
Whole-Genome Analysis of Multidrug-Resistant Salmonella Enteritidis Strains Isolated from Poultry Sources in Korea

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Salmonella Isolation
2.3. Antibiotic Susceptibility Test
2.4. Extraction and WGS of MDR Salmonella Genomic DNA
2.5. Assembly, Polishing, and Annotation of MDR Salmonella DNA
2.6. Data Analysis
3. Results
3.1. Prevalence of Salmonella spp.
3.2. Antibiotic Resistance Profiles of the Isolated Salmonella spp.
3.3. Results of Whole-Genome Sequencing and In-Silico Serotyping of MDR Salmonella spp.
3.4. Antibiotic Resistance Profiles of MDR Salmonella Enteritidis
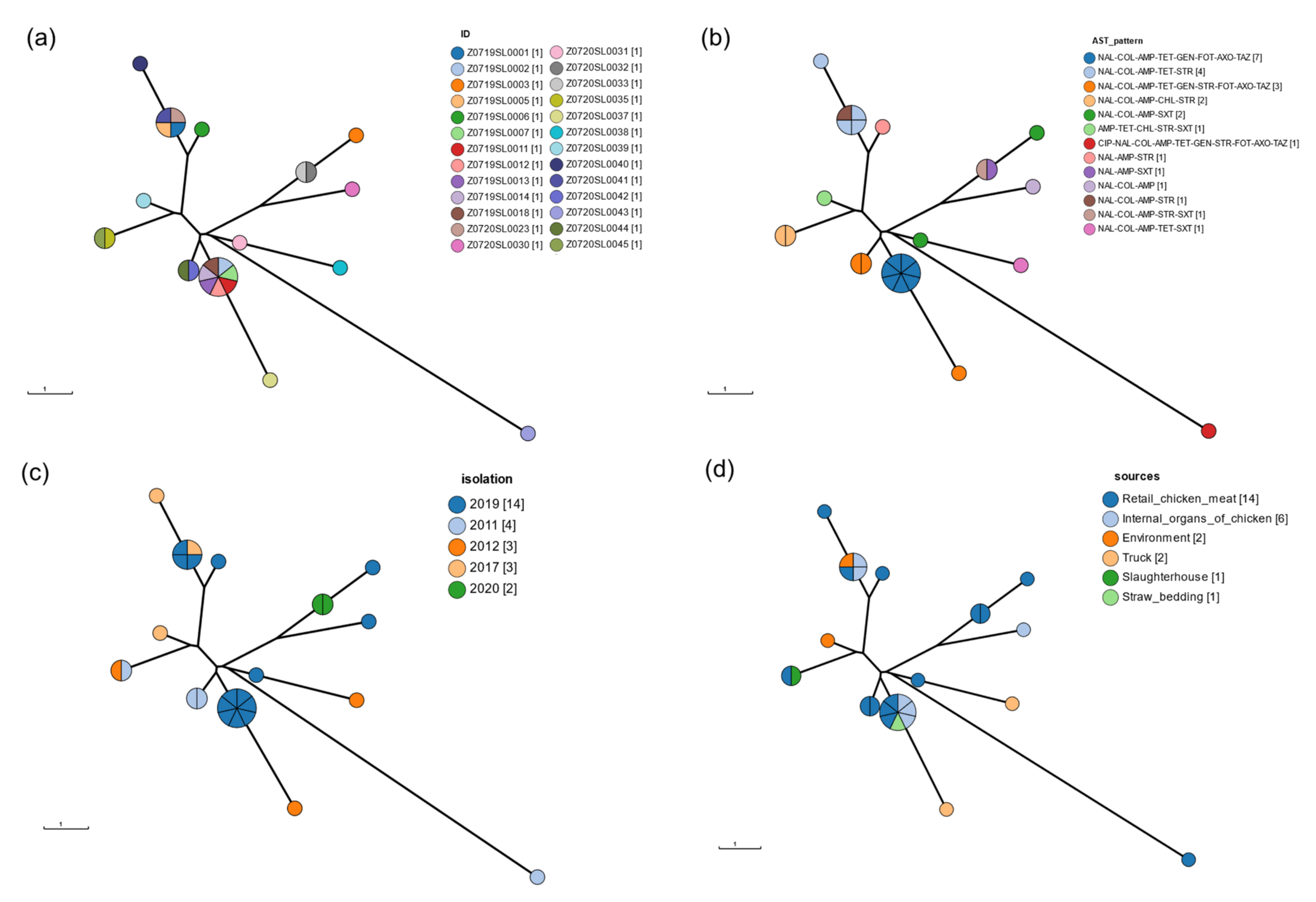
3.5. CgMLST
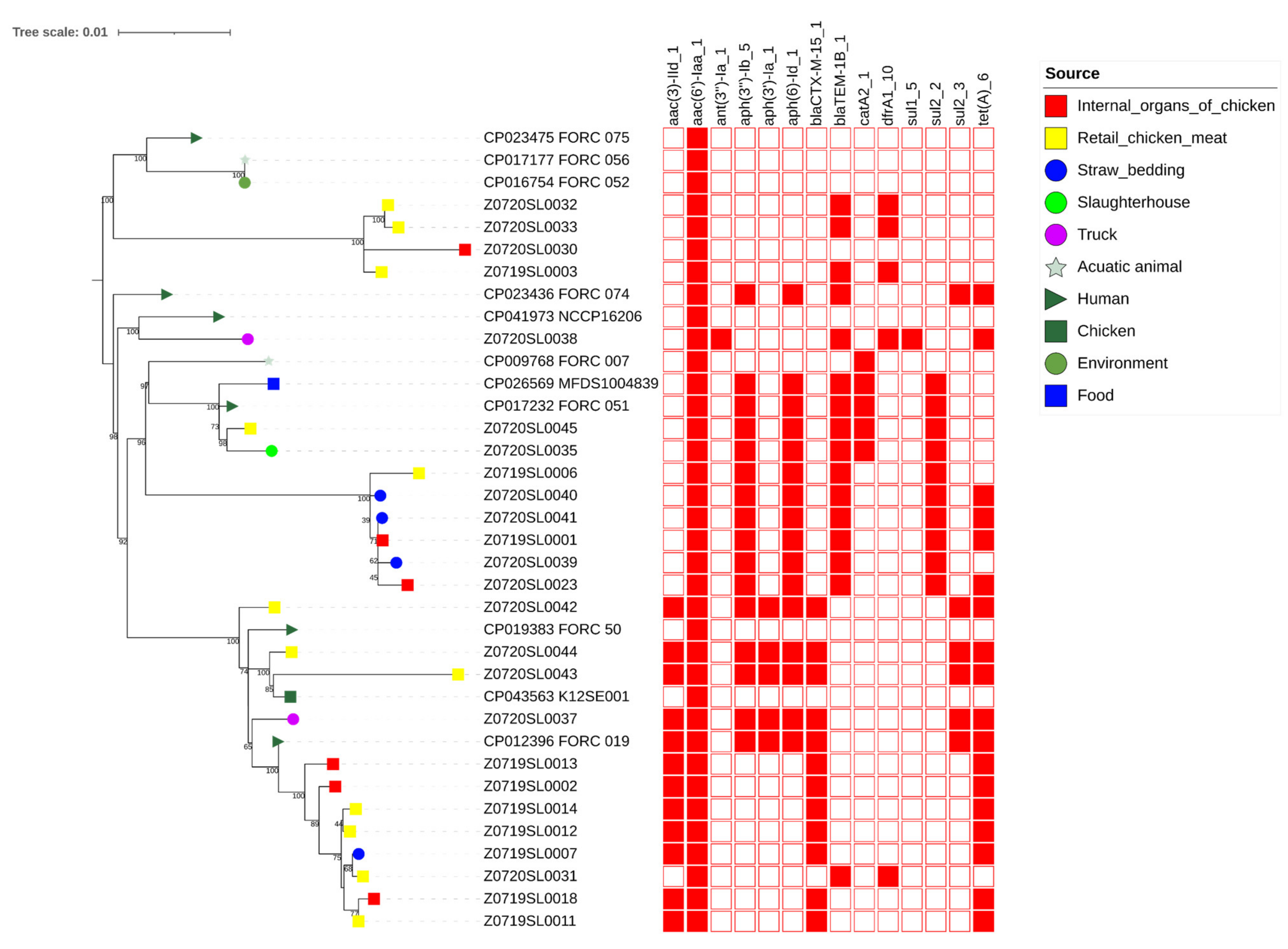
3.6. wgSNP Phylogenetic Analysis of the Salmonella Enteritidis Strains
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dewey-Mattia, D.; Manikonda, K.; Hall, A.J.; Wise, M.E.; Crowe, S.J. Surveillance for Foodborne Disease Outbreaks—United States, 2009–2015. MMWR Surveill. Summ. 2018, 67, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Eng, S.-K.; Pusparajah, P.; Mutalib, N.-S.A.; Ser, H.-L.; Chan, K.-G.; Lee, L.-H. Salmonella: A Review on Pathogenesis, Epidemiology and Antibiotic Resistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar] [CrossRef] [Green Version]
- Omwandho, C.O.A.; Kubota, T. Salmonella Enterica Serovar Enteritidis: A Mini-Review of Contamination Routes and Limitations to Effective Control. Jpn. Agric. Res. Q. 2010, 44, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Hogue, A.; White, P.; Guard-Petter, J.; Schlosser, W.; Gast, R.; Ebel, E.; Farrar, J.; Gomez, T.; Madden, J.; Madison, M.; et al. Epidemiology and Control of Egg-Associated Salmonella Enteriditis in the United States of America. Rev. Sci. Tech. 1997, 16, 542–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sin, M.; Yoon, S.; Kim, Y.B.; Noh, E.B.; Seo, K.W.; Lee, Y.J. Molecular Characteristics of Antimicrobial Resistance Determinants and Integrons in Salmonella Isolated from Chicken Meat in Korea. J. Appl. Poult. Res. 2020, 29, 502–514. [Google Scholar] [CrossRef]
- Choi, S.-W.; Ha, J.-S.; Kim, B.-Y.; Lee, D.-H.; Park, J.-K.; Youn, H.-N.; Hong, Y.-H.; Lee, S.-B.; Lee, J.-B.; Park, S.-Y.; et al. Prevalence and Characterization of Salmonella Species in Entire Steps of a Single Integrated Broiler Supply Chain in Korea. Poult. Sci. 2014, 93, 1251–1257. [Google Scholar] [CrossRef] [PubMed]
- Seo, K.W.; Lee, Y.J. Prevalence and Characterization of β-Lactamases Genes and Class 1 Integrons in Multidrug-Resistant Escherichia coli Isolates from Chicken Meat in Korea. Microb. Drug Resist. 2018, 24, 1599–1606. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Li, X.; Liao, Q.; Li, F.; Zhang, J.; Kan, B. The Emergence and Outbreak of Multidrug-Resistant Typhoid Fever in China. Emerg. Microbes Infect. 2019, 5, 1–6. [Google Scholar] [CrossRef]
- Crump, J.A.; Mintz, E.D. Global Trends in Typhoid and Paratyphoid Fever. Clin. Infect. Dis. 2010, 50, 241–246. [Google Scholar] [CrossRef]
- Tamang, M.D.; Nam, H.-M.; Kim, T.-S.; Jang, G.-C.; Jung, S.-C.; Lim, S.-K. Emergence of Extended-Spectrum β-Lactamase (CTX-M-15 and CTX-M-14)-Producing Nontyphoid Salmonella with Reduced Susceptibility to Ciprofloxacin among Food Animals and Humans in Korea. J. Clin. Microbiol. 2011, 49, 2671–2675. [Google Scholar] [CrossRef] [Green Version]
