
Potential Risk Factors Associated with Infection with Bovine Leukaemia Virus in Dairy and Beef Cattle in Taiwan


Abstract
:1. Introduction
2. Results
2.1. Prevalence and Genotype of BLV Env Proviral DNA in Beef and Dairy Cows
2.2. Distributions of BLV Proviral DNA Prevalence among the Farm Features of Beef and Dairy Cows
2.3. Differences of BLV Expression among the Farm Features of Lactating Dairy Cows and Heifers
2.4. Prevalence of BLV Env Proviral DNA in the Breeding Bulls, Lactating Dairy Cows and Heifers from Farm 3
3. Discussion
4. Materials and Methods
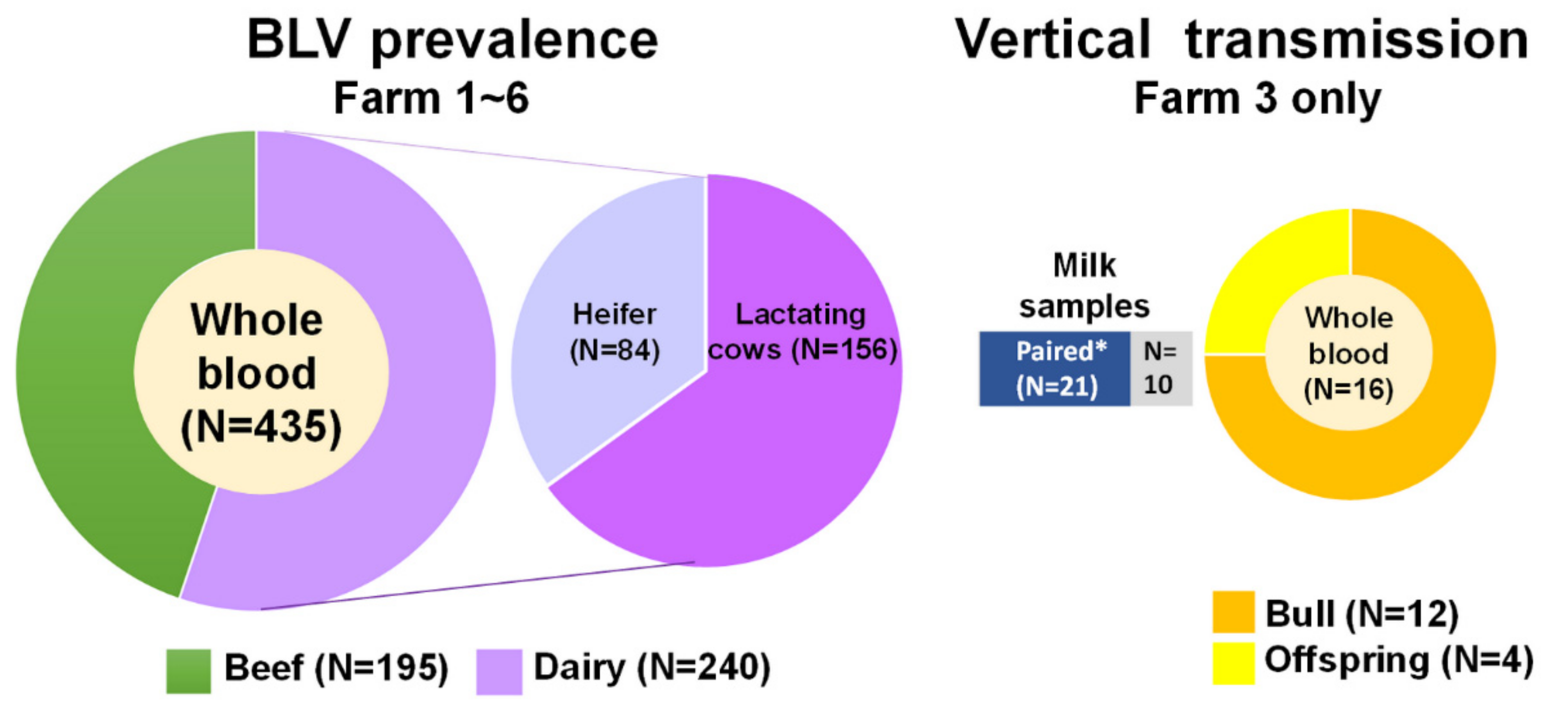
4.1. Sample Collection
4.2. DNA Extraction and Detection of BLV Proviral DNA by Nested PCR
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burny, A.; Cleuter, Y.; Kettmann, R.; Mammerickx, M.; Marbaix, G.; Portetelle, D.; van den Broeke, A.; Willems, L.; Thomas, R. Bovine leukaemia: Facts and hypotheses derived from the study of an infectious cancer. Vet. Microbiol. 1988, 17, 197–218. [Google Scholar] [CrossRef]
- Nekouei, O.; VanLeeuwen, J.; Stryhn, H.; Kelton, D.; Keefe, G. Lifetime effects of infection with bovine leukemia virus on longevity and milk production of dairy cows. Prev. Vet. Med. 2016, 133, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sargeant, J.M.; Kelton, D.F.; Martin, S.W.; Mann, E.D. Associations between farm management practices, productivity, and bovine leukemia virus infection in Ontario dairy herds. Prev. Vet. Med. 1997, 31, 211–221. [Google Scholar] [CrossRef]
- Bartlett, P.C.; Norby, B.; Byrem, T.M.; Parmelee, A.; Ledergerber, J.T.; Erskine, R.J. Bovine leukemia virus and cow longevity in Michigan dairy herds. J. Dairy Sci. 2013, 96, 1591–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benitez, O.J.; Norby, B.; Bartlett, P.C.; Maeroff, J.E.; Grooms, D.L. Impact of bovine leukemia virus infection on beef cow longevity. Prev. Vet. Med. 2020, 181, 105055. [Google Scholar] [CrossRef]
- Buehring, G.C.; Philpott, S.M.; Choi, K.Y. Humans have antibodies reactive with Bovine leukemia virus. AIDS Res. Hum. Retrovir. 2003, 19, 1105–1113. [Google Scholar] [CrossRef]
