
Molecular Typing and Antimicrobial Susceptibility Profiles of Streptococcus uberis Isolated from Sheep Milk


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
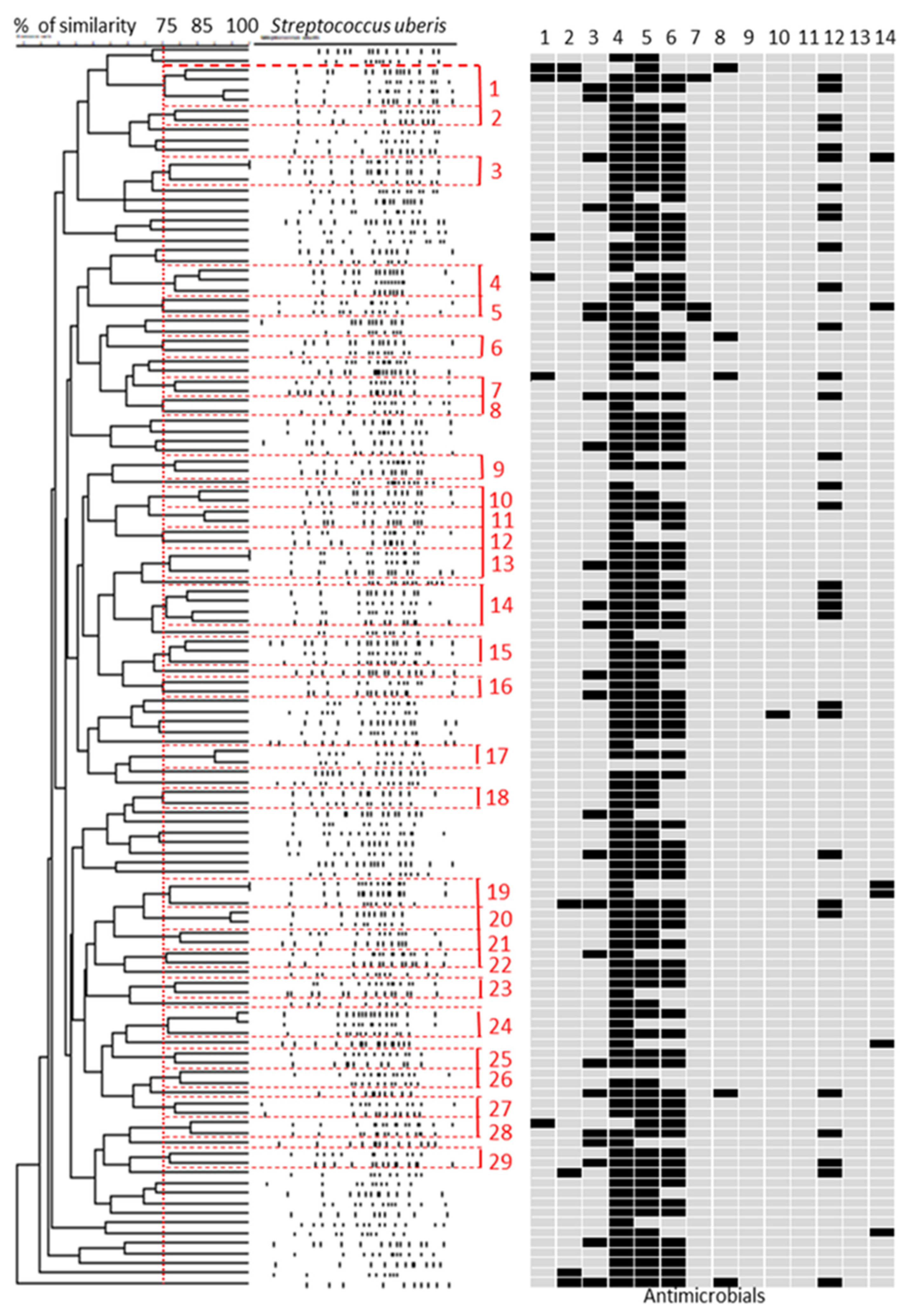
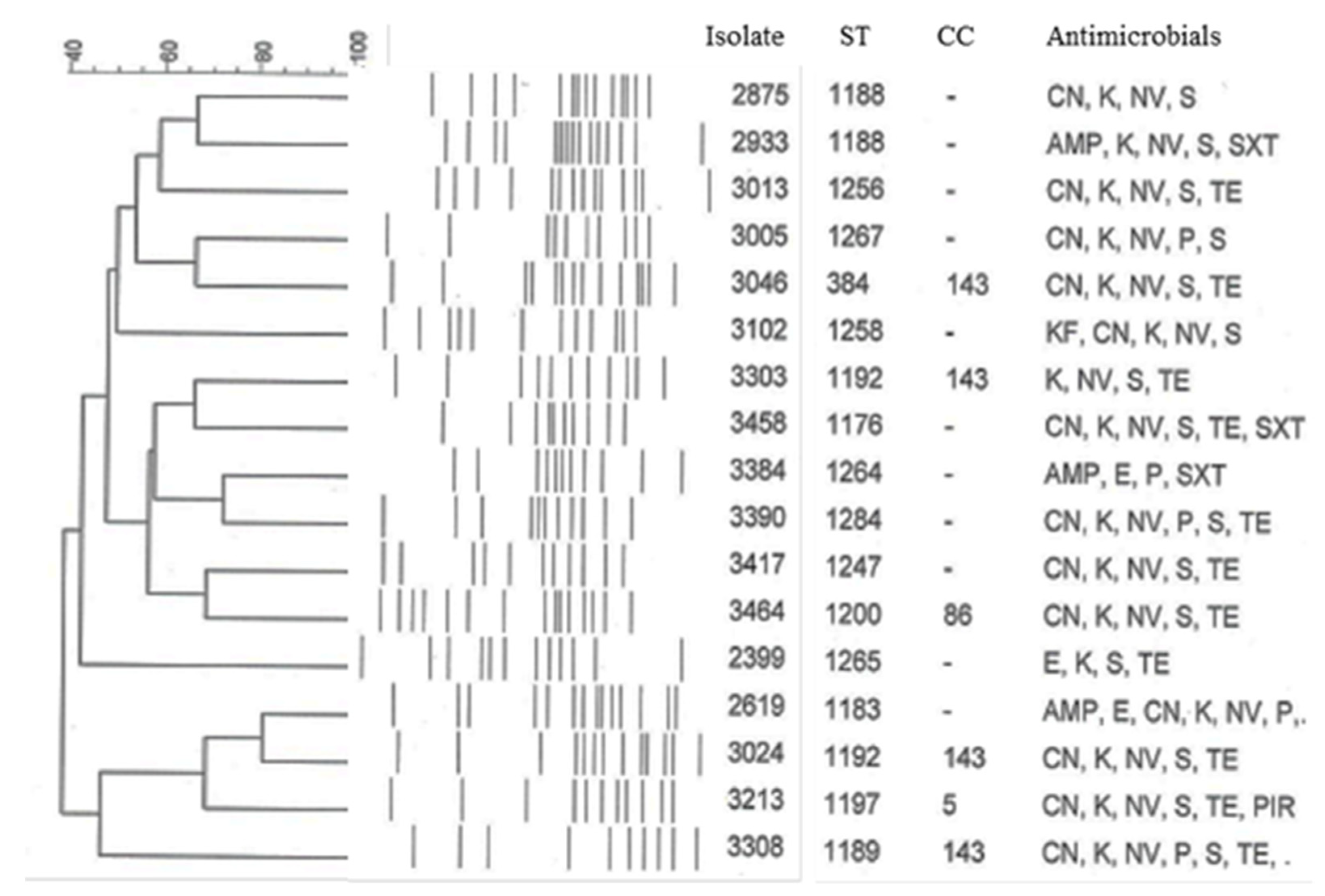
2.1. Streptococcus uberis Genotyping
2.2. Antimicrobial Susceptibility and Resistance Genes
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates and DNA Extraction
4.2. MLST
4.3. PFGE
4.4. Antimicrobial Susceptibility Testing
4.5. Detection of Resistance Genes by PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marogna, G.; Rolesu, S.; Lollai, S.; Tola, S.; Leori, S.G. Clinical findings in sheep farms affected by recurrent bacterial mastitis. Small Rumin. Res. 2010, 88, 119–125. [Google Scholar] [CrossRef]
- Marogna, G.; Pilo, C.; Vidili, A.; Tola, S.; Schianchi, G.; Leori, S.G. Comparison of clinical findings, microbiological results, and farming parameters in goat herds affected by recurrent infectious mastitis. Small Rumin. Res. 2011, 102, 74–83. [Google Scholar] [CrossRef]
- Gelasakis, A.I.; Mavrogianni, V.S.; Petridis, I.G.; Vasileiou, N.G.C.; Fthenakis, G.C. Mastitis in sheep: The last 10 years and the future of research. Vet. Microbiol. 2015, 181, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Dore, S.; Liciardi, M.; Amatiste, S.; Bergagna, S.; Bolzoni, G.; Caligiuri, V.; Cerrone, A.; Farina, G.; Montagna, C.O.; Saletti, M.A.; et al. Survey on small ruminant bacterial mastitis in Italy, 2013-2014. Small Rumin. Res. 2016, 141, 91–93. [Google Scholar] [CrossRef] [Green Version]
- Rosa, M.N.; Agnoletti, F.; Lollai, S.; Tola, S. Comparison of PCR-RFLP, API® 20 Strep and MALDI-TOF MS for identification of Streptococcus spp. collected from sheep and goat milk samples. Small Rumin. Res. 2019, 180, 35–40. [Google Scholar] [CrossRef]
- Gilchrist, T.L.; Smith, D.G.E.; Fitzpatrick, J.L.; Zadoks, R.N.; Fontaine, M.C. Comparative molecular analysis of ovine and bovine Streptococcus uberis isolates. J. Dairy Sci. 2013, 96, 962–970. [Google Scholar] [CrossRef]
- Käppeli, N.; Morach, M.; Zurfluh, K.; Corti, S.; Nüesch-Inderbinen, M.; Stephan, R. Sequence types and antimicrobial resistance profiles of Streptococcus uberis isolated from bovine mastitis. Front. Vet. Sci. 2019, 6, 234. [Google Scholar] [CrossRef] [Green Version]