- Dessie, H.K.; Bae, D.H.; Lee, Y.J. Characterization of Integrons and Their Cassettes in Escherichia coli and Salmonella Isolates from Poultry in Korea. Poult. Sci. 2013, 92, 3036–3043. [Google Scholar] [CrossRef] [PubMed]
- Hyeon, J.-Y.; Chon, J.-W.; Hwang, I.-G.; Kwak, H.-S.; Kim, M.-S.; Kim, S.-K.; Choi, I.-S.; Song, C.-S.; Park, C.; Seo, K.-H. Prevalence, Antibiotic Resistance, and Molecular Characterization of Salmonella Serovars in Retail Meat Products. J. Food Prot. 2016, 74, 161–166. [Google Scholar] [CrossRef]
- Boxrud, D.; Pederson-Gulrud, K.; Wotton, J.; Medus, C.; Lyszkowicz, E.; Besser, J.; Bartkus, J.M. Comparison of Multiple-Locus Variable-Number Tandem Repeat Analysis, Pulsed-Field Gel Electrophoresis, and Phage Typing for Subtype Analysis of Salmonella Enterica Serotype Enteritidis. J. Clin. Microbiol. 2006, 45, 536–543. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-H.; Hyeon, J.-Y.; Kim, J.; Kim, J.S.; Kim, S.J.; Jeon, S.-E.; Choi, S.-W.; Hong, W.-T.; Song, C.-S.; Lee, S.-W. Close Genetic Relationship between Salmonella Enterica Serovar Enteritidis Isolated from Patients with Diarrhoea and Poultry in the Republic of Korea. Clin. Microbiol. Infect. 2015, 21, e68–e70. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.-S.; Oh, J.-Y.; Kwon, Y.-K.; Lee, D.-Y.; Jeong, O.-M.; Choi, B.-K.; Youn, S.-Y.; Jeon, B.-W.; Lee, H.-J.; Lee, H.-S. Public Health Significance of Major Genotypes of Salmonella Enterica Serovar Enteritidis Present in Both Human and Chicken Isolates in Korea. Res. Vet. Sci. 2017, 112, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Allard, M.W.; Luo, Y.; Strain, E.; Pettengill, J.; Timme, R.; Wang, C.; Li, C.; Keys, C.E.; Zheng, J.; Stones, R.; et al. On the Evolutionary History, Population Genetics and Diversity among Isolates of Salmonella Enteritidis PFGE Pattern JEGX01.0004. PLoS ONE 2013, 8, e55254. [Google Scholar] [CrossRef]
- Saeed, A.M.; Walk, S.T.; Arshad, M.; Whittam, T.S. Clonal Structure and Variation in Virulence of Salmonella Enteritidis Isolated from Mice, Chickens, and Humans. J. AOAC Int. 2006, 89, 504–511. [Google Scholar] [CrossRef]
- Taylor, A.J.; Lappi, V.; Wolfgang, W.J.; Lapierre, P.; Palumbo, M.J.; Medus, C.; Boxrud, D. Characterization of Foodborne Outbreaks of Salmonella Enterica Serovar Enteritidis with Whole-Genome Sequencing Single Nucleotide Polymorphism-Based Analysis for Surveillance and Outbreak Detection. J. Clin. Microbiol. 2015, 53, 3334–3340. [Google Scholar] [CrossRef] [Green Version]
- Jackson, B.R.; Tarr, C.; Strain, E.; Jackson, K.A.; Conrad, A.; Carleton, H.; Katz, L.S.; Stroika, S.; Gould, L.H.; Mody, R.K.; et al. Implementation of Nationwide Real-Time Whole-Genome Sequencing to Enhance Listeriosis Outbreak Detection and Investigation. Clin. Infect. Dis. 2016, 63, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Den Bakker, H.C.; Allard, M.W.; Bopp, D.; Brown, E.W.; Fontana, J.; Iqbal, Z.; Kinney, A.; Limberger, R.; Musser, K.A.; Shudt, M.; et al. Rapid Whole-Genome Sequencing for Surveillance of Salmonella Enterica Serovar Enteritidis. Emerg. Infect. Dis. 2014, 20, 1306–1314. [Google Scholar] [CrossRef]