- Buehring, G.C.; Shen, H.M.; Jensen, H.M.; Choi, K.Y.; Sun, D.; Nuovo, G. Bovine leukemia virus DNA in human breast tissue. Emerg. Infect. Dis. 2014, 20, 772–782. [Google Scholar] [CrossRef] [Green Version]
- LaDronka, R.M.; Ainsworth, S.; Wilkins, M.J.; Norby, B.; Byrem, T.M.; Bartlett, P.C. Prevalence of Bovine Leukemia Virus Antibodies in US Dairy Cattle. Vet. Med. Int. 2018, 2018, 5831278. [Google Scholar] [CrossRef] [Green Version]
- VanLeeuwen, J.A.; Keefe, G.P.; Tremblay, R.; Power, C.; Wichtel, J.J. Seroprevalence of infection with Mycobacterium avium subspecies paratuberculosis, bovine leukemia virus, and bovine viral diarrhea virus in maritime Canada dairy cattle. Can. Vet. J. 2001, 42, 193–198. [Google Scholar]
- Murakami, K.; Kobayashi, S.; Konishi, M.; Kameyama, K.; Yamamoto, T.; Tsutsui, T. The recent prevalence of bovine leukemia virus (BLV) infection among Japanese cattle. Vet. Microbiol. 2011, 148, 84–88. [Google Scholar] [CrossRef]
- Yang, Y.; Fan, W.; Mao, Y.; Yang, Z.; Lu, G.; Zhang, R.; Zhang, H.; Szeto, C.; Wang, C. Bovine leukemia virus infection in cattle of China: Association with reduced milk production and increased somatic cell score. J. Dairy Sci. 2016, 99, 3688–3697. [Google Scholar] [CrossRef] [Green Version]
- Nekouei, O.; VanLeeuwen, J.; Sanchez, J.; Kelton, D.; Tiwari, A.; Keefe, G. Herd-level risk factors for infection with bovine leukemia virus in Canadian dairy herds. Prev. Vet. Med. 2015, 119, 105–113. [Google Scholar] [CrossRef]
- Meas, S.; Usui, T.; Ohashi, K.; Sugimoto, C.; Onuma, M. Vertical transmission of bovine leukemia virus and bovine immunodeficiency virus in dairy cattle herds. Vet. Microbiol. 2002, 84, 275–282. [Google Scholar] [CrossRef]
- Gillet, N.A.; Gutierrez, G.; Rodriguez, S.M.; de Brogniez, A.; Renotte, N.; Alvarez, I.; Trono, K.; Willems, L. Massive depletion of bovine leukemia virus proviral clones located in genomic transcriptionally active sites during primary infection. PLoS Pathog. 2013, 9, e1003687. [Google Scholar] [CrossRef]
