloff, R.E. (Ed.) Mauritania: A Country Study, 2nd ed.; Federal Research Division; U.S. Government Printing Office for the Library of Congress: Washington, DC, USA, 1990; pp. 1–218.
- Nafa, K.; Reghis, A.; Osmani, N.; Baghli, L.; Aït-Abbes, H.; Benabadji, M.; Kaplan, J.C.; Vulliamy, T.; Luzzatto, L. At least five polymorphic mutants account for the prevalence of glucose-6-phosphate dehydrogenase deficiency in Algeria. Hum. Genet. 1994, 94, 513–517. [Google Scholar] [CrossRef] [PubMed]
- Doss, C.G.; Alasmar, D.R.; Bux, R.I.; Sneha, P.; Bakhsh, F.D.; Al-Azwani, I.; Bekay, R.E.; Zayed, H. Genetic epidemiology of glucose-6-phosphate dehydrogenase deficiency in the Arab world. Sci. Rep. 2016, 6, 37284. [Google Scholar] [CrossRef] [Green Version]
- Migot-Nabias, F.; Pelleau, S.; Watier, L.; Guitard, J.; Toly, C.; De Araujo, C.; Ngom, M.I.; Chevillard, C.; Gaye, O.; Garcia, A. Red blood cell polymorphisms in relation to Plasmodium falciparum asymptomatic parasite densities and morbidity in Senegal. Microbes Infect. 2006, 8, 2352–2358. [Google Scholar] [CrossRef] [PubMed]
- Carter, N.; Pamba, A.; Duparc, S.; Waitumbi, J.N. Frequency of glucose-6-phosphate dehydrogenase deficiency in malaria patients from six African countries enrolled in two randomized anti-malarial clinical trials. Malar. J. 2011, 10, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tine, R.C.K.; Ndiaye, M.; Hansson, H.H.; Ndour, C.T.; Faye, B.; Alifrangis, M.; Sylla, K.; Ndiaye, J.L.; Magnussen, P.; Bygbjerg, I.C.; et al. The association between malaria parasitaemia, erythrocyte polymorphisms, malnutrition and anaemia in children less than 10 years in Senegal: A case control study. BMC Res. Notes 2012, 5, 565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiga, B.; Dolo, A.; Campino, S.; Sepulveda, N.; Corran, P.; Rockett, K.A.; Troye-Blomberg, M.; Doumbo, O.K.; Clark, T.G. Glucose-6-phosphate dehydrogenase polymorphisms and susceptibility to mild malaria in Dogon and Fulani, Mali. Malar. J. 2014, 13, 270. [Google Scholar] [CrossRef] [Green Version]
- Benyachou, B. L’anémie par Déficit en G6PD Chez L’enfant. MD Thesis, Faculté de Médecine et de Pharmacie de Fès, Fès, Morocco,2008. Available online: http://scolarite.fmp-usmba.ac.ma/cdim/mediatheque/e_theses/4-08.pdf (accessed on 30 January 2021).
- Benabadji, M.; Merad, F.; Benmoussa, M.; Trabuchet, G.; Junien, C.; Dreyfus, J.C.; Kaplan, J.C. Heterogeneity of glucose-6-phosphate dehydrogenase deficiency in Algeria. Study in Northern Algeria with description of five new variants. Hum. Genet. 1978, 40, 177–184. [Google Scholar] [CrossRef]
- Saoud, M.Z.; Benkirane, S.; Bennis, F.Z.; Mechal, Y.; Mamad, H.; Dahmani, F.; Berchane, Z.; Woumki, A.; Masrar, A. Déficit en glucose-6-phosphate déshydrogénase: À propos de 54 cas. Hématologie 2019, 25, 16. [Google Scholar] [CrossRef]
- De Araujo, C.; Migot-Nabias, F.; Guitard, J.; Pelleau, S.; Vulliamy, T.; Ducrocq, R. The role of the G6PD AEth376G/968C allele in glucose-6-phosphate dehydrogenase deficiency in the Seerer population of Senegal. Haematologica 2006, 91, 262–263. [Google Scholar]
- Dolo, A.; Maiga, B.; Guindo, A.; Diakité, S.A.; Diakite, M.; Tapily, A.; Traoré, M.; Sangaré, B.; Arama, C.; Daou, M.; et al. Frequency of glucose-6-phosphate dehydrogenase deficiency (A-376/202) in three Malian ethnic groups. Bull. Soc. Pathol. Exot. 2014, 107, 165–170. [Google Scholar] [CrossRef]
- Hadley, T.J. Invasion of erythrocytes by malaria parasites: A cellular and molecular overview. Annu. Rev. Microbiol. 1986, 40, 451–477. [Google Scholar] [CrossRef]