- Wente, N.; Klocke, D.; Paduch, J.H.; Zhang, Y.; Tho Seeth, M.; Zoche-Golob, V.; Reinecke, F.; Mohr, E.; Krömker, V. Associations between Streptococcus uberis strains from the animal environment and clinical bovine mastitis cases. J. Dairy Sci. 2019, 102, 9360–9369. [Google Scholar] [CrossRef] [PubMed]
- Wald, R.; Baumgartner, M.; Gutschireiter, J.; Bazzanella, B.; Lichtmannsperger, K.; Wagner, M.; Wittek, T.; Stessl, B. Comparison of the population structure of Streptococcus uberis mastitis isolates from Austrian small-scale dairy farms and a Slovakian large-scale farm. J. Dairy Sci. 2020, 103, 1820–1830. [Google Scholar] [CrossRef]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel-electrophoresis: Criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coffey, T.J.; Pullinger, G.D.; Urwin, R.; Jolley, K.A.; Wilson, S.M.; Maiden, M.C.; Leigh, J.A. First insights into the evolution of Streptococcus uberis: A multilocus sequence typing scheme that enables investigation of its population biology. Appl. Environ. Microbiol. 2006, 72, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomita, T.; Meehan, B.; Wongkattiya, N.; Malmo, J.; Pullinger, G.; Leigh, J.; Deighton, M. Identification of Streptococcus uberis multilocus sequence types highly associated with mastitis. Appl. Environ. Microbiol. 2008, 74, 114–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, H.M.; Lim, S.K.; Kang, H.M.; Kim, J.M.; Moon, J.S.; Jang, K.C.; Joo, Y.S.; Kang, M.I.; Jung, S.C. Antimicrobial resistance of streptococci isolated from mastitic bovine milk samples in Korea. J. Vet. Diagn. Investig. 2009, 21, 698–701. [Google Scholar] [CrossRef] [PubMed]
- Fessia, A.S.; Dieser, S.A.; Raspanti, C.G.; Odierno, M. Genotyping and study of adherence-related genes of Streptococcus uberis isolates from bovine mastitis. Microb. Pathog. 2019, 130, 295–301. [Google Scholar] [CrossRef]
- Tomazi, T.; Freu, G.; Gomes Alves, B.; de Souza Filho, A.F.; Heinemann, M.B.; Veiga Dos Santos, M. Genotyping and antimicrobial resistance of Streptococcus uberis isolated from bovine clinical mastitis. PLoS ONE 2019, 14, e0223719. [Google Scholar] [CrossRef]