- Gutierrez, G.; Alvarez, I.; Politzki, R.; Lomonaco, M.; Dus Santos, M.J.; Rondelli, F.; Fondevila, N.; Trono, K. Natural progression of Bovine Leukemia Virus infection in Argentinean dairy cattle. Vet. Microbiol. 2011, 151, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Monti, G.E.; Frankena, K.; De Jong, M.C. Evaluation of natural transmission of bovine leukaemia virus within dairy herds of Argentina. Epidemiol. Infect. 2007, 135, 228–237. [Google Scholar] [CrossRef]
- Kobayashi, S.; Tsutsui, T.; Yamamoto, T.; Hayama, Y.; Kameyama, K.; Konishi, M.; Murakami, K. Risk factors associated with within-herd transmission of bovine leukemia virus on dairy farms in Japan. BMC Vet. Res. 2010, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, J.C.; Li, C.Y.; Hsu, W.L.; Chuang, S.T. Molecular Epidemiological and Serological Studies of Bovine Leukemia Virus in Taiwan Dairy Cattle. Front. Vet. Sci. 2019, 6, 427. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.W.; Lv, W.F.; Cong, W.; Meng, Q.F.; Wang, C.F.; Shan, X.F.; Qian, A.D. Mycobacterium avium Subspecies paratuberculosis and Bovine Leukemia Virus Seroprevalence and Associated Risk Factors in Commercial Dairy and Beef Cattle in Northern and Northeastern China. Biomed. Res. Int. 2015, 2015, 315173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaworski, J.P.; Petersen, M.I.; Carignano, H.A.; Trono, K.G. Spontaneous virus reactivation in cattle chronically infected with bovine leukemia virus. BMC Vet. Res. 2019, 15, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, S.; Hidano, A.; Tsutsui, T.; Yamamoto, T.; Hayama, Y.; Nishida, T.; Muroga, N.; Konishi, M.; Kameyama, K.; Murakami, K. Analysis of risk factors associated with bovine leukemia virus seropositivity within dairy and beef breeding farms in Japan: A nationwide survey. Res. Vet. Sci. 2014, 96, 47–53. [Google Scholar] [CrossRef]
- Perez O’Brien, A.M.; Utsunomiya, Y.T.; Meszaros, G.; Bickhart, D.M.; Liu, G.E.; Van Tassell, C.P.; Sonstegard, T.S.; Da Silva, M.V.; Garcia, J.F.; Solkner, J. Assessing signatures of selection through variation in linkage disequilibrium between taurine and indicine cattle. Genet. Sel. Evol. 2014, 46, 19. [Google Scholar] [CrossRef] [Green Version]