- Langhi, D.M., Jr.; Bordin, J.O. Duffy blood group and malaria. Hematology 2006, 11, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, R. Plasmodium vivax in Africa: Hidden in plain sight? Trends Parasitol. 2007, 23, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Wurtz, N.; Mint Lekweiry, K.; Bogreau, H.; Pradines, B.; Rogier, C.; Ould Mohamed Salem Boukhary, A.; Hafid, J.E.; Ould Ahmedou Salem, M.S.; Trape, J.F.; Basco, L.K.; et al. Vivax malaria in Mauritania includes infection of a Duffy-negative individual. Malar. J. 2011, 10, 336. [Google Scholar] [CrossRef] [Green Version]
- Culleton, R.; Carter, R. African Plasmodium vivax: Distribution and origins. Int. J. Parasitol. 2012, 42, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Golassa, L.; Amenga-Etego, L.; Lo, E.; Amambua-Ngwa, A. The biology of unconventional invasion of Duffy-negative reticulocytes by Plasmodium vivax and its implication in malaria epidemiology and public health. Malar. J. 2020, 19, 299. [Google Scholar] [CrossRef]
- Popovici, J.; Roesch, C.; Rougeron, V. The enigmatic mechanisms by which Plasmodium vivax infects Duffy-negative individuals. PLoS Pathog. 2020, 16, e1008258. [Google Scholar] [CrossRef] [Green Version]
- Ouldabdallahi, M.; Ouldbezeid, M.; Dieye, M.; Yacine, B.; Faye, O. Study of the incidence of malaria in febrile patients and among schoolchildren in the region of Trarza, Islamic Republic of Mauritania. Bull. Soc. Pathol. Exot. 2011, 104, 288–290. [Google Scholar] [CrossRef]
- Ould Ahmedou Salem, M.S.; Basco, L.K.; Ouldabdallahi, M.; Mint Lekweiry, K.; Konaté, L.; Faye, O.; Ould Mohamed Salem Boukhary, A. Malaria-associated morbidity during the rainy season in Saharan and Sahelian zones in Mauritania. Acta Trop. 2015, 152, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ouldabdallahi Moukah, M.; Ba, O.; Ba, H.; Ould Khairy, M.L.; Faye, O.; Bogreau, H.; Simard, F.; Basco, L.K. Malaria in three epidemiological strata in Mauritania. Malar. J. 2016, 15, 204. [Google Scholar] [CrossRef] [Green Version]
- Ba, H.; Duffy, C.W.; Ahouidi, A.D.; Deh, Y.B.; Diallo, M.Y.; Tandia, A.; Conway, D.J. Widespread distribution of Plasmodium vivax malaria in Mauritania on the interface of the Maghreb and West Africa. Malar. J. 2016, 15, 80. [Google Scholar] [CrossRef] [Green Version]
- Ba, H.; Auburn, S.; Jacob, C.G.; Goncalves, S.; Duffy, C.W.; Stewart, L.B.; Price, R.N.; Deh, Y.B.; Diallo, M.Y.; Tandia, A.; et al. Multi-locus genotyping reveals established endemicity of a geographically distinct Plasmodium vivax population in Mauritania, West Africa. PLoS Negl. Trop. Dis. 2020, 14, e0008945. [Google Scholar] [CrossRef]
- Diallo, S.M.; Bogreau, H.; Papa Mze, N.; Ould Ahmedou Salem, M.S.; Ould Khairy, M.L.; Parola, P.; Basco, L.; Ould Mohamed Salem Boukhary, A. Malaria epidemiology in Kobeni department, southeastern Mauritania from 2015 to 2017. Infect. Dis. Poverty 2020, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, A.; Beutler, E.; Motulsky, A.G. Human glucose-6-phosphate dehydrogenase variants. Bull. World Health Organ. 1971, 45, 243–253. [Google Scholar] [PubMed]
- World Health Organization. Malaria Policy Advisory Committee Meeting. Point-of-care G6PD testing to support safe use of primaquine for the treatment of vivax malaria. In WHO Evidence Review Group Meeting Report, 8–9 October 2014; WHO/HTM/GMP/MPAC/2015.6; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Howes, R.E.; Chan, E.R.; Rakotomanga, T.A.; Schulte, S.; Gibson, J.; Zikursh, M.; Franchard, T.; Ramiranirina, B.; Ratsimbasoa, A.; Zimmerman, P.A. Prevalence and genetic variants of G6PD deficiency among two Malagasy populations living in Plasmodium vivax-endemic areas. Malar. J. 2017, 16, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Office National de la Statistique. Annuaire statistique 2019. Ministère des Affaires Economiques et de Promotion des Secteurs Productifs. November 2020. Available online: http://www.ons.mr/images/Archive/doc/publication/Annuaire_Statistique_2019.pdf (accessed on 13 March 2021).