- Brown, E.; Dessai, U.; McGarry, S.; Gerner-Smidt, P. Use of Whole-Genome Sequencing for Food Safety and Public Health in the United States. Foodborne Pathog. Dis. 2019, 16, 441–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Bakker, H.C.; Switt, A.I.M.; Cummings, C.A.; Hoelzer, K.; Degoricija, L.; Rodriguez-Rivera, L.D.; Wright, E.M.; Fang, R.; Davis, M.; Root, T.; et al. A Whole-Genome Single Nucleotide Polymorphism-Based Approach to Trace and Identify Outbreaks Linked to a Common Salmonella Enterica Subsp. Enterica Serovar Montevideo Pulsed-Field Gel Electrophoresis Type. Appl. Environ. Microbiol. 2011, 77, 8648–8655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiden, M.C.J.; Bygraves, J.A.; Feil, E.; Morelli, G.; Russell, J.E.; Urwin, R.; Zhang, Q.; Zhou, J.; Zurth, K.; Caugant, D.A.; et al. Multilocus Sequence Typing: A Portable Approach to the Identification of Clones within Populations of Pathogenic Microorganisms. Proc. Natl. Acad. Sci. USA 1998, 95, 3140–3145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, M.; Zhao, S.; Pettengill, J.; Luo, Y.; Monday, S.R.; Abbott, J.; Ayers, S.L.; Cinar, H.N.; Muruvanda, T.; Li, C.; et al. Comparative Genomic Analysis and Virulence Differences in Closely Related Salmonella Enterica Serotype Heidelberg Isolates from Humans, Retail Meats, and Animals. Genome Biol. Evol. 2014, 6, 1046–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bratcher, H.B.; Corton, C.; Jolley, K.A.; Parkhill, J.; Maiden, M.C. A Gene-by-Gene Population Genomics Platform: De Novo Assembly, Annotation and Genealogical Analysis of 108 Representative Neisseria meningitidis Genomes. BMC Genom. 2014, 15, 1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leekitcharoenphon, P.; Hendriksen, R.S.; Hello, S.L.; Weill, F.-X.; Baggesen, D.L.; Jun, S.-R.; Ussery, D.W.; Lund, O.; Crook, D.W.; Wilson, D.J.; et al. Global Genomic Epidemiology of Salmonella Enterica Serovar Typhimurium DT104. Appl. Environ. Microbiol. 2016, 82, 2516–2526. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Desai, P.T.; den Bakker, H.C.; Mikoleit, M.; Tolar, B.; Trees, E.; Hendriksen, R.S.; Frye, J.G.; Porwollik, S.; Weimer, B.C.; et al. Genomic Epidemiology of Salmonella Enterica Serotype Enteritidis Based on Population Structure of Prevalent Lineages. Emerg. Infect. Dis. 2014, 20, 1481–1489. [Google Scholar] [CrossRef] [Green Version]
- Toro, M.; Retamal, P.; Ayers, S.; Barreto, M.; Allard, M.; Brown, E.W.; Gonzalez-Escalona, N. Whole-Genome Sequencing Analysis of Salmonella Enterica Serovar Enteritidis Isolates in Chile Provides Insights into Possible Transmission between Gulls, Poultry, and Humans. Appl. Environ. Microbiol. 2016, 82, 6223–6232. [Google Scholar] [CrossRef] [Green Version]
- Pearce, M.E.; Alikhan, N.-F.; Dallman, T.J.; Zhou, Z.; Grant, K.; Maiden, M.C.J. Comparative Analysis of Core Genome MLST and SNP Typing within a European Salmonella Serovar Enteritidis Outbreak. Int. J. Food Microbiol. 2018, 274, 1–11. [Google Scholar] [CrossRef]
- Park, A.K.; Shin, E.; Kim, S.; Park, J.; Jeong, H.J.; Chun, J.-H.; Hwang, K.J.; Kim, J. Traveller-Associated High-Level Ciprofloxacin-Resistant Salmonella Enterica Serovar Kentucky in the Republic of Korea. J. Glob. Antimicrob. Resist. 2020, 22, 190–194. [Google Scholar] [CrossRef]
