
Pb103 Regulates Zygote/Ookinete Development in Plasmodium berghei via Double Zinc Finger Domains

Abstract
:1. Introduction
2. Results and Discussion
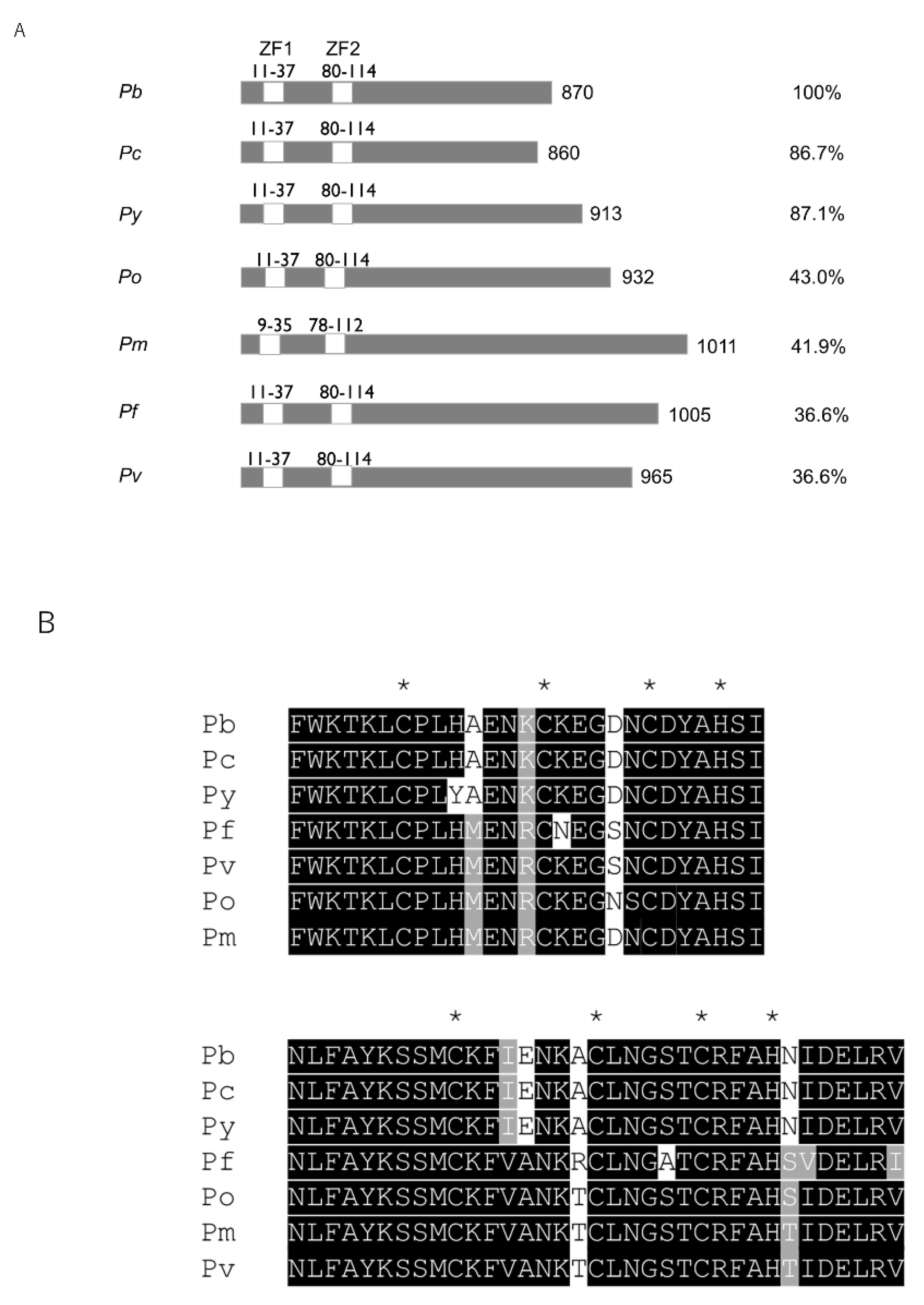
2.1. Pb103 Is Conserved in Plasmodium
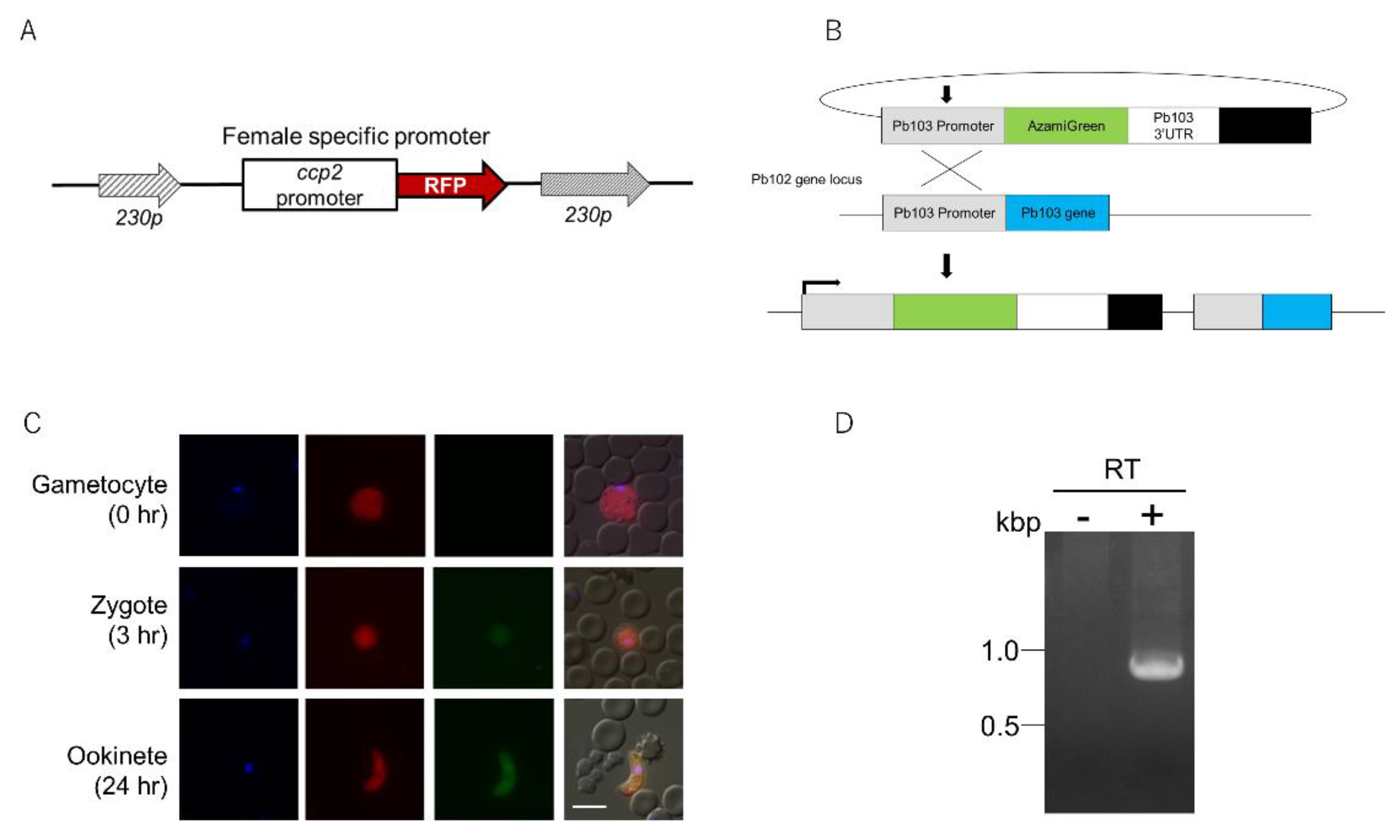
2.2. Expression Analysis of Pb103 by Using the Pb103-AGFP Reporter
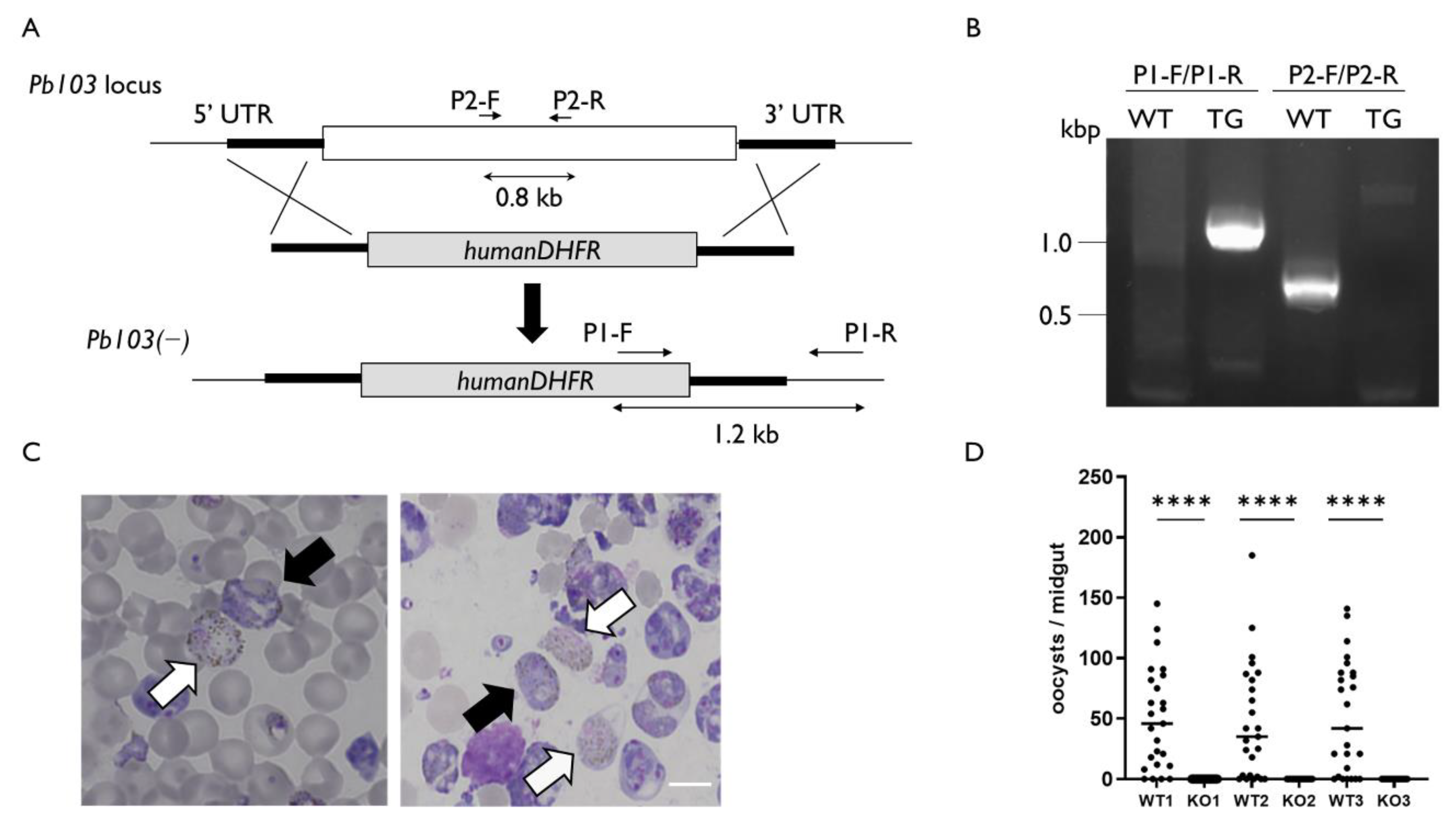
2.3. Pb103 Is Essential for Parasite Reproduction
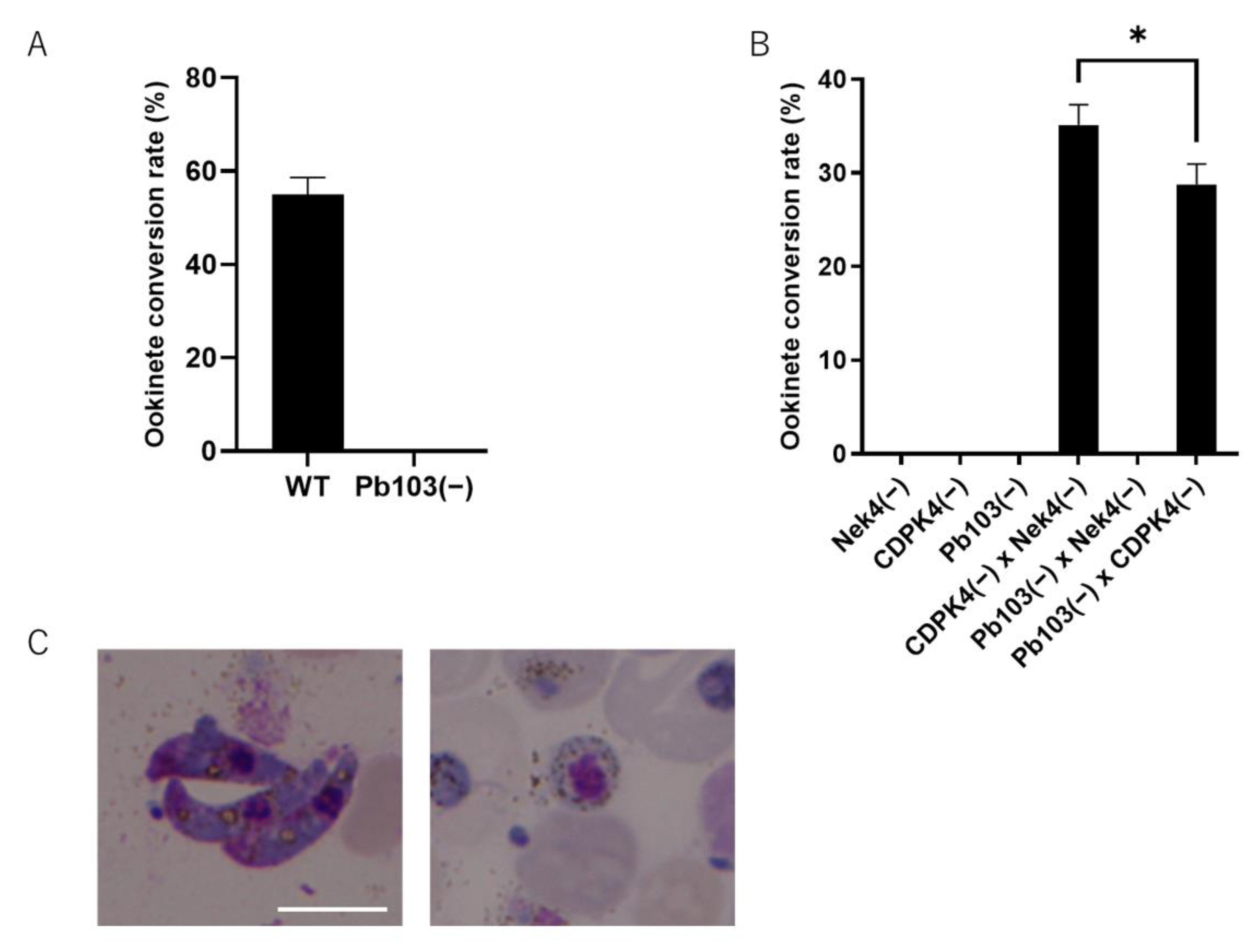
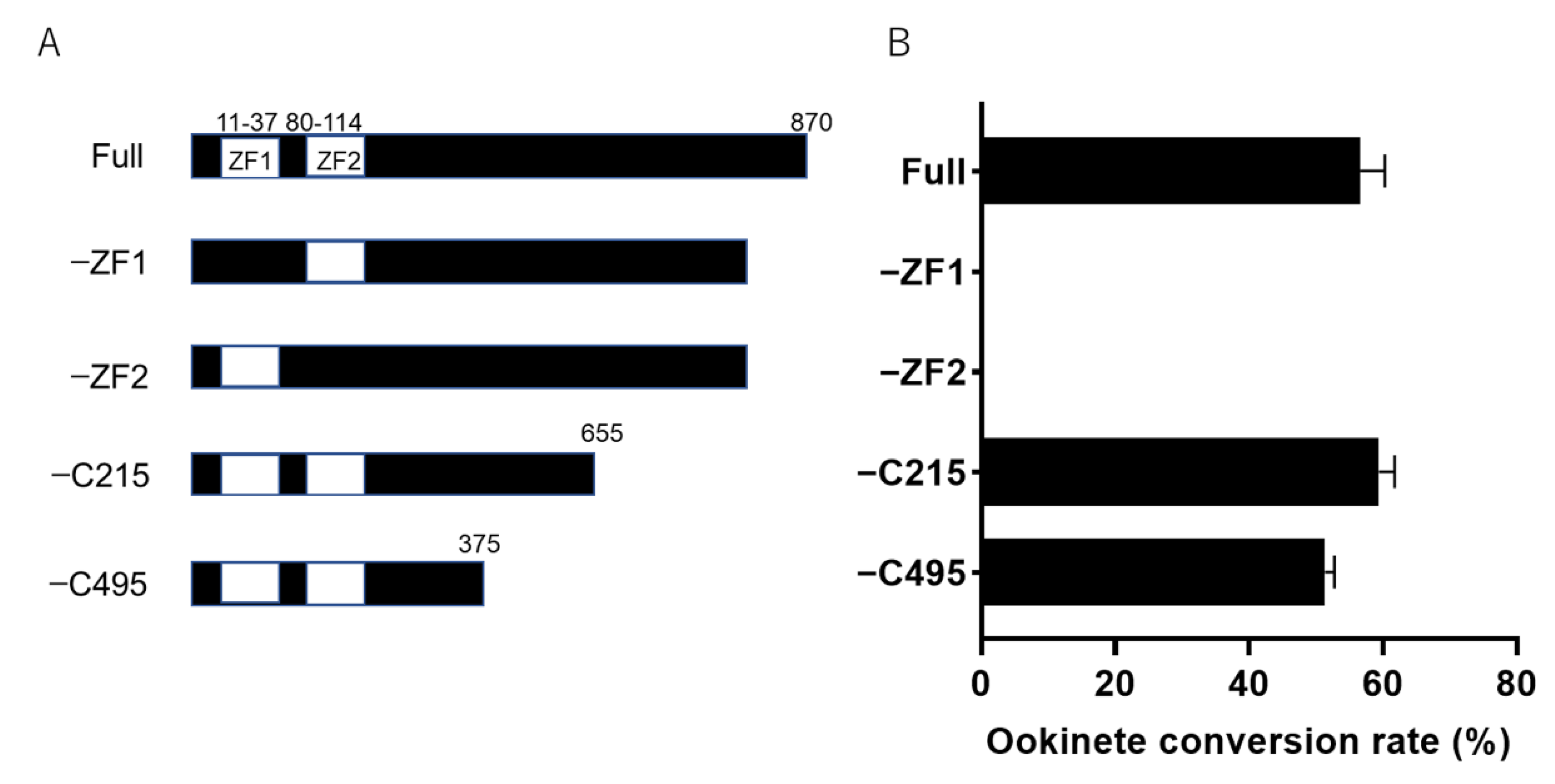
2.4. Zinc Finger Domains Are Indispensable for Ookinete Maturation
2.5. Mechanisms of Post-Translation Repression of Pb103
3. Methods
3.1. Ethics Statement
3.2. Animals and Parasites
3.3. Generation and Expression Analysis of the Pb103-AGFP Reporter
3.4. Reverse Transcription PCR (RT-PCR)
3.5. Targeting Disruption of Pb103, PbCDPK4, and PbNek-4 Genes
3.6. Development of Pb03(−) in Mosquitoes and Transmission to Mice
3.7. In Vitro Cross-Fertilization Assay
3.8. Generation of Partially Truncated Pb103 Mutants
3.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. 2020 World Malaria Report; WHO Press: Geneva, Switzerland, 2020. [Google Scholar]
- Kubo-Irie, M.; Hirai, M.; Irie, M.; Mohri, H. Postulated Process of Axoneme Organization in the Male Gametogenesis of Malaria Parasite Plasmodium berghei. Zool. Sci. 2020, 38, 187–192. [Google Scholar] [CrossRef]
- Billker, O.; Shaw, M.K.; Margos, G.; Sinden, R.E. The Roles of Temperature, PH and Mosquito Factors as Triggers of Male and Female Gametogenesis of Plasmodium berghei in Vitro. Parasitology 1997, 115, 1–7. [Google Scholar] [CrossRef]