- Vezina, B.; Al-harbi, H.; Ramay, H.R.; Soust, M.; Moore, R.J.; Olchowy, T.W.J.; Alawneh, J.I. Sequence characterisation and novel insights into bovine mastitis-associated Streptococcus uberis in dairy herds. Sci. Rep. 2021, 11, 3046. [Google Scholar] [CrossRef] [PubMed]
- Lollai, S.A.; Ziccheddu, M.; Di Mauro, C.; Manunta, D.; Nudda, A.; Leori, G. Profile and evolution of antimicrobial resistance of ovine mastitis pathogens (1995–2004). Small Rumin. Res. 2008, 74, 249–254. [Google Scholar] [CrossRef]
- Rato, M.G.; Bexiga, R.; Nunes, S.F.; Cavaco, L.M.; Vilela, C.L.; Santos-Sanches, I. Molecular epidemiology and population structure of bovine Streptococcus uberis. J. Dairy Sci. 2008, 91, 4542–4551. [Google Scholar] [CrossRef] [Green Version]
- Shome, B.R.; Bhuvana, M.; Mitra, S.D.; Krithiga, N.; Shome, R.; Velu, D.; Banerjee, A.; Barbuddhe, S.B.; Prabhudas, K.; Rahman, H. Molecular characterization of Streptococcus agalactiae and Streptococcus uberis isolates from bovine milk. Trop. Anim. Health Prod. 2012, 44, 1981–1992. [Google Scholar] [CrossRef]
- Pullinger, G.D.; Coffey, T.J.; Maiden, M.C.; Leigh, J.A. Multilocus-sequence typing analysis reveals similar populations of Streptococcus uberis are responsible for bovine intramammary infections of short and long duration. Vet. Microbiol. 2007, 119, 194–204. [Google Scholar] [CrossRef]
- McDougall, S.; Parkinson, T.J.; Leyland, M.; Anniss, F.M.; Fenwick, S.G. Duration of infection and strain variation in Streptococcus uberis isolated from cows’ milk. J. Dairy Sci. 2004, 87, 2062–2072. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 4th ed.; CLSI Document VET01S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Rato, M.G.; Bexiga, R.; Florindo, C.; Vilela, C.L.; Santos-Sanches, I. Antimicrobial resistance and molecular epidemiology of streptococci from bovine mastitis. Vet. Microbiol. 2013, 161, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Taber, H.W.; Mueller, J.P.; Miller, P.F.; Arrow, A.S. Bacterial uptake of aminoglycoside antibiotics. Microbiol. Rev. 1987, 51, 439–457. [Google Scholar] [CrossRef] [PubMed]