- Kim, S.; Kim, E.; Park, S.; Hahn, T.-W.; Yoon, H. Genomic Approaches for Understanding the Characteristics of Salmonella Enterica subsp. Enterica Serovar Typhimurium ST1120, Isolated from Swine Feces in Korea. J. Microbiol. Biotechnol. 2017, 27, 1983–1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyeon, J.-Y.; Li, S.; Mann, D.A.; Zhang, S.; Kim, K.-J.; Lee, D.-H.; Deng, X.; Song, C.-S. Whole-Genome Sequencing Analysis of Salmonella Enterica Serotype Enteritidis Isolated from Poultry Sources in South Korea, 2010–2017. Pathogens 2021, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Halatsi, K.; Oikonomou, I.; Lambiri, M.; Mandilara, G.; Vatopoulos, A.; Kyriacou, A. PCR Detection of Salmonella spp. Using Primers Targeting the Quorum Sensing Gene sdiA. FEMS Microbiol. Lett. 2006, 259, 201–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; CLSI Document M100-S29; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of Long, Error-Prone Reads Using Repeat Graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Zhang, S.; Yin, Y.; Jones, M.B.; Zhang, Z.; Kaiser, B.L.D.; Dinsmore, B.A.; Fitzgerald, C.; Fields, P.I.; Deng, X. Salmonella Serotype Determination Utilizing High-Throughput Genome Sequencing Data. J. Clin. Microbiol. 2015, 53, 1685–1692. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Alikhan, N.-F.; Sergeant, M.J.; Luhmann, N.; Vaz, C.; Francisco, A.P.; Carriço, J.A.; Achtman, M. GrapeTree: Visualization of Core Genomic Relationships among 100,000 Bacterial Pathogens. Genome Res. 2018, 28, 1395–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of Acquired Antimicrobial Resistance Genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Gardner, S.N.; Slezak, T.; Hall, B.G. KSNP3.0: SNP Detection and Phylogenetic Analysis of Genomes without Genome Alignment or Reference Genome. Bioinformatics 2015, 31, 2877–2878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, L.-H.; Chiu, C.-H.; Chu, C.; Ou, J.T. Antimicrobial Resistance in Nontyphoid Salmonella Serotypes: A Global Challenge. Clin. Infect. Dis. 2004, 39, 546–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, S.D.; Flores, F.S.; dos Santos, L.R.; Brandelli, A. Antimicrobial Resistance in Salmonella Enteritidis Strains Isolated from Broiler Carcasses, Food, Human and Poultry-Related Samples. Int. J. Food Microbiol. 2005, 97, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Thung, T.Y.; Mahyudin, N.A.; Basri, D.F.; Radzi, C.W.J.W.M.; Nakaguchi, Y.; Nishibuchi, M.; Radu, S. Prevalence and Antibiotic Resistance of Salmonella Enteritidis and Salmonella Typhimurium in Raw Chicken Meat at Retail Markets in Malaysia. Poult. Sci. 2016, 95, 1888–1893. [Google Scholar] [CrossRef] [PubMed]
- Lamas, A.; Fernandez-No, I.C.; Miranda, J.M.; Vázquez, B.; Cepeda, A.; Franco, C.M. Prevalence, Molecular Characterization and Antimicrobial Resistance of Salmonella Serovars Isolated from Northwestern Spanish Broiler Flocks (2011–2015). Poult. Sci. 2016, 95, 2097–2105. [Google Scholar] [CrossRef]
- Cui, M.; Xie, M.; Qu, Z.; Zhao, S.; Wang, J.; Wang, Y.; He, T.; Wang, H.; Zuo, Z.; Wu, C. Prevalence and Antimicrobial Resistance of Salmonella Isolated from an Integrated Broiler Chicken Supply Chain in Qingdao, China. Food Control 2016, 62, 270–276. [Google Scholar] [CrossRef]
- Ahmed, M.A.E.-G.E.-S.; Zhong, L.-L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.-B. Colistin and Its Role in the Era of Antibiotic Resistance: An Extended Review (2000–2019). Emerg. Microbes Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef] [Green Version]