- Hirai, M.; Arai, M.; Mori, T.; Miyagishima, S.; Kawai, S.; Kita, K.; Kuroiwa, T.; Terenius, O.; Matsuoka, H. Male Fertility of Malaria Parasites Is Determined by GCS1, a Plant-Type Reproduction Factor. Curr. Biol. 2008, 18, 607–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Tewari, R.; Ning, J.; Blagborough, A.M.; Garbom, S.; Pei, J.; Grishin, N.V.; Steele, R.E.; Sinden, R.E.; Snell, W.J.; et al. The Conserved Plant Sterility Gene HAP2 Functions after Attachment of Fusogenic Membranes in Chlamydomonas and Plasmodium Gametes. Gene Dev. 2008, 22, 1051–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dijk, M.R.; Janse, C.J.; Thompson, J.; Waters, A.P.; Braks, J.A.M.; Dodemont, H.J.; Stunnenberg, H.G.; van Gemert, G.-J.; Sauerwein, R.W.; Eling, W. A Central Role for P48/45 in Malaria Parasite Male Gamete Fertility. Cell 2001, 104, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, J.L.; Gómez-Díaz, E. The Second Life of Plasmodium in the Mosquito Host: Gene Regulation on the Move. Brief. Funct. Genom. 2019, 18, 313–357. [Google Scholar] [CrossRef] [PubMed]
- Coelho, C.H.; Rappuoli, R.; Hotez, P.J.; Duffy, P.E. Transmission-Blocking Vaccines for Malaria: Time to Talk about Vaccine Introduction. Trends Parasitol. 2019, 35, 483–486. [Google Scholar] [CrossRef]
- Hirai, M.; Mori, T. Fertilization Is a Novel Attacking Site for the Transmission Blocking of Malaria Parasites. Acta Trop. 2010, 114, 157–161. [Google Scholar] [CrossRef]