- Kaczorek, E.; Małaczewska, J.; Wójcik, R.; Rękawek, W.; Siwicki, A.K. Phenotypic and genotypic antimicrobial susceptibility pattern of Streptococcus spp. isolated from cases of clinical mastitis in dairy cattle in Poland. J. Dairy Sci. 2017, 100, 6442–6453. [Google Scholar] [CrossRef]
- Ruegg, P.L.; Oliveira, L.; Jin, W.; Okwumabua, O. Phenotypic antimicrobial susceptibility and occurrence of selected resistance genes in gram-positive mastitis pathogens isolated from Wisconsin dairy cows. J. Dairy Sci. 2015, 98, 4521–4534. [Google Scholar] [CrossRef]
- Gao, J.; Yu, F.Q.; Luo, L.P.; He, J.Z.; Hou, R.G.; Zhang, H.Q.; Li, S.M.; Su, J.L.; Han, B. Antibiotic resistance of Streptococcus agalactiae from cows with mastitis. Vet. J. 2012, 194, 423–424. [Google Scholar] [CrossRef]
- Dogan, B.; Schukken, Y.H.; Santisteban, C.; Boor, K.J. Distribution of serotypes and antimicrobial resistance genes among Streptococcus agalactiae isolates from bovine and human hosts. J. Clin. Microbiol. 2005, 43, 5899–5906. [Google Scholar] [CrossRef] [Green Version]
- Onni, T.; Sanna, G.; Larsen, J.; Tola, S. Antimicrobial susceptibilities and population structure of Staphylococcus epidermidis associated with ovine mastitis. Vet. Microbiol. 2011, 148, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Francisco, A.P.; Bugalho, M.; Ramirez, M.; Carrico, J.A. Global optimal eBURST analysis of multilocus typing data using a graphic matroid approach. BMC Bioinform. 2009, 10, 152–167. [Google Scholar] [CrossRef] [Green Version]
- Poyart-Salmeron, C.; Carlier, C.; Trieu-Cuot, P.; Courtieu, A.L.; Courvalin, P. Transferable plasmid-mediated antibiotic resistance in Listeria monocytogenes. Lancet 1990, 335, 1422–1426. [Google Scholar] [CrossRef]
- Poyart, C.; Celli, J.; Trieu-Cuot, P. Conjugative transposition of Tn916-related elements from Enterococcus faecalis to Escherichia coli and Pseudomonas fluorescens. Antimicrob. Agents Chemother. 1995, 39, 500–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vesterholm-Nielsen, M.; Larsen, M.O.; Olsen, J.E.; Aarestrup, F.M. Occurrence of the blaZ gene in penicillin resistant Staphylococcus aureus isolated from bovine mastitis in Denmark. Acta Vet. Scand. 