- Panei, C.J.; Larsen, A.E.; Fuentealba, N.A.; Metz, G.E.; Echeverría, M.G.; Galosi, C.M.; Valera, A.R. Study of horn flies as vectors of bovine leukemia virus. Open Vet. J. 2019, 9, 33–37. [Google Scholar] [CrossRef] [Green Version]
- Foil, L.D.; Seger, C.L.; French, D.D.; Issel, C.J.; McManus, J.M.; Ohrberg, C.L.; Ramsey, R.T. Mechanical transmission of bovine leukemia virus by horse flies (Diptera: Tabanidae). J. Med. Entomol. 1988, 25, 374–376. [Google Scholar] [CrossRef]
- Hopkins, S.G.; DiGiacomo, R.F. Natural transmission of bovine leukemia virus in dairy and beef cattle. Vet. Clin. N. Am. Food Anim. Pract. 1997, 13, 107–128. [Google Scholar] [CrossRef]
- Ruiz, V.; Porta, N.G.; Lomonaco, M.; Trono, K.; Alvarez, I. Bovine Leukemia Virus Infection in Neonatal Calves. Risk Factors and Control Measures. Front. Vet. Sci. 2018, 5, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benitez, O.J.; Roberts, J.N.; Norby, B.; Bartlett, P.C.; Takeshima, S.N.; Watanuki, S.; Aida, Y.; Grooms, D.L. Breeding bulls as a potential source of bovine leukemia virus transmission in beef herds. J. Am. Vet. Med. Assoc. 2019, 254, 1335–1340. [Google Scholar] [CrossRef] [PubMed]
- Benitez, O.J.; Roberts, J.N.; Norby, B.; Bartlett, P.C.; Maeroff, J.E.; Grooms, D.L. Lack of Bovine leukemia virus transmission during natural breeding of cattle. Theriogenology 2019, 126, 187–190. [Google Scholar] [CrossRef]
- Chen, Y.C.; Chang, C.C.; Hsu, W.L.; Chuang, S.T. Dairy cattle with bovine leukaemia virus RNA show significantly increased leukocyte counts. Vet. J. 2020, 257, 105449. [Google Scholar] [CrossRef] [PubMed]
- Fechner, H.; Kurg, A.; Geue, L.; Blankenstein, P.; Mewes, G.; Ebner, D.; Beier, D. Evaluation of polymerase chain reaction (PCR) application in diagnosis of bovine leukaemia virus (BLV) infection in naturally infected cattle. Zent. Vet. B 1996, 43, 621–630. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Factors | BLV Detection | ||
|---|---|---|---|
| Positive (n = 125) | Negative (n = 310) | p-Value | |
| Purpose of production | <0.001 § | ||
