
Analysis of Schistosoma mansoni Extracellular Vesicles Surface Glycans Reveals Potential Immune Evasion Mechanism and New Insights on Their Origins of Biogenesis

 , ,
, ,
Abstract
:1. Introduction
2. Results
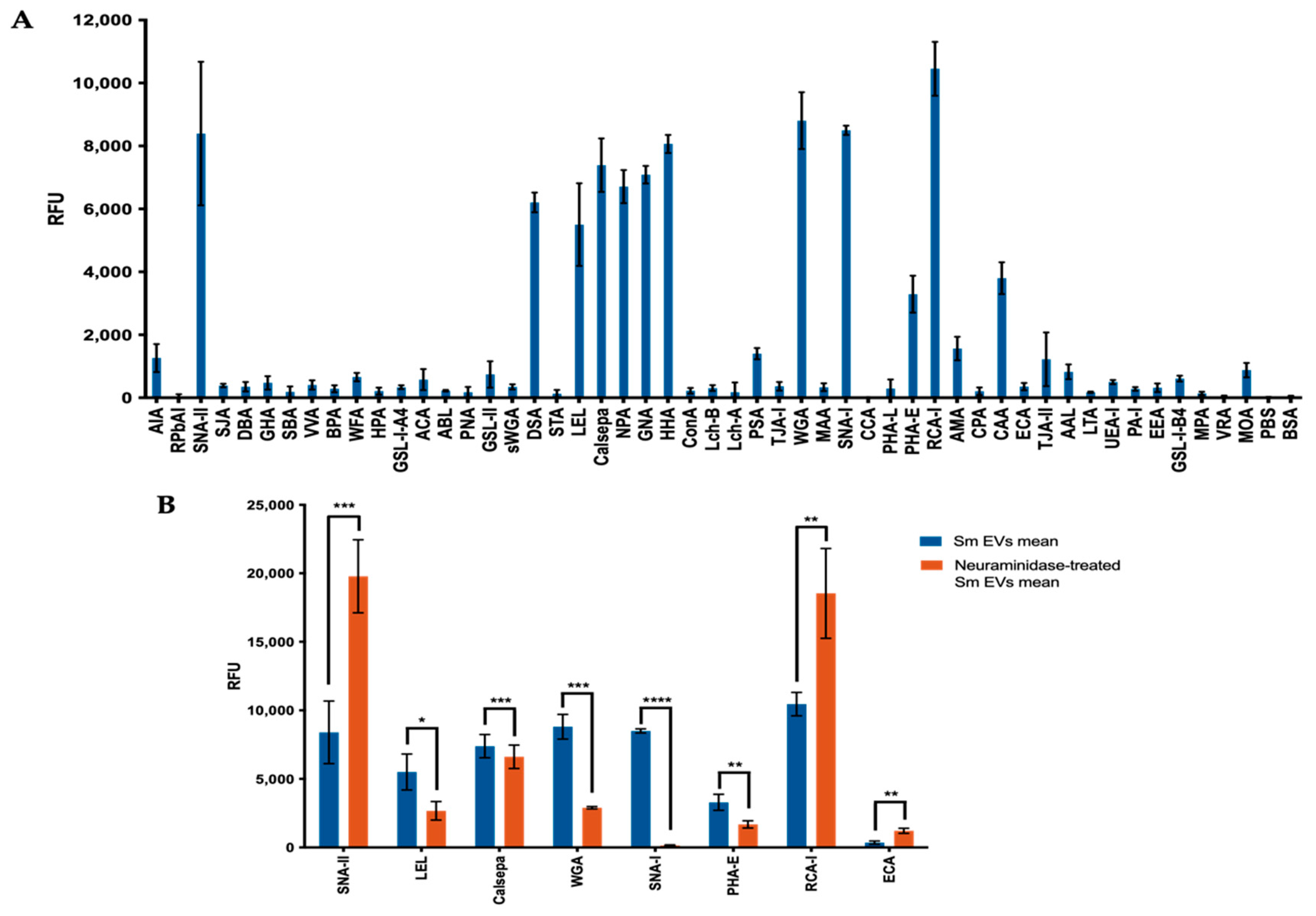
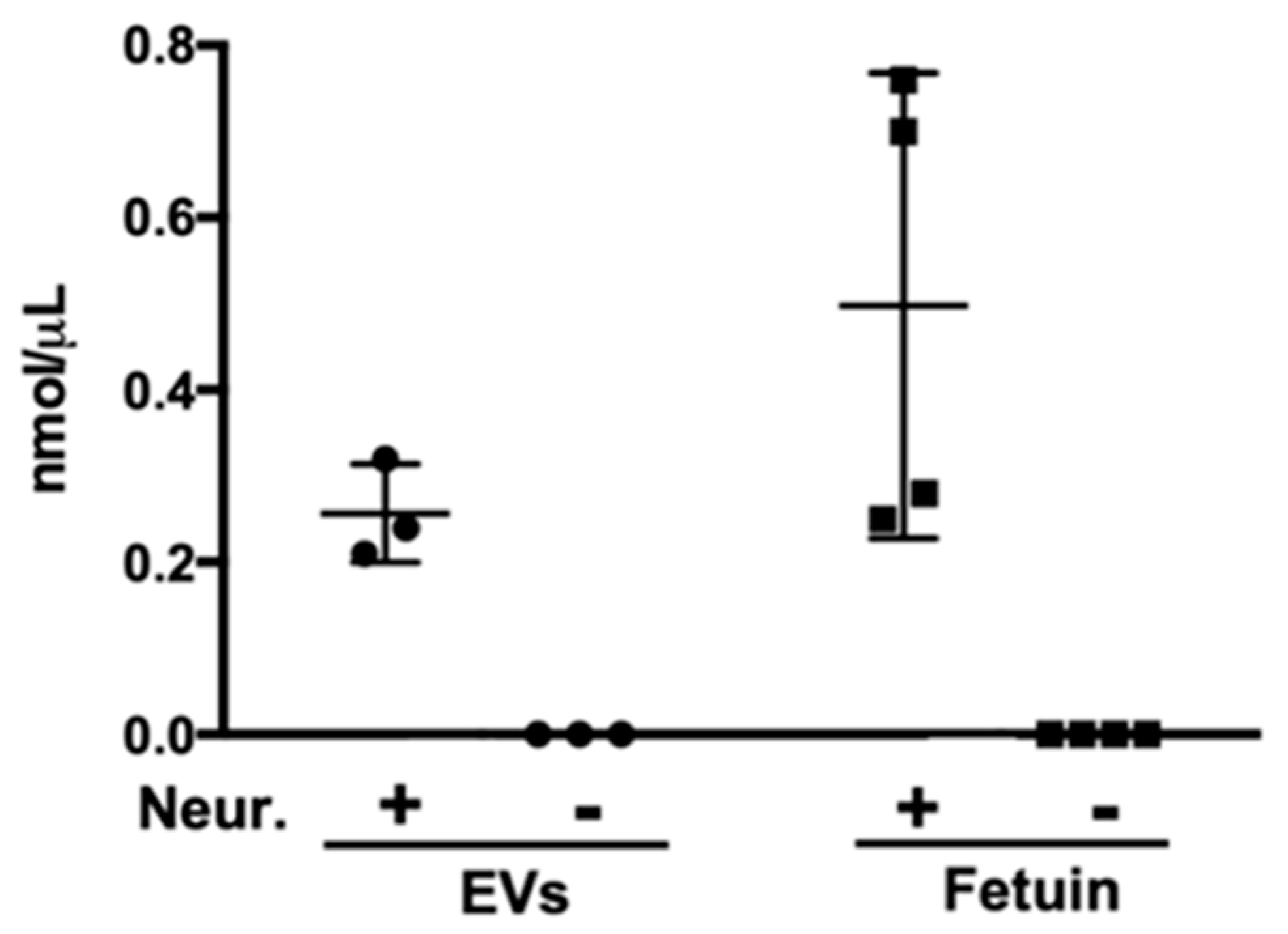
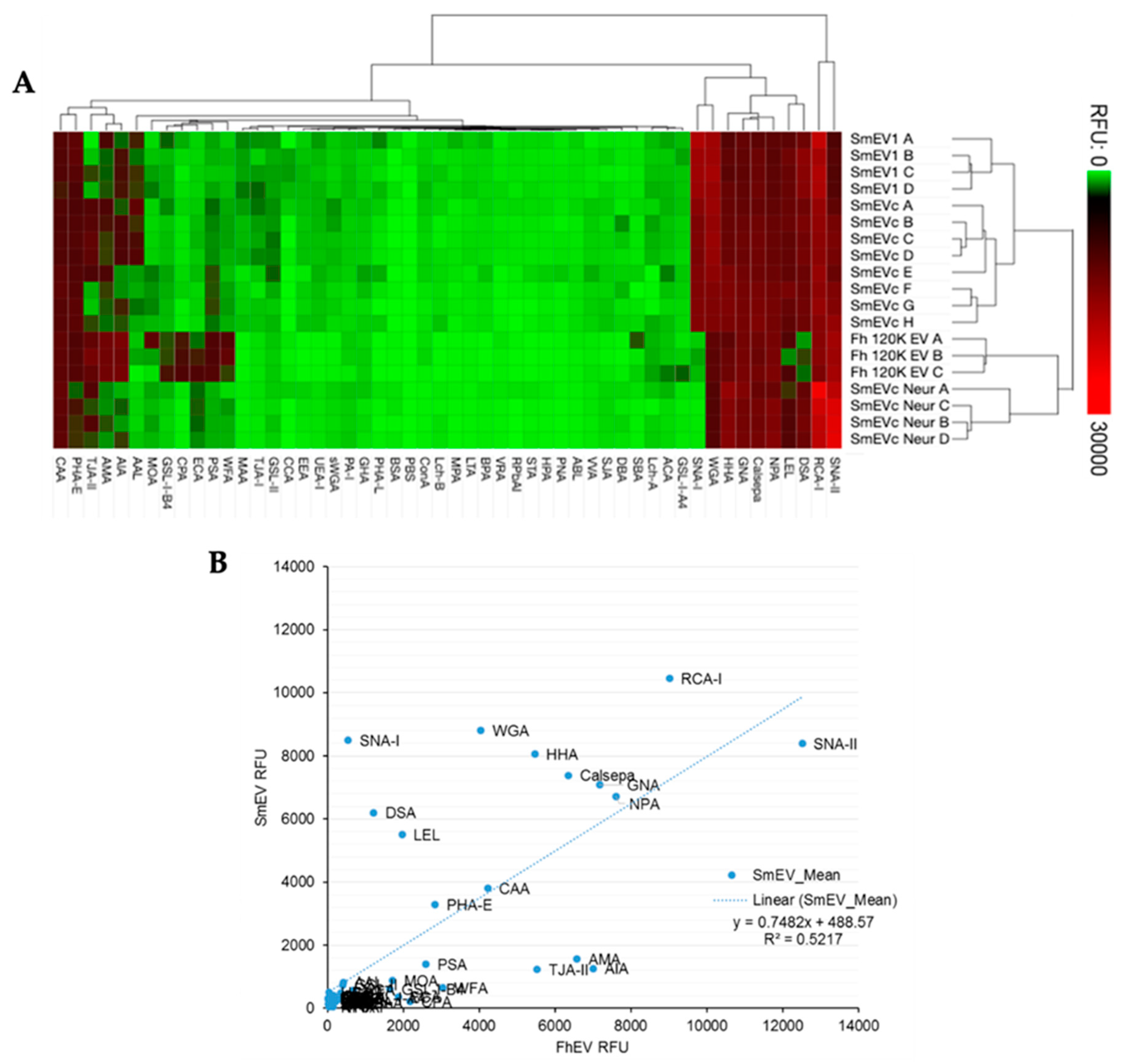
2.1. Sialic Acid Residues Are Detected on Schistosoma mansoni-Derived EVs
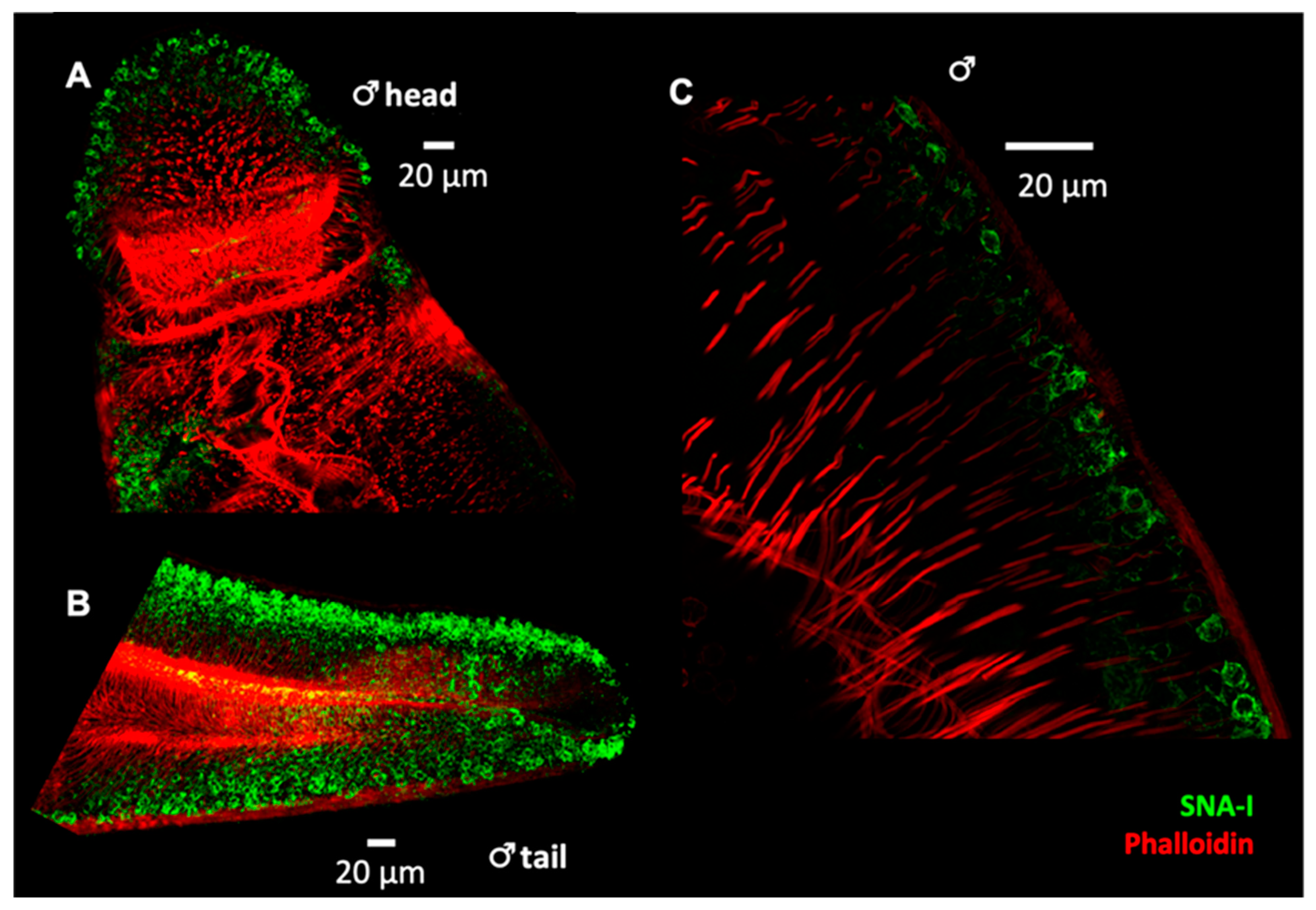
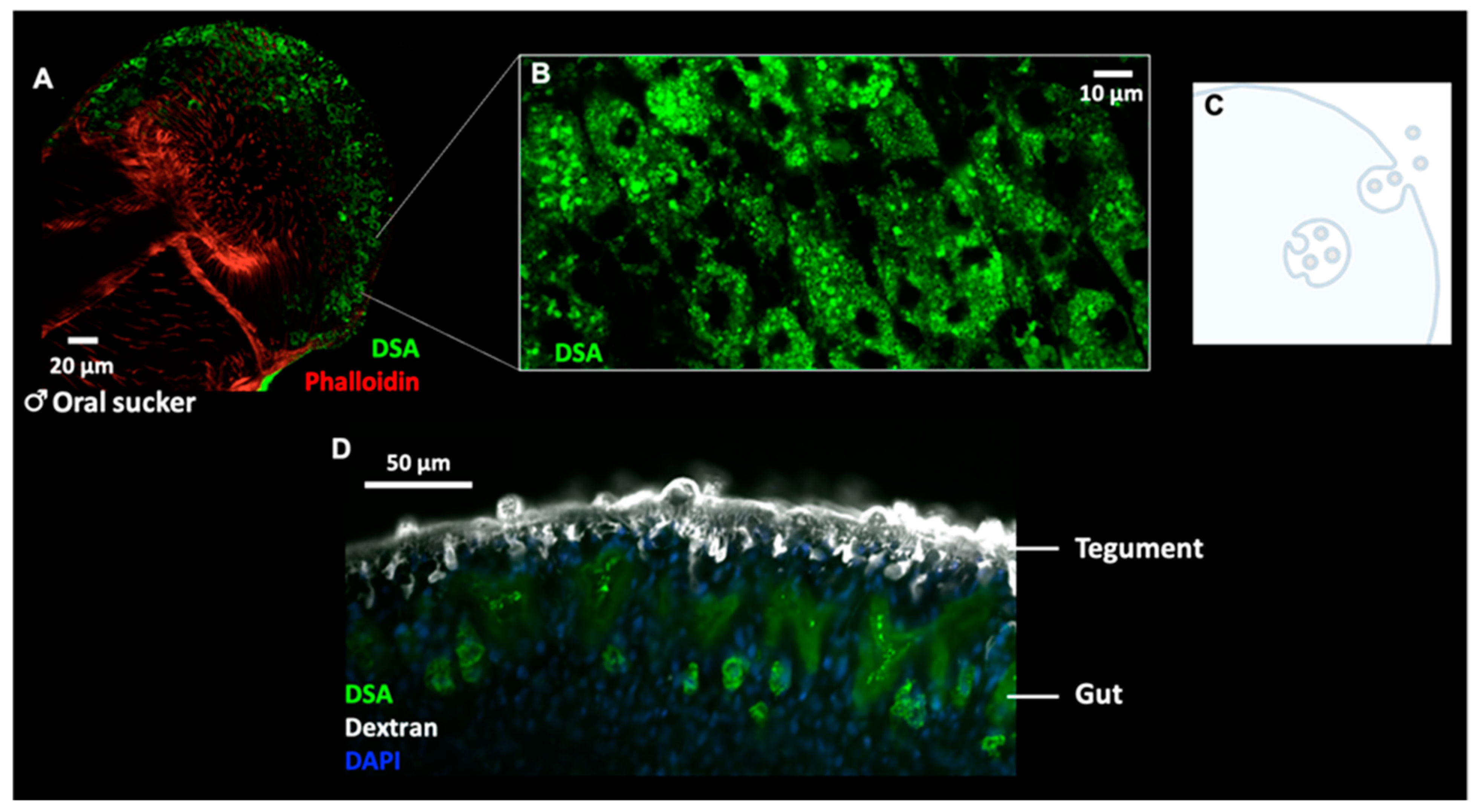
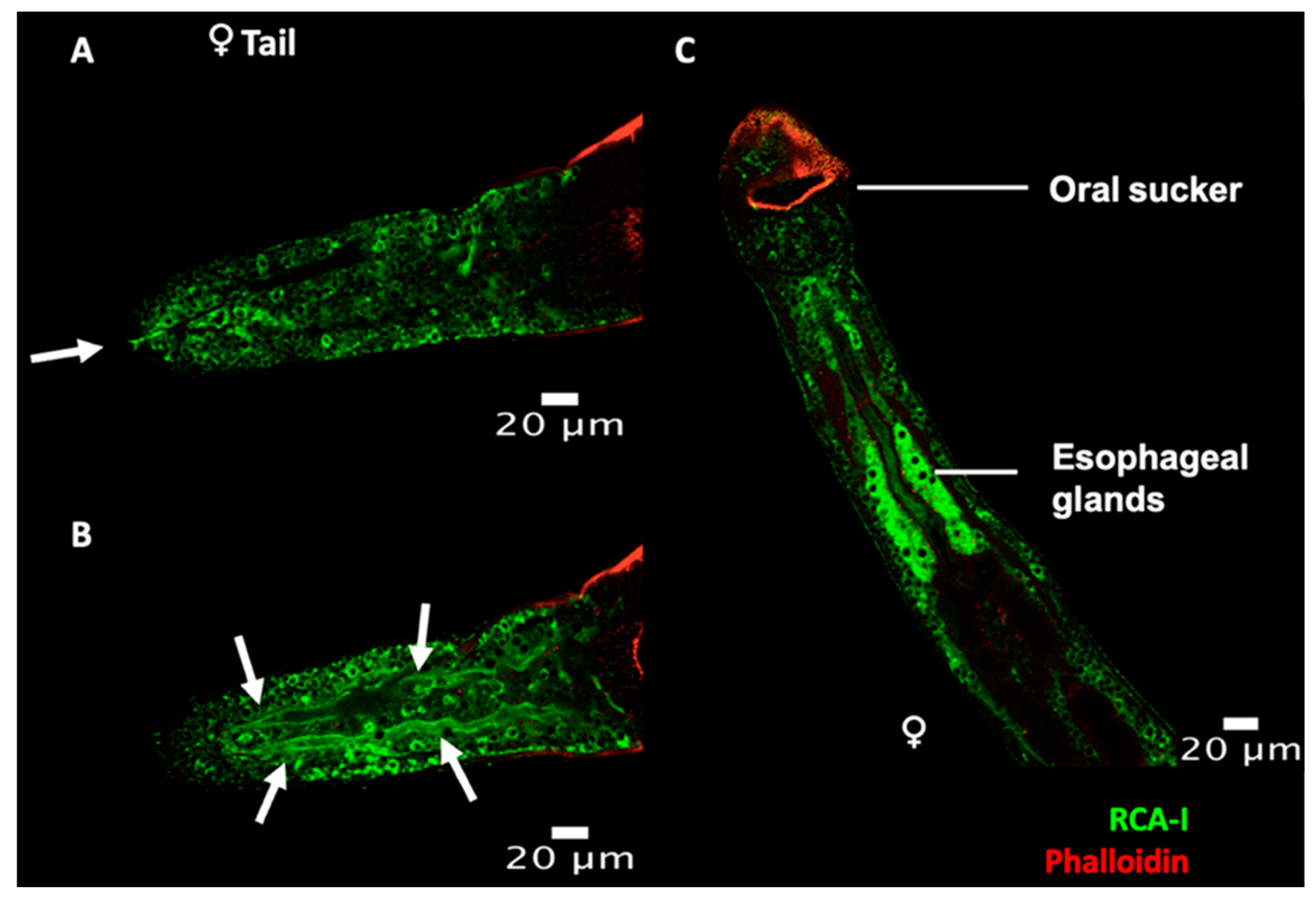
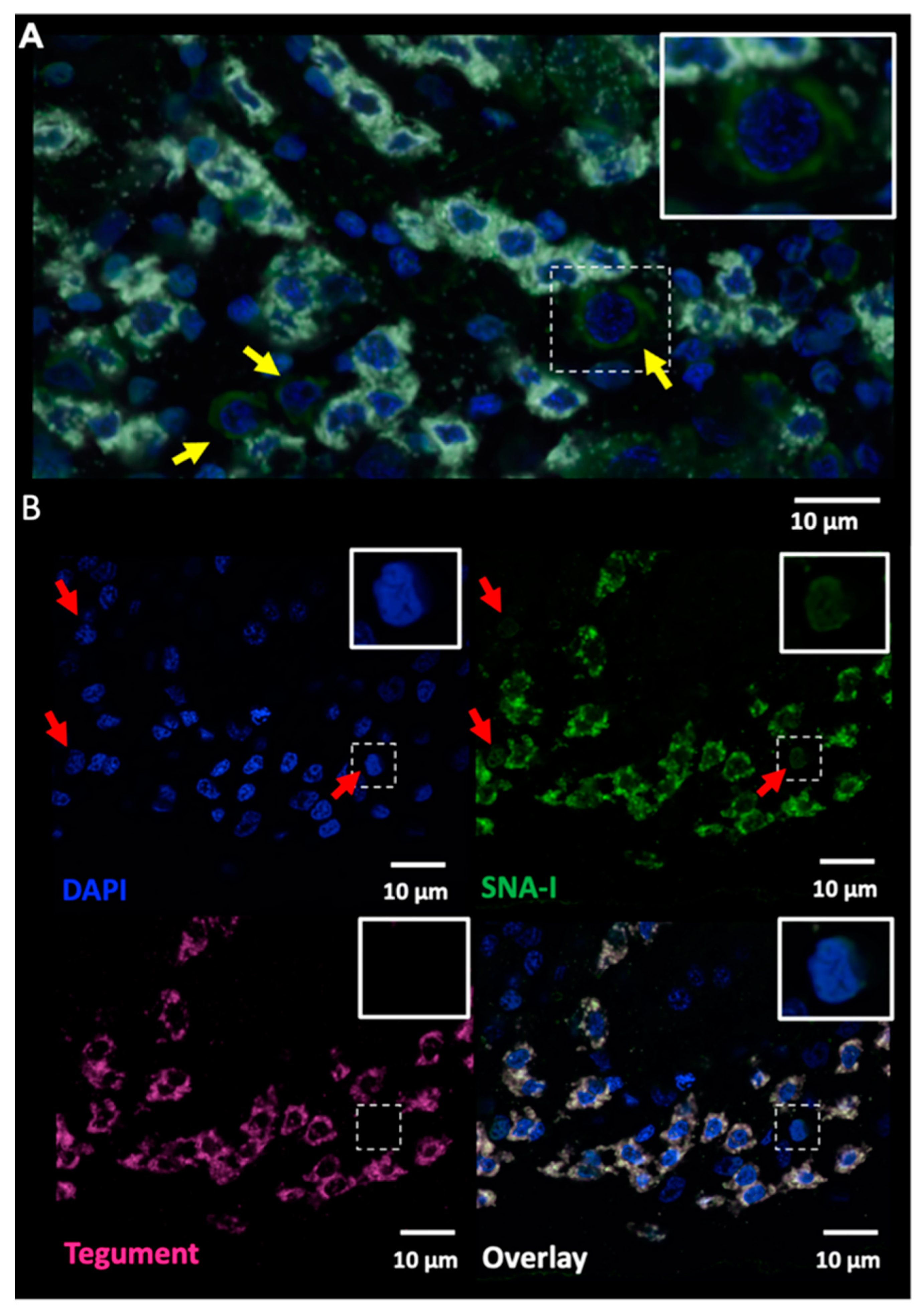