1999, 40, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.B.; Frimodt-Moller, N.; Aarestrup, F.M. Presence of erm gene classes in gram-positive bacteria of animal and human origin in Denmark. FEMS Microbiol. Lett. 1999, 170, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Villasenor-Sierra, A.; Katahira, E.; Jaramillo-Valdivia, A.N.; Barajas-Garcia Mde, L.; Bryant, A.; Morfin-Otero, R.; Marquez-Diaz, F.; Tinoco, J.C.; Sanchez-Corona, J.; Stevens, D.L. Phenotypes and genotypes of erythromycin-resistant Streptococcus pyogenes strains isolated from invasive and non-invasive infections from Mexico and the USA during 1999–2010. Int. J. Infect. Dis. 2012, 16, 178–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, F.; Malik, S.A.; Ahmed, J.; Ullah, F.; Shah, S.M.; Ayaz, M.; Hussain, S.; Khatoon, L. Investigation of the genetic basis of tetracycline resistance in Staphylococcus aureus from Pakistan. Trop. J. Pharm. Res. 2012, 11, 925–931. [Google Scholar] [CrossRef] [Green Version]
- Aarestrup, F.M.; Agerso, Y.; Gerner-Smidt, P.; Madsen, M.; Jensen, L.B. Comparison of antimicrobial resistance phenotypes and resistance genes in Enterococcus faecalis and Enterococcus faecium from humans in the community, broilers, and pigs in Denmark. Diagn. Microbiol. Infect. Dis. 2000, 37, 127–137. [Google Scholar] [CrossRef]
- Liu, L.C.; Tsai, J.C.; Hsueh, P.R.; Tseng, S.P.; Hung, W.C.; Chen, H.J.; Teng, L.J. Identification of tet(S) gene area in tetracycline–resistant Streptococcus dysgalactiae ssp. equisimilis clinical isolates. J. Antimicrob. Agents Chemother. 2008, 61, 453–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosa, N.M.; Duprè, I.; Azara, E.; Longheu, C.M.; Tola, S. Molecular Typing and Antimicrobial Susceptibility Profiles of Streptococcus uberis Isolated from Sheep Milk. Pathogens 2021, 10, 1489. https://doi.org/10.3390/pathogens10111489
Rosa NM, Duprè I, Azara E, Longheu CM, Tola S. Molecular Typing and Antimicrobial Susceptibility Profiles of Streptococcus uberis Isolated from Sheep Milk. Pathogens. 2021; 10(11):1489. https://doi.org/10.3390/pathogens10111489
Chicago/Turabian StyleRosa, Nives Maria, Ilaria Duprè, Elisa Azara, Carla Maria Longheu, and Sebastiana Tola. 2021. "Molecular Typing and Antimicrobial Susceptibility Profiles of Streptococcus uberis Isolated from Sheep Milk" Pathogens 10, no. 11: 1489. https://doi.org/10.3390/pathogens10111489