| Beef | 23 | 172 | |
| Dairy | 102 | 138 | |
| Age (months) | 34.38 ± 1.90 † | 20.78 ± 1.13 † | <0.001 ‡ |
| Breed | 0.001 § | ||
| Holstein | 118 | 249 | |
| Angus | 6 | 55 | |
| Mixed | 1 | 6 | |
| Gender | <0.001 § | ||
| Male | 13 | 97 | |
| Female | 109 | 199 | |
| Castrated male | 3 | 14 | |
| Herd size (head) | 0.015 § | ||
| <50 | 19 | 47 | |
| 50–200 | 5 | 45 | |
| 201–400 | 97 | 205 | |
| >400 | 4 | 13 | |
| Management pattern | 0.004 ¶ | ||
| Intensive | 123 | 281 | |
| Pasture | 2 | 29 | |
| Farm owner | 0.015 § | ||
| 1 | 4 | 13 | |
| 2 | 2 | 29 | |
| 3 | 34 | 70 | |
| 4 | 27 | 89 | |
| 5 | 18 | 42 | |
| 6 | 40 | 67 | |
| Artificial insemination | <0.001 § | ||
| No | 20 | 139 | |
| No (Bull only) | 28 | 58 | |
| Yes | 63 | 100 | |
| Yes (Combined) | 14 | 13 | |
| Regular rectal palpation | <0.001 § | ||
| No | 29 | 196 | |
| Yes | 96 | 114 | |
| Sterilized colostrum | 0.008 § | ||
| No | 6 | 42 | |
| Yes | 119 | 268 | |
| Vaccination | 0.865 § | ||
| No | 17 | 43 | |
| Yes (Needle sharing) | 66 | 171 | |
| Yes (One shot per needle) | 42 | 96 | |
| Factors | BLV in Beef Cattle | BLV in Dairy Cows | ||||
|---|---|---|---|---|---|---|
| Positive (n = 23) | Negative (n = 172) | p-Value | Positive (n = 102) | Negative (n = 138) | p-Value | |
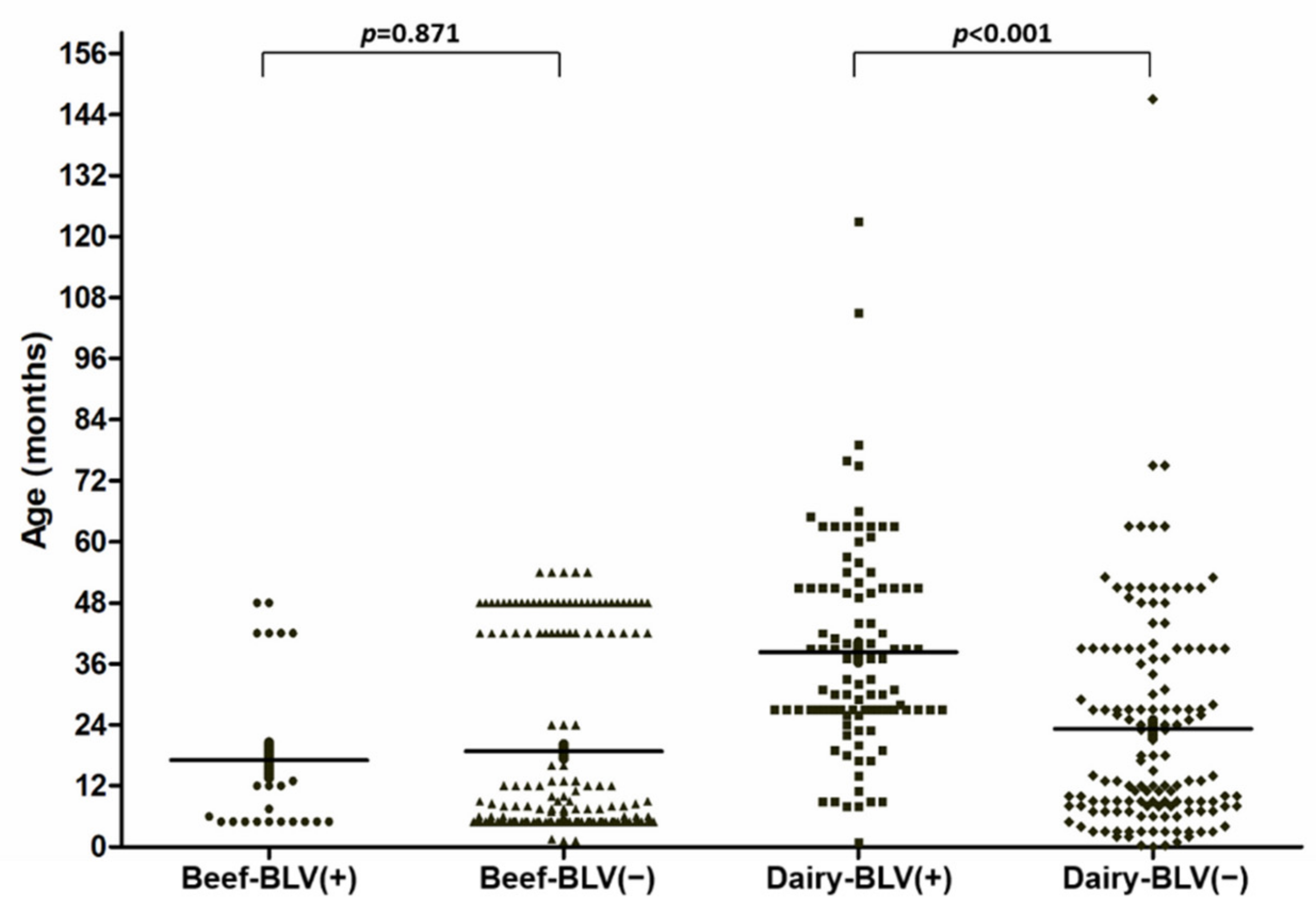
| Age (months) | 17.07 ± 3.49 † | 19.08 ± 1.41 † | 0.871 ‡ | 38.28 ± 1.99 † | 23.20 ± 1.80 † | <0.001 ‡ |
| Breed | 0.841 § | NA | ||||
| Holstein | 16 | 111 | 102 | 138 | ||
| Angus | 6 | 55 | 0 | 0 | ||
| Mixed | 1 | 6 | 0 | 0 | ||
| Gender | 0.702 § | NA | ||||
| Male | 13 | 97 | 0 | 0 | ||
| Female | 7 | 61 | 102 | 138 | ||
| Castrated Male | 3 | 14 | 0 | 0 | ||
| Herd size | 0.276 § | 0.003 § | ||||
| <50 | 4 | 13 | 0 | 0 | ||
| 50–200 | 2 | 35 | 17 | 12 | ||
| 201–400 | 3 | 27 | 2 | 18 | ||
| >400 | 14 | 97 | 83 | 108 | ||
| Management pattern | 0.542 ¶ | NA | ||||
| Intensive | 21 | 143 | 102 | 138 | ||
| Pasture | 2 | 29 | 0 | 0 | ||
| Farm owner | 0.514 § | 0.216 § | ||||
| 1 | 4 | 13 | 0 | 0 | ||
| 2 | 2 | 29 | 0 | 0 | ||
| 3 | 3 | 27 | 31 | 43 | ||
| 4 | 10 | 77 | 17 | 12 | ||