- Painter, H.J.; Campbell, T.L.; Llinás, M. The Apicomplexan AP2 Family: Integral Factors Regulating Plasmodium Development. Mol. Biochem. Parasit. 2011, 176, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sinha, A.; Hughes, K.R.; Modrzynska, K.K.; Otto, T.D.; Pfander, C.; Dickens, N.J.; Religa, A.A.; Bushell, E.; Graham, A.L.; Cameron, R.; et al. A Cascade of DNA-Binding Proteins for Sexual Commitment and Development in Plasmodium. Nature 2014, 507, 253–257. [Google Scholar] [CrossRef]
- Kafsack, B.F.; Rovira-Graells, N.; Clark, T.G.; Bancells, C.; Crowley, V.M.; Campino, S.G.; Williams, A.E.; Drought, L.G.; Kwiatkowski, D.P.; Baker, D.A.; et al. A Transcriptional Switch Underlies Commitment to Sexual Development in Malaria Parasites. Nature 2014, 507, 248–252. [Google Scholar] [CrossRef] [Green Version]
- Yuda, M.; Iwanaga, S.; Kaneko, I.; Kato, T. Global Transcriptional Repression: An Initial and Essential Step for Plasmodium Sexual Development. Proc. Natl. Acad. Sci. USA 2015, 112, 12824–12829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuda, M.; Kaneko, I.; Iwanaga, S.; Murata, Y.; Kato, T. Female-specific Gene Regulation in Malaria Parasites by an AP2-family Transcription Factor. Mol. Microbiol. 2020, 113, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, I.; Iwanaga, S.; Kato, T.; Kobayashi, I.; Yuda, M. Genome-Wide Identification of the Target Genes of AP2-O, a Plasmodium AP2-Family Transcription Factor. PLoS Pathog. 