- Rhouma, M.; Beaudry, F.; Thériault, W.; Letellier, A. Colistin in Pig Production: Chemistry, Mechanism of Antibacterial Action, Microbial Resistance Emergence, and One Health Perspectives. Front. Microbiol. 2016, 7, 1789. [Google Scholar] [CrossRef]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Negrea, A.; Rhen, M.; Andersson, D.I. Genetic Analysis of Colistin Resistance in Salmonella Enterica Serovar Typhimurium. Antimicrob. Agents Chemother. 2009, 53, 2298–2305. [Google Scholar] [CrossRef] [Green Version]
- Sekyere, J.O.; Govinden, U.; Bester, L.A.; Essack, S.Y. Colistin and Tigecycline Resistance in Carbapenemase-producing Gram-negative Bacteria: Emerging Resistance Mechanisms and Detection Methods. J. Appl. Microbiol. 2016, 121, 601–617. [Google Scholar] [CrossRef]
- Morales, A.S.; de Araújo, J.F.; de Moura Gomes, V.T.; Costa, A.T.R.; dos Prazeres Rodrigues, D.; Ferreira, T.S.P.; de Lima Filsner, P.H.N.; Felizardo, M.R.; Moreno, A.M. Colistin Resistance in Escherichia coli and Salmonella Enterica Strains Isolated from Swine in Brazil. Sci. World J. 2012, 2012, 109795. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Feng, Y.; Liu, L.; Wei, L.; Kang, M.; Zong, Z. Identification of Novel Mobile Colistin Resistance Gene mcr-10. Emerg. Microbes Infect. 2020, 9, 508–516. [Google Scholar] [CrossRef] [Green Version]
- Rule, R.; Mbelle, N.; Sekyere, J.O.; Kock, M.; Hoosen, A.; Said, M. A Rare Case of Colistin-Resistant Salmonella Enteritidis Meningitis in an HIV-Seropositive Patient. BMC Infect. Dis. 2019, 19, 806. [Google Scholar] [CrossRef]
- White, D.G.; Zhao, S.; Sudler, R.; Ayers, S.; Friedman, S.; Chen, S.; McDermott, P.F.; McDermott, S.; Wagner, D.D.; Meng, J. The Isolation of Antibiotic-Resistant Salmonella from Retail Ground Meats. N. Engl. J. Med. 2001, 345, 1147–1154. [Google Scholar] [CrossRef] [Green Version]
- Williamson, D.A.; Lane, C.R.; Easton, M.; Valcanis, M.; Strachan, J.; Veitch, M.G.; Kirk, M.D.; Howden, B.P. Increasing Antimicrobial Resistance in Nontyphoidal Salmonella Isolates in Australia from 1979 to 2015. Antimicrob. Agents Chemother. 2018, 62, e02012-17. [Google Scholar] [CrossRef] [Green Version]
- Trongjit, S.; Angkititrakul, S.; Tuttle, R.E.; Poungseree, J.; Padungtod, P.; Chuanchuen, R. Prevalence and Antimicrobial Resistance in Salmonella Enterica Isolated from Broiler Chickens, Pigs and Meat Products in Thailand-Cambodia Border Provinces. Microbiol. Immunol. 2017, 61, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Strain | CIP | NAL | IMI | COL | AMP | TET | CHL | AZI | GEN | STR | AMI | SXT | FOT | AXO | FOX | TAZ | Number of Resistance | Number of Antibiotics Classes | Isolation Year | Source of Isolation |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Z0719SL0002 | I | R | S | R | R | R | S | S | R | S | S | S | R | R | S | R | 8 | 6 | 2019 | Internal organs of chicken |
| Z0719SL0007 | I | R | S | R | R | R | S | S | R | S | S | S | R | R | S | R | 8 | 6 | 2019 | Straw bedding samples |
| Z0719SL0011 | I | R | S | R | R | R | S | S | R | S | S | S | R | R | S | R | 8 | 6 | 2019 | Retail chicken meat |
| Z0719SL0012 | I | R | S | R | R | R | S | S | R | S | S | S | R | R | S | R | 8 | 6 | 2019 | Retail chicken meat |