| 5 | 4 | 26 | 14 | 16 | ||
| 6 | 0 | 0 | 40 | 67 | ||
| Artificial insemination | 0.522 § | <0.001 § | ||||
| No | 17 | 111 | 3 | 28 | ||
| No (Bull only) | 6 | 55 | 22 | 3 | ||
| Yes | 0 | 6 | 63 | 94 | ||
| Yes (Combined) | 0 | 0 | 14 | 13 | ||
| Regular rectal palpation | 0.260 ¶ | <0.001 § | ||||
| No | 21 | 137 | 8 | 59 | ||
| Yes | 2 | 35 | 94 | 79 | ||
| Sterilized colostrum | 0.862 § | NA | ||||
| No | 6 | 42 | 0 | 0 | ||
| Yes | 17 | 130 | 102 | 138 | ||
| Vaccination | 0.525 § | 0.355 § | ||||
| No | 3 | 27 | 14 | 16 | ||
| Yes (Needle sharing) | 18 | 116 | 48 | 55 | ||
| Yes (One shot per needle) | 2 | 29 | 40 | 67 | ||
| Factors | Age (months) | ||
|---|---|---|---|
| ≤15.5 (n = 79) | >15.5 (n = 161) | p-Value | |
| Type | <0.001 ¶ | ||
| Lactating | 1 | 155 | |
| Heifer | 78 | 6 | |
| BLV | <0.001 § | ||
| Negative | 70 | 68 | |
| Positive | 9 | 93 | |
| Herd size | <0.001 § | ||
| 50–200 | 0 | 29 | |
| 201–400 | 20 | 0 | |
| >400 | 59 | 132 | |
| Farm owner | <0.001 § | ||
| 3 | 39 | 35 | |
| 4 | 0 | 29 | |
| 5 | 0 | 30 | |
| 6 | 40 | 107 | |
| Artificial insemination | <0.001 § | ||
| No | 31 | 0 | |
| No (Bull only) | 0 | 25 | |
| Yes | 47 | 110 | |
| Yes (Combined) | 1 | 26 | |
| Regular rectal palpation | <0.001 § | ||
| No | 67 | 0 | |
| Yes | 12 | 161 | |
| Vaccination | <0.001 § | ||
| No | 0 | 30 | |
| Yes (Needle sharing) | 39 | 64 | |
| Yes (One shot per needle) | 40 | 67 | |
| Factors | BLV in Lactating Dairy Cows | BLV in Heifers | ||||
|---|---|---|---|---|---|---|
| Positive (n = 90) | Negative (n = 66) | p-Value | Positive (n = 12) | Negative (n = 72) | p-Value | |
| Age (months) | 41.94 ± 1.94 † | 39.98 ± 2.36 † | 0.336 ‡ | 10.83 ± 1.41 † | 7.81 ± 0.51 † | 0.049 ‡ |
| Herd size | 0.911 § | 0.722 ¶ | ||||
| 50–200 | 17 | 12 | 2 | 18 | ||
| >400 | 73 | 54 | 10 | 54 | ||
| Farm owner | 0.038 § | 0.656 § | ||||
| 3 | 24 | 6 | 7 | 37 | ||
| 4 | 17 | 12 | 0 | 0 | ||
| 5 | 14 | 16 | 0 | 0 | ||
| 6 | 35 | 32 | 5 | 35 | ||
| Artificial insemination | 0.004 § | 0.521 ¶ | ||||
| No | 0 | 0 | 3 | 28 | ||
| No (Bull only) | 22 | 3 | 0 | 0 | ||