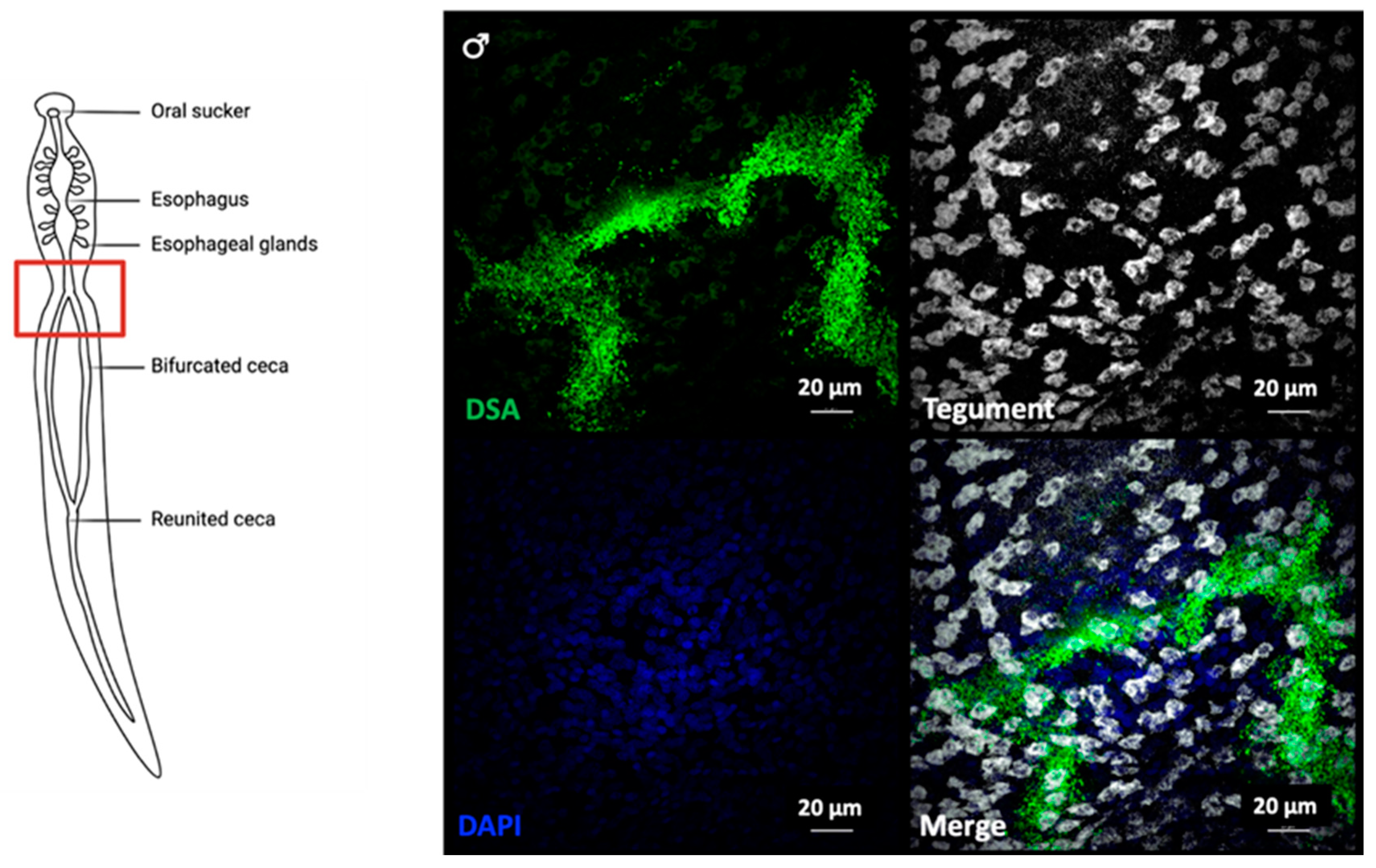
2.2. Lectin Immunocytochemistry and In Situ Hybridisation Reveal Sub-Tegumental Cells as a Potential Source of EVs in Schistosoma mansoni
3. Discussion
4. Materials and Methods
4.1. Parasites
4.2. Isolation of Adult S. mansoni Extracellular Vesicles
4.3. Soluble Worm Antigen Preparation
4.4. Profiling of Fluorescently-Labeled EVs with Lectin Microarrays
4.5. Neuraminidase Treatment
4.6. Free Sialic Acid Detection Assay
4.7. Lectin- and Immunofluorescence Microscopy of Parasites
4.8. Fluorescence In Situ Hybridisation (FISH) Lectin Co-Labelling