- Boushab, M.B.; Ould Ahmedou Salem, M.S.; Ould Mohamed Salem Boukhary, A.; Parola, P.; Basco, L. Clinical features and mortality associated with severe malaria in adults in southern Mauritania. Trop. Med. Infect. Dis. 2021, 22, 1. [Google Scholar] [CrossRef]
- Ouldabdallahi, M.; Alew, I.; Salem, M.S.; Dit Dialaw Ba, M.; Boukhary, A.O.; Khairy, M.L.; Aziz, M.B.; Ringwald, P.; Basco, L.K.; Niang, S.D.; et al. Efficacy of artesunate-amodiaquine for the treatment of acute uncomplicated falciparum malaria in southern Mauritania. Malar. J. 2014, 13, 496. [Google Scholar] [CrossRef]
- Ould Ahmedou Salem, M.S.; Mohamed Lemine, Y.O.; Deida, J.M.; Lemrabott, M.A.; Ouldabdallahi, M.; Ba, M.D.; Boukhary, A.O.; Khairy, M.L.; Abdel Aziz, M.B.; Ringwald, P.; et al. Efficacy of chloroquine for the treatment of Plasmodium vivax in the Saharan zone in Mauritania. Malar. J. 2015, 14, 39. [Google Scholar] [CrossRef] [Green Version]
- Ezz El-Deen, Z.M.; Hussin, N.F.; Abdel Hamid, T.A.; Abdel Migeed, O.R.; Samy, R.M. G6PD deficiency and G6PD (Mediterranean and silent) polymorphisms in Egyptian infants with neonatal hyperbilirubinemia. Lab. Med. 2013, 44, 228–234. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.Y.; Cheng, A.; Lee, A.; Kuang, W.J.; Hillier, L.; Green, P.; Schlessinger, D.; Ciccodicola, A.; D’Urso, M. Sequence of human glucose-6-phosphate dehydrogenase cloned in plasmids and a yeast artificial chromosome. Genomics 1991, 10, 792–800. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Locality | Ethnic Group | N | Genotype | ||
|---|---|---|---|---|---|
| A- | A | B | |||
| Atar | White Moors | 52 | 4 (7.7) | 2 (3.8) | 46 (88.5) |
| Black Moors | 2 | 0 | 0 | 2 (100) | |
| Nouakchott | White Moors | 71 | 1 (1.4) | 2 (2.8) | 68 (95.8) |
| Black Moors | 19 | 1 (5.3) | 0 | 18 (94.7) | |
| Pular | 9 | 0 | 1 (11.1) | 8 (88.9) | |
| Wolof | 2 | 0 | 0 | 2 (100) | |
| Soninke | 1 | 0 | 0 | 1 (100) | |
| Aleg | White Moors | 29 | 0 | 0 | 29 (100) |
| Black Moors | 12 | 0 | 2 (16.7) | 10 (83.3) | |
| Pular | 1 | 0 | 0 | 1 (100) | |
| Wolof | 1 | 1 (100) | 0 | 0 | |
| Kiffa | White Moors | 9 | 0 | 2 (22.2) | 7 (77.8) |
| Black Moors | 2 | 0 | 1 (50.0) | 1 (50.0) | |
| Kobeni | White Moors | 41 | 2 (4.9) | 6 (14.6) | 33 (80.5) |
| Black Moors | 58 | 7 (12.1) | 12 (20.7) | 39 (67.2) | |
| Pular | 21 | 0 | 7 (33.3) | 14 (66.7) | |
| Soninke | 4 | 0 | 1 (25.0) | 3 (75.0) | |