| Z0719SL0013 | I | R | S | R | R | R | S | S | R | S | S | S | R | R | S | R | 8 | 6 | 2019 | Internal organs of chicken |
| Z0719SL0014 | I | R | S | R | R | R | S | S | R | S | S | S | R | R | S | R | 8 | 6 | 2019 | Retail chicken meat |
| Z0719SL0018 | I | R | S | R | R | R | S | S | R | S | S | S | R | R | S | R | 8 | 6 | 2019 | Internal organs of chicken |
| Z0720SL0031 | I | R | S | R | R | R | S | S | R | S | S | S | R | R | S | R | 8 | 6 | 2019 | Retail chicken meat |
| Z0719SL0004 | I | R | S | S | R | S | S | S | S | S | S | S | R | R | R | R | 6 | 3 | 2019 | Retail chicken meat |
| Z0719SL0001 | I | R | S | R | R | R | S | S | S | R | S | S | S | S | S | S | 5 | 5 | 2019 | Internal organs of chicken |
| Z0720SL0023 | I | R | S | R | R | R | S | S | S | R | S | S | S | S | S | S | 5 | 5 | 2019 | Internal organs of chicken |
| Z0719SL0008 | R | R | S | S | R | S | R | S | S | S | S | S | S | S | R | S | 5 | 4 | 2019 | Straw bedding samples |
| Z0719SL0009 | I | R | S | S | R | R | R | S | S | S | S | R | S | S | S | S | 5 | 5 | 2019 | Slaughterhouse |
| Z0719SL0010 | I | R | S | S | R | R | R | S | S | S | S | R | S | S | S | S | 5 | 5 | 2019 | Slaughterhouse |
| Z0719SL0021 | I | R | S | S | R | R | R | S | S | S | S | R | S | S | S | S | 5 | 5 | 2019 | Slaughterhouse |
| Z0720SL0026 | I | R | S | S | R | R | R | S | S | S | S | R | S | S | S | S | 5 | 5 | 2019 | Slaughterhouse |
| Z0719SL0003 | I | R | S | R | R | S | S | S | S | S | S | R | S | S | S | S | 4 | 4 | 2019 | Retail chicken meat |
| Z0719SL0005 | I | R | S | R | R | S | S | S | S | R | S | S | S | S | S | S | 4 | 4 | 2019 | Retail chicken meat |
| Z0719SL0017 | R | S | S | S | R | R | I | S | S | R | S | S | S | S | S | S | 4 | 4 | 2019 | Slaughterhouse |
| Z0719SL0022 | R | S | S | S | R | R | I | S | S | R | S | S | S | S | S | S | 4 | 4 | 2019 | Slaughterhouse |
| Z0720SL0029 | I | R | S | S | R | S | S | S | S | S | S | S | R | R | I | I | 4 | 3 | 2019 | Retail chicken meat |
| Z0719SL0006 | I | R | S | S | R | S | S | S | S | R | S | S | S | S | S | S | 3 | 3 | 2019 | Retail chicken meat |
| Z0719SL0015 | I | S | S | S | R | R | S | S | S | R | S | S | S | S | S | S | 3 | 3 | 2019 | Slaughterhouse |
| Z0719SL0016 | I | S | S | S | R | R | S | S | S | R | S | S | S | S | S | S | 3 | 3 | 2019 | Slaughterhouse |
| Z0719SL0019 | I | S | S | S | R | R | S | S | S | R | S | S | S | S | S | S | 3 | 3 | 2019 | Internal organs of chicken |
| Z0719SL0020 | I | S | S | S | R | R | S | S | S | R | S | S | S | S | S | S | 3 | 3 | 2019 | Slaughterhouse |
| Z0720SL0025 | I | S | S | S | R | R | S | S | S | R | S | S | S | S | S | S | 3 | 3 | 2019 | Slaughterhouse |
| Z0720SL0027 | I | S | S | S | R | R | S | S | S | R | S | S | I | S | S | S | 3 | 3 | 2019 | Slaughterhouse |
| Z0720SL0030 | I | R | S | R | R | S | S | S | S | S | S | S | S | S | S | S | 3 | 3 | 2019 | Internal organs of chicken |
| Z0720SL0028 | I | S | S | S | R | R | S | S | S | R | S | S | S | S | S | S | 3 | 3 | 2019 | Slaughterhouse |
| Z0720SL0033 | I | R | S | R | R | S | S | S | S | R | S | R | S | S | S | S | 5 | 5 | 2020 | Retail chicken meat |
| Z0720SL0034 | R | R | S | S | R | R | I | S | S | R | S | S | S | S | S | S | 5 | 4 | 2020 | Retail chicken meat |