2015, 11, e1004905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modrzynska, K.; Pfander, C.; Chappell, L.; Yu, L.; Suarez, C.; Dundas, K.; Gomes, A.; Goulding, D.; Rayner, J.C.; Choudhary, J.; et al. A Knockout Screen of ApiAP2 Genes Reveals Networks of Interacting Transcriptional Regulators Controlling the Plasmodium Life Cycle. Cell Host Microbe 2017, 21, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lasonder, E.; Rijpma, S.R.; van Schaijk, B.C.L.; Hoeijmakers, W.A.M.; Kensche, P.R.; Gresnigt, M.S.; Italiaander, A.; Vos, M.W.; Woestenenk, R.; Bousema, T.; et al. Integrated Transcriptomic and Proteomic Analyses of P. Falciparum Gametocytes: Molecular Insight into Sex-Specific Processes and Translational Repression. Nucleic Acids Res. 2016, 44, 6087–6101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mair, G.R.; Braks, J.A.M.; Garver, L.S.; Wiegant, J.C.A.G.; Hall, N.; Dirks, R.W.; Khan, S.M.; Dimopoulos, G.; Janse, C.J.; Waters, A.P. Regulation of Sexual Development of Plasmodium by Translational Repression. Science 2006, 313, 667–669. [Google Scholar] [CrossRef] [Green Version]
- Mair, G.R.; Lasonder, E.; Garver, L.S.; Franke-Fayard, B.; Carret, C.K.; Wiegant, J.; Dirks, R.W.; Dimopoulos, G.; Janse, C.J.; Waters, A.P. Universal Features of Post-Transcriptional Gene Regulation Are Critical for Plasmodium Zygote Development. PLoS Pathog. 2010, 6, e1000767. [Google Scholar] [CrossRef] [Green Version]
- Nguyen-Chi, M.; Morello, D. RNA-Binding Proteins, RNA Granules, and Gametes: Is Unity Strength? Reproduction 2011, 142, 803–817. [Google Scholar] [CrossRef] [Green Version]