| Rosso | White Moors | 41 | 3 (7.3) | 3 (7.3) | 35 (85.4) |
| Black Moors | 47 | 7 (14.9) | 11 (23.4) | 29 (61.7) | |
| Pular | 19 | 2 (10.5) | 2 (10.5) | 15 (79.0) | |
| Wolof | 5 | 0 | 0 | 5 (100) | |
| Soninke | 1 | 0 | 0 | 1 (100) | |
| Total | 447 | 28 (6.3) | 52 (11.6) | 367 (82.1) | |
| Locality | Ethnic Group | N | Genotype | |||||
|---|---|---|---|---|---|---|---|---|
| A-A- | AA- | BA- | AA | BB | BA | |||
| Atar | White Moors | 47 | 0 | 0 | 0 | 1 (2.1) | 44 (93.6) | 2 (4.3) |
| Nouakchott | White Moors | 83 | 1 (1.2) | 0 | 0 | 1 (1.2) | 70 (84.3) | 11 (13.3) |
| Black Moors | 31 | 1 (3.2) | 1 (3.2) | 0 | 1 (3.2) | 21 (67.8) | 7 (22.6) | |
| Pular | 5 | 1 (20.0) | 0 | 0 | 0 | 4 (80.0) | 0 | |
| Wolof | 3 | 0 | 0 | 0 | 1 (33.3) | 2 (66.7) | 0 | |
| Soninke | 2 | 0 | 0 | 0 | 0 | 2 (100) | 0 | |
| Aleg | White Moors | 23 | 1 (4.3) | 0 | 0 | 0 | 21 (91.4) | 1 (4.3) |
| Black Moors | 11 | 0 | 0 | 1 9.1) | 2 (18.2) | 6 (54.5) | 2 (18.2) | |
| Pular | 2 | 0 | 0 | 1 (50.0) | 0 | 1 (50.0) | 0 | |
| Kiffa | White Moors | 5 | 0 | 0 | 0 | 0 | 5 (100) | 0 |
| Black Moors | 3 | 0 | 0 | 0 | 0 | 1 (33.3) | 2 (66.7) | |
| Pular | 1 | 0 | 0 | 0 | 0 | 0 | 1 (100) | |
| Kobeni | White Moors | 57 | 1 (1.8) | 1 (1.8) | 3 (5.2) | 1 (1.8) | 48 (84.2) | 3 (5.2) |
| Black Moors | 77 | 1 (1.3) | 4 (5.2) | 10 (13.0) | 6 (7.8) | 38 (49.3) | 18 (23.4) | |
| Pular | 48 | 0 | 2 (4.2) | 2 (4.2) | 0 | 31 (64.6) | 13 (27.0) | |
| Wolof | 2 | 0 | 0 | 0 | 0 | 2 (100) | 0 | |
| Soninke | 15 | 0 | 1 (6.7) | 0 | 1 (6.7) | 10 (66.6) | 3 (20.0) | |
| Rosso | White Moors | 41 | 0 | 0 | 2 (4.9) | 2 (4.9) | 29 (70.7) | 8 (19.5) |
| Black Moors | 54 | 1 (1.9) | 1 (1.9) | 4 (7.4) | 4 (7.4) | 30 (55.5) | 14 (25.9) | |
| Pular | 28 | 0 | 1 (3.6) | 5 (17.9) | 2 (7.1) | 16 (57.1) | 4 (14.3) | |
| Wolof | 11 | 1 (9.1) | 0 | 1 (9.1) | 2 (18.2) | 4 (36.4) | 3 (27.2) | |
| Total | 549 | 8 (1.5) | 11 (2.0) | 29 (5.3) | 24 (4.4) | 385 (70.1) | 92 (16.7) | |
| Ethnic Group | n/N (%) | G6PD*A- Allelic Variants (n; % Among Men with African-Type G6PD Genotypes) | |||
|---|---|---|---|---|---|
| 202A | 542T | 680T | 968C | ||
| White Moors | 10/243 (4.1) | 6 (21.4) | 2 (7.1) | 0 | 2 (7.1) |
| Black Moors | 15/140 (10.7) | 12 (42.8) | 0 | 0 | 3 (10.7) |
| Pular | 2/50 (4.0) | 0 | 0 | 0 | 2 (7.1) |
| Wolof | 1/8 (12.5) | 1 (3.6) | 0 | 0 | 0 |
| Soninke | 0/6 (0) | 0 | 0 | 0 | 0 |
| Total | 28/447 (6.3) | 19 | 2 | 0 | 7 |