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mutapi, F.; Maizels, R.; Fenwick, A.; Woolhouse, M. Human schistosomiasis in the post mass drug administration era. Lancet Infect. Dis. 2016, 17, e42–e48. [Google Scholar] [CrossRef] [Green Version]
- Kyu, H.H.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national disability-adjusted life-years (DALYs) for 359 diseases and injuries and healthy life expectancy (HALE) for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1859–1922. [Google Scholar] [CrossRef] [Green Version]
- Pearce, E.; Sher, A. Mechanisms of immune evasion in schistosomiasis. Contrib. Microbiol. Immunol. 1987, 8, 219–232. [Google Scholar]
- Zaccone, P.; Burton, O.T.; Gibbs, S.; Miller, N.; Jones, F.M.; Dunne, D.W.; Cooke, A. Immune Modulation by Schistosoma mansoni Antigens in NOD Mice: Effects on Both Innate and Adaptive Immune Systems. J. Biomed. Biotechnol. 2010, 2010, 795210. [Google Scholar] [CrossRef] [Green Version]
- Harris, A.R.; Russell, R.J.; Charters, A.D. A review of schistosomiasis in immigrants in Western Australia, demonstrating the unusual longevity of Schistosoma mansoni. Trans. R. Soc. Trop. Med. Hyg. 1984, 78, 385–388. [Google Scholar] [CrossRef]