- Beckmann, B.M.; Horos, R.; Fischer, B.; Castello, A.; Eichelbaum, K.; Alleaume, A.-M.; Schwarzl, T.; Curk, T.; Foehr, S.; Huber, W.; et al. The RNA-Binding Proteomes from Yeast to Man Harbour Conserved EnigmRBPs. Nat. Commun. 2015, 6, 10127. [Google Scholar] [CrossRef]
- Reddy, B.N.; Shrestha, S.; Hart, K.J.; Liang, X.; Kemirembe, K.; Cui, L.; Lindner, S.E. A Bioinformatic Survey of RNA-Binding Proteins in Plasmodium. BMC Genom. 2015, 16, 890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngwa, C.J.; Farrukh, A.; Pradel, G. Zinc Finger Proteins of Plasmodium Falciparum. Cell Microbiol. 2021, e13387. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Fan, Q.; Parker, D.; Li, X.; Li, J.; Cui, L. Puf Mediates Translation Repression of Transmission-Blocking Vaccine Candidates in Malaria Parasites. PLoS Pathog. 2013, 9, e1003268. [Google Scholar] [CrossRef] [Green Version]
- Chêne, A.; Vembar, S.S.; Rivière, L.; Lopez-Rubio, J.J.; Claes, A.; Siegel, T.N.; Sakamoto, H.; Scheidig-Benatar, C.; Hernandez-Rivas, R.; Scherf, A. PfAlbas Constitute a New Eukaryotic DNA/RNA-Binding Protein Family in Malaria Parasites. Nucleic Acids Res. 2012, 40, 3066–3077. [Google Scholar] [CrossRef] [PubMed]
- Komaki-Yasuda, K.; Okuwaki, M.; Nagata, K.; Kawazu, S.; Kano, S. Identification of a Novel and Unique Transcription Factor in the Intraerythrocytic Stage of Plasmodium Falciparum. PLoS ONE 2013, 8, e74701. [Google Scholar] [CrossRef]