| Z0720SL0032 | I | R | S | S | R | S | S | S | S | S | S | R | S | S | S | S | 3 | 3 | 2020 | Retail chicken meat |
| Z0720SL0035 | I | R | S | R | R | R | S | S | S | R | S | S | S | S | S | S | 5 | 5 | 2012 | Slaughterhouse |
| Z0720SL0037 | I | R | S | R | R | R | S | S | R | R | S | S | R | R | S | R | 9 | 6 | 2012 | Truck |
| Z0720SL0038 | S | S | S | S | R | R | R | S | S | R | S | R | S | S | S | S | 5 | 5 | 2012 | Truck |
| Z0720SL0039 | I | R | S | R | R | R | S | S | S | S | S | R | S | S | S | S | 5 | 5 | 2017 | Environment |
| Z0720SL0040 | I | R | S | R | R | R | S | S | S | R | S | S | S | S | S | S | 5 | 5 | 2017 | Retail chicken meat |
| Z0720SL0041 | I | R | S | R | R | S | R | S | S | R | S | S | S | S | S | S | 5 | 5 | 2017 | Environment |
| Z0720SL0042 | I | R | S | R | R | R | S | S | R | R | S | S | R | R | S | R | 9 | 6 | 2011 | Retail chicken meat |
| Z0720SL0043 | R | R | S | R | R | R | S | S | R | R | S | S | R | R | S | R | 10 | 6 | 2011 | Retail chicken meat |
| Z0720SL0044 | I | R | S | R | R | R | S | S | R | R | S | S | R | R | S | R | 9 | 6 | 2011 | Retail chicken meat |
| Z0720SL0045 | I | R | S | R | R | S | R | S | S | R | S | S | S | S | S | S | 5 | 5 | 2011 | Retail chicken meat |
| Sample Name | Data Output (gb) | Fold Coverage (X) | Chromosome Size (bp) | Number of Plasmid | Serovar |
|---|---|---|---|---|---|
| Z0720SL0023 | 1.8 | 375.0 | 4,783,705 | 1 | Enteritidis |
| Z0719SL0001 | 5.2 | 1083.3 | 4,783,876 | 1 | Enteritidis |
| Z0719SL0004 | 1.8 | 375.0 | 4,673,348 | 0 | Virchow |
| Z0719SL0003 | 6.4 | 1333.3 | 4,679,604 | 3 | Enteritidis |
| Z0719SL0002 | 5.9 | 1229.2 | 4,680,702 | 1 | Enteritidis |
| Z0719SL0005 | 1.75 | 364.6 | 4,779,036 | 1 | Enteritidis |
| Z0719SL0006 | 5.9 | 1229.2 | 4,779,850 | 0 | Enteritidis |
| Z0719SL0007 | 2.8 | 583.3 | 4,681,486 | 1 | Enteritidis |
| Z0719SL0008 | 2.5 | 520.8 | 4,670,331 | 2 | Virchow |
| Z0719SL0011 | 4.5 | 937.5 | 4,681,460 | 1 | Enteritidis |
| Z0719SL0012 | 4.7 | 979.2 | 4,681,475 | 1 | Enteritidis |
| Z0719SL0009 | 2.1 | 437.5 | 4,809,470 | 1 | Albany |
| Z0719SL0010 | 2.1 | 437.5 | 4,809,485 | 2 | Albany |
| Z0719SL0013 | 4.8 | 1000.0 | 4,683,147 | 1 | Enteritidis |
| Z0719SL0014 | 5.3 | 1104.2 | 4,678,918 | 1 | Enteritidis |
| Z0719SL0018 | 1 | 208.3 | 4,681,459 | 1 | Enteritidis |
| Z0719SL0019 | 7 | 1458.3 | 4,843,579 | 1 | Agona |
| Z0720SL0025 | 0.62 | 129.2 | 4,878,409 | 1 | Agona |
| Z0719SL0021 | 2.85 | 593.8 | 4,844,531 | 0 | Albany |
| Z0720SL0026 | 0.42 | 87.5 | 4,844,485 | 0 | Albany |
| Z0720SL0027 | 0.6 | 125.0 | 4,593,080 | 1 | Virchow |
| Z0720SL0028 | 2.09 | 435.4 | 4,677,146 | 2 | Virchow |
| Z0719SL0015 | 2.7 | 562.5 | 4,843,592 | 1 | Agona |
| Z0719SL0016 | 5 | 1041.7 | 4,843,581 | 1 | Agona |
| Z0719SL0017 | 4.9 | 1020.8 | 4,877,928 | 1 | Agona |
| Z0719SL0022 | 2.05 | 427.1 | 4,877,150 | 1 | Agona |
| Z0719SL0020 | 2.05 | 427.1 | 4,878,418 | 1 | Agona |
| Z0720SL0030 | 0.51 | 106.3 | 4,547,043 | 0 | Enteritidis |
| Z0720SL0031 | 0.54 | 112.5 | 4,681,359 | 4 | Enteritidis |
| Z0720SL0032 | 3.3 | 687.5 | 4,679,600 | 4 | Enteritidis |
| Z0720SL0033 | 1.05 | 218.8 | 4,679,611 | 4 | Enteritidis |
| Z0720SL0034 | 1.1 | 229.2 | 4,670,318 | 0 | Virchow |
| Z0720SL0035 | 1.31 | 272.9 | 4,680,380 | 1 | Enteritidis |
| Z0720SL0037 | 1.55 | 322.9 | 4,680,192 | 3 | Enteritidis |