- Carson, J.; Gobert, G. Modulation of the Host Immune Response by Schistosome Egg-Secreted Proteins Is a Critical Avenue of Host–Parasite Communication. Pathogens 2021, 10, 863. [Google Scholar] [CrossRef] [PubMed]
- Buck, A.H.; Coakley, G.; Simbari, F.; McSorley, H.J.; Quintana, J.F.; Le Bihan, T.; Kumar, S.; Abreu-Goodger, C.; Lear, M.; Harcus, Y.; et al. Exosomes secreted by nematode parasites transfer small RNAs to mammalian cells and modulate innate immunity. Nat. Commun. 2014, 5, 5488. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Liu, J.; Dao, J.; Lu, K.; Li, H.; Gu, H.; Liu, J.; Feng, X.; Cheng, G. Molecular characterization of S. japonicum exosome-like vesicles reveals their regulatory roles in parasite-host interactions. Sci. Rep. 2016, 6, 25885. [Google Scholar] [CrossRef]
- Cass, C.L.; Johnson, J.R.; Califf, L.L.; Xu, T.; Hernandez, H.J.; Stadecker, M.J.; Yates, J.R., III; Williams, D.L. Proteomic analysis of Schistosoma mansoni egg secretions. Mol. Biochem. Parasitol. 2007, 155, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Carson, J.; Robinson, M.W.; Hsieh, M.H.; Cody, J.; Le, L.; You, H.; McManus, D.P.; Gobert, G.N. A