- Müller, K.; Silvie, O.; Mollenkopf, H.-J.; Matuschewski, K. Pleiotropic Roles for the Plasmodium berghei RNA Binding Protein UIS12 in Transmission and Oocyst Maturation. Front. Cell Infect. Microbiol. 2021, 11, 624945. [Google Scholar] [CrossRef] [PubMed]
- Yeoh, L.M.; Goodman, C.D.; Mollard, V.; McFadden, G.I.; Ralph, S.A. Comparative Transcriptomics of Female and Male Gametocytes in Plasmodium berghei and the Evolution of Sex in Alveolates. BMC Genom. 2017, 18, 734. [Google Scholar] [CrossRef] [Green Version]
- Ponzi, M.; Sidén-Kiamos, I.; Bertuccini, L.; Currà, C.; Kroeze, H.; Camarda, G.; Pace, T.; Franke-Fayard, B.; Laurentino, E.C.; Louis, C.; et al. Egress of Plasmodium berghei Gametes from Their Host Erythrocyte Is Mediated by the MDV-1/PEG3 Protein. Cell Microbiol. 2009, 11, 1272–1288. [Google Scholar] [CrossRef]
- Rosales-Ronquillo, M.C.; Silverman, P.H. In Vitro Ookinete Development of the Rodent Malarial Parasite, Plasmodium berghei. J. Parasitol. 1974, 60, 819–824. [Google Scholar] [CrossRef]
- Braks, J.A.M.; Mair, G.R.; Franke-Fayard, B.; Janse, C.J.; Waters, A.P. A Conserved U-Rich RNA Region Implicated in Regulation of Translation in Plasmodium Female Gametocytes. Nucleic Acids Res. 2008, 36, 1176–1186. [Google Scholar] [CrossRef] [Green Version]
- Guerreiro, A.; Deligianni, E.; Santos, J.M.; Silva, P.; Louis, C.; Pain, A.; Janse, C.J.; Franke-Fayard, B.; Carret, C.K.; Siden-Kiamos, I.; et al. Genome-Wide RIP-Chip Analysis of Translational Repressor-Bound MRNAs in the Plasmodium Gametocyte. Genome Biol. 