| Z0720SL0038 | 1.56 | 325.0 | 4,680,091 | 2 | Enteritidis |
| Z0720SL0039 | 1.53 | 318.8 | 4,807,544 | 2 | Enteritidis |
| Z0720SL0040 | 2.55 | 531.3 | 4,782,444 | 1 | Enteritidis |
| Z0720SL0041 | 1.32 | 275.0 | 4,783,583 | 1 | Enteritidis |
| Z0720SL0042 | 1.29 | 268.8 | 4,678,693 | 1 | Enteritidis |
| Z0720SL0043 | 1.47 | 306.3 | 4,664,874 | 1 | Enteritidis |
| Z0720SL0044 | 1.36 | 283.3 | 4,680,669 | 1 | Enteritidis |
| Z0720SL0045 | 1.49 | 310.4 | 4,679,466 | 1 | Enteritidis |
| Antibiotic Resistance | Sources of Isolation | Antibiotic Resistance Gene | No. of Isolates | No. of Antibiotics | No. of Classes |
|---|---|---|---|---|---|
| NAL-COL-AMP-TET-GEN-FOT-AXO-TAZ | Retail chicken meat | aac(6′)-Iaa_1 aac(3)-IId_1 blaCTX-M-15_1 tet(A)_6 | 3 | 8 | 6 |
| Straw for bedding | 1 | ||||
| Internal organs of chicken | 3 | ||||
| NAL-COL-AMP-TET-GEN-STR-FOT-AXO-TAZ | Retail chicken meat | aac(6′)-Iaa_1 sul2_3 aph(3″)-Ib_5 aph(6)-Id_1 aph(3′)-Ia_1 aac(3)-IId_1 blaCTX-M-15_1 tet(A)_6 | 2 | 9 | 6 |
| Truck | 1 | ||||
| CIP-NAL-COL-AMP-TET-GEN-STR-FOT-AXO-TAZ | Retail chicken meat | aac(6′)-Iaa_1 aac(3)-IId_1 sul2_3 aph(3″)-Ib_5 aph(6)-Id_1 aph(3′)-Ia_1 tet(A)_6 blaCTX-M-15_1 | 1 | 10 | 6 |
| NAL-COL-AMP-TET-STR | Retail chicken meat | aac(6′)-Iaa_1 blaTEM-1B_1 aph(6)-Id_1 aph(3″)-Ib_5 tet(A)_6 aph(6)-Id_1 aph(3″)-Ib_5 sul2_2 | 1 | 5 | 5 |
| Environment | 1 | ||||
| Internal organs of chicken | 2 | ||||
| NAL-COL-AMP-STR-SXT | Retail chicken meat | aac(6′)-Iaa_1 blaTEM-1B_1 dfrA1_10 | 1 | 5 | 5 |
| NAL-COL-AMP-CHL-STR | Slaughterhouse | aac(6′)-Iaa_1 blaTEM-1B_1 aph(6)-Id_1 aph(3′)-Ib_5 tet(A)_6 aph(6)-Id_1 aph(3″)-Ib_5 sul2_2 | 1 | 5 | 5 |
| Retail chicken meat | 1 | ||||
| NAL-COL-AMP-TET-SXT | Truck | aac(6′)-Iaa_1 blaTEM-1B_1 aph(6)-Id_1 aph(3″)-Ib_5 tet(A)_6 aph(6)-Id_1 aph(3″)-Ib_5 sul2_2 | 1 | 5 | 5 |
| AMP-TET-CHL-STR-SXT | Environment | aac(6′)-Iaa_1 catA2_1 sul2_2 aph(3″)-Ib_5 aph(6)-Id_1 blaTEM-1B_1 | 1 | 5 | 5 |
| NAL-COL-AMP-SXT | Retail chicken meat | aac(6′)-Iaa_1 blaTEM-1B_1 dfrA1_10 | 2 | 4 | 4 |
| NAL-COL-AMP-STR | Retail chicken meat | sul2_2 aph(3″)-Ib_5 aph(6)-Id_1 blaTEM-1B_1 aac(6′)-Iaa_1 | 1 | 4 | 4 |
| NAL-AMP-STR | Retail chicken meat | sul2_2 aph(3″)-Ib_5 aph(6)-Id_1 blaTEM-1B_1 aac(6′)-Iaa_1 | 1 | 3 | 3 |
| NAL-COL-AMP | Internal organs of chicken | aac(6′)-Iaa_1 | 1 | 3 | 3 |
| NAL-AMP-SXT | Retail chicken meat | dfrA1_10 blaTEM-1B_1 aac(6′)-Iaa_1 | 1 | 3 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
La, T.-M.; Kim, T.; Lee, H.-J.; Lee, J.-B.; Park, S.-Y.; Choi, I.-S.; Lee, S.-W. Whole-Genome Analysis of Multidrug-Resistant Salmonella Enteritidis Strains Isolated from Poultry Sources in Korea. Pathogens 2021, 10, 1615. https://doi.org/10.3390/pathogens10121615
La T-M, Kim T, Lee H-J, Lee J-B, Park S-Y, Choi I-S, Lee S-W. Whole-Genome Analysis of Multidrug-Resistant Salmonella Enteritidis Strains Isolated from Poultry Sources in Korea. Pathogens. 2021; 10(12):1615. https://doi.org/10.3390/pathogens10121615
Chicago/Turabian StyleLa, Tae-Min, Taesoo Kim, Hong-Jae Lee, Joong-Bok Lee, Seung-Yong Park, In-Soo Choi, and Sang-Won Lee. 2021. "Whole-Genome Analysis of Multidrug-Resistant Salmonella Enteritidis Strains Isolated from Poultry Sources in Korea" Pathogens 10, no. 12: 1615. https://doi.org/10.3390/pathogens10121615