| Ethnic Group | n/N (%) | G6PD*A- Allelic Variants (n; % Among Females with the African-Type G6PD Genotypes) | ||||
|---|---|---|---|---|---|---|
| Heterozygote | Homozygote | |||||
| 202G/A | 542A/T | 968T/C | 202A/A | 968C/C | ||
| White Moors | 9/256 (3.5) | 5 (10.4) | 1 (2.1) | 0 | 2 (4.2) | 1 (2.1) |
| Black Moors | 24/176 (13.6) | 14 (29.2) | 1 (2.1) | 6 (12.5) | 1 (2.1) | 2 (4.2) |
| Pular | 12/84 (14.3) | 3 (6.2) | 0 | 8 (16.7) | 0 | 1 (2.1) |
| Wolof | 2/16 (12.5) | 1 (2.1) | 0 | 0 | 0 | 1 (2.1) |
| Soninke | 1/17 (5.9) | 0 | 0 | 1 (2.1) | 0 | 0 |
| Total | 48/549 (8.7) | 23 (47.9) | 2 (4.2) | 15 (31.2) | 3 (6.2) | 5 (10.4) |
| Primers | Primer Sequence | Tm (°C) | Size (bp) | Enzyme |
|---|---|---|---|---|
| Exon 5 | ||||
| A376G_F | 5′-CCCAGGCCACCCCAGAGGAGA-3′ | 58 | 308 | Fok I |
| A376G_R | 5′-CGGCCCCGGACACGCTCATAG-3′ | |||
| Exons 6-7 | ||||
| C563T_F | 5′-ACTCCCCGAAGAGGGGTTCAAGG-3′ | 62 | 547 | Mbo II (C563T) |
| C563T_R | 5′-CCAGCCTCCCAGGAGAGAGGAAG-3′ | Acc III (G542T) Bst NI (G680T) | ||
| Exon 4 | ||||
| G202A_F | 5′-CCACCACTGCCCCTGTGACCT-3′ | 65 | 216 | Nla III |
| G202A_R | 5′-GGCCCTGACACCACCCACCTT-3 | |||
| Exon 9 | ||||
| T968C_F | 5′-TCCCTGCACCCCAACTCAAC-3′ | 65 | 283 | Nci I |
| T968C_R | 5′-CCAGTTCTGCCTTGCTGGGC-3′ | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djigo, O.K.M.; Ould Ahmedou Salem, M.S.; Diallo, S.M.; Bollahi, M.A.; Boushab, B.M.; Garre, A.; Papa Mze, N.; Basco, L.; Briolant, S.; Ould Mohamed Salem Boukhary, A. Molecular Epidemiology of G6PD Genotypes in Different Ethnic Groups Residing in Saharan and Sahelian Zones of Mauritania. Pathogens 2021, 10, 931. https://doi.org/10.3390/pathogens10080931
Djigo OKM, Ould Ahmedou Salem MS, Diallo SM, Bollahi MA, Boushab BM, Garre A, Papa Mze N, Basco L, Briolant S, Ould Mohamed Salem Boukhary A. Molecular Epidemiology of G6PD Genotypes in Different Ethnic Groups Residing in Saharan and Sahelian Zones of Mauritania. Pathogens. 2021; 10(8):931. https://doi.org/10.3390/pathogens10080931
Chicago/Turabian StyleDjigo, Oum Kelthoum Mamadou, Mohamed Salem Ould Ahmedou Salem, Sileye Mamadou Diallo, Mohamed Abdallahi Bollahi, Boushab Mohamed Boushab, Aymeric Garre, Nasserdine Papa Mze, Leonardo Basco, Sébastien Briolant, and Ali Ould Mohamed Salem Boukhary. 2021. "Molecular Epidemiology of G6PD Genotypes in Different Ethnic Groups Residing in Saharan and Sahelian Zones of Mauritania" Pathogens 10, no. 8: 931. https://doi.org/10.3390/pathogens10080931