2014, 15, 493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebastian, S.; Brochet, M.; Collins, M.O.; Schwach, F.; Jones, M.L.; Goulding, D.; Rayner, J.C.; Choudhary, J.S.; Billker, O. A Plasmodium Calcium-Dependent Protein Kinase Controls Zygote Development and Transmission by Translationally Activating Repressed MRNAs. Cell Host Microbe. 2012, 12, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janse, C.J.; Ramesar, J.; Waters, A.P. High-Efficiency Transfection and Drug Selection of Genetically Transformed Blood Stages of the Rodent Malaria Parasite Plasmodium berghei. Nat. Protoc. 2006, 1, 346–356. [Google Scholar] [CrossRef]
- Braks, J.A.M.; Franke-Fayard, B.; Kroeze, H.; Janse, C.J.; Waters, A.P. Development and Application of a Positive–Negative Selectable Marker System for Use in Reverse Genetics in Plasmodium. Nucleic Acids Res. 2006, 34, e39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwayama, H.; Obara, S.; Morio, T.; Katoh, M.; Urushihara, H.; Tanaka, Y. PCR-Mediated Generation of a Gene Disruption Construct without the Use of DNA Ligase and Plasmid Vectors. Nucleic Acids Res. 2002, 30, e2. [Google Scholar] [CrossRef] [PubMed]
- Billker, O.; Dechamps, S.; Tewari, R.; Wenig, G.; Franke-Fayard, B.; Brinkmann, V. Calcium and a Calcium-Dependent Protein Kinase Regulate Gamete Formation and Mosquito Transmission in a Malaria Parasite. Cell 2004, 117, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Reininger, L.; Billker, O.; Tewari, R.; Mukhopadhyay, A.; Fennell, C.; Dorin-Semblat, D.; Doerig, C.; Goldring, D.; Harmse, L.; Ranford-Cartwright, L.; et al. A NIMA-Related Protein Kinase Is Essential for Completion of the Sexual Cycle of Malaria Parasites*. J. Biol. Chem. 2005, 280, 31957–31964. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, M.; Hirai, M.; Tachibana, S.-I.; Mori, T.; Mita, T. Fitness of Sulfadoxine-Resistant Plasmodium berghei Harboring a Single Mutation in Dihydropteroate Synthase (Dhps). Acta Trop. 2021, 222, 106049. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gametocytemia (%) | Sex Ratio (♂:♀) | |
|---|---|---|
| Pb103(−) | 0.24 | 1:2.38 |
| WT | 0.25 | 1:1.96 |
| Parasites | No. of Salivary Glands | Infected/Injected |
|---|---|---|
| WT | 5 | 1/1 |
| 6 | 1/1 | |
| 4 | 1/1 | |
| Pb103(−) | 15 | 0/1 |
| 20 | 0/1 | |
| 15 | 0/1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirai, M.; Maeta, A.; Mori, T.; Mita, T. Pb103 Regulates Zygote/Ookinete Development in Plasmodium berghei via Double Zinc Finger Domains. Pathogens 2021, 10, 1536. https://doi.org/10.3390/pathogens10121536
Hirai M, Maeta A, Mori T, Mita T. Pb103 Regulates Zygote/Ookinete Development in Plasmodium berghei via Double Zinc Finger Domains. Pathogens. 2021; 10(12):1536. https://doi.org/10.3390/pathogens10121536
Chicago/Turabian StyleHirai, Makoto, Akimasa Maeta, Toshiyuki Mori, and Toshihiro Mita. 2021. "Pb103 Regulates Zygote/Ookinete Development in Plasmodium berghei via Double Zinc Finger Domains" Pathogens 10, no. 12: 1536. https://doi.org/10.3390/pathogens10121536