| Yes | 54 | 50 | 9 | 44 | ||
| Combined | 14 | 13 | 0 | 0 | ||
| Regular rectal palpation | NA | 0.251 ¶ | ||||
| No | 0 | 0 | 8 | 59 | ||
| Yes | 90 | 66 | 4 | 13 | ||
| Vaccination | 0.059 § | 0.656 § | ||||
| No | 14 | 16 | 0 | 0 | ||
| Yes (Needle sharing) | 41 | 18 | 7 | 37 | ||
| Yes (One shot per needle) | 35 | 32 | 5 | 35 | ||
| Factors | Farm Owner | |||||
|---|---|---|---|---|---|---|
| 1 (n = 17) | 2 (n = 31) | 3 (n = 120) | 4 (n = 116) | 5 (n = 60) | 6 (n = 107) | |
| Production usage | Beef | Beef | Mixed | Mixed | Mixed | Dairy |
| Types | ||||||
| Beef | 17 | 31 | 30 | 87 | 30 | 0 |
| Dairy | 0 | 0 | 30 | 29 | 30 | 67 |
| Heifer | 0 | 0 | 44 | 0 | 0 | 40 |
| Offspring | 0 | 0 | 4 | 0 | 0 | 0 |
| Breeding bull | 0 | 0 | 12 | 0 | 0 | 0 |
| Age (months) | 10.15 ± 0.55 † | 48.00 ± 0.00 † | 18.87 ± 1.55 † | 13.59 ± 1.50 † | 46.80 ± 2.63 † | 25.89 ± 1.83 † |
| Breed | ||||||
| Holstein | 16 | 0 | 120 | 110 | 30 | 107 |
| Angus | 0 | 31 | 0 | 0 | 30 | 0 |
| Mixed | 1 | 0 | 0 | 6 | 0 | 0 |
| Gender | ||||||
| Male | 0 | 0 | 41 | 81 | 0 | 0 |
| Female | 1 | 31 | 74 | 35 | 60 | 107 |
| Castrated male | 16 | 0 | 5 | 0 | 0 | 0 |
| Herd size | <50 | 50–200 | >400 | 50–200 | >400 | >400 |
| Management pattern | Intensive | Pasture | Intensive | Intensive | Intensive | Intensive |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-C.; Chin, W.-Y.; Chang, C.-C.; Chuang, S.-T.; Hsu, W.-L. Potential Risk Factors Associated with Infection with Bovine Leukaemia Virus in Dairy and Beef Cattle in Taiwan. Pathogens 2021, 10, 1553. https://doi.org/10.3390/pathogens10121553
Chen Y-C, Chin W-Y, Chang C-C, Chuang S-T, Hsu W-L. Potential Risk Factors Associated with Infection with Bovine Leukaemia Virus in Dairy and Beef Cattle in Taiwan. Pathogens. 2021; 10(12):1553. https://doi.org/10.3390/pathogens10121553
Chicago/Turabian StyleChen, Yi-Chen, Wen-Yu Chin, Chao-Chin Chang, Shih-Te Chuang, and Wei-Li Hsu. 2021. "Potential Risk Factors Associated with Infection with Bovine Leukaemia Virus in Dairy and Beef Cattle in Taiwan" Pathogens 10, no. 12: 1553. https://doi.org/10.3390/pathogens10121553