comparative proteomics analysis of the egg secretions of three major schistosome species. Mol. Biochem. Parasitol. 2020, 240, 111322. [Google Scholar] [CrossRef]
- Reamtong, O.; Simanon, N.; Thiangtrongjit, N.S.T.; Limpanont, Y.; Chusongsang, P.; Chusongsang, Y.; Anuntakarun, S.; Payungporn, S.; Phuphisut, O.; Adisakwattana, P. Proteomic analysis of adult Schistosoma mekongi somatic and excretory-secretory proteins. Acta Trop. 2019, 202, 105247. [Google Scholar] [CrossRef]
- Samoil, V.; Dagenais, M.; Ganapathy, V.; Aldridge, J.; Glebov, A.; Jardim, A.; Ribeiro, P. Vesicle-based secretion in schistosomes: Analysis of protein and microRNA (miRNA) content of exosome-like vesicles derived from Schistosoma mansoni. Sci. Rep. 2018, 8, 3286. [Google Scholar] [CrossRef] [PubMed]
- Sotillo, J.; Pearson, M.; Potriquet, J.; Becker, L.; Pickering, D.; Mulvenna, J.; Loukas, A. Extracellular vesicles secreted by Schistosoma mansoni contain protein vaccine candidates. Int. J. Parasitol. 2016, 46, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Li, Z.; Shen, J.; Liu, Z.; Liang, J.; Wu, X.; Sun, X.; Wu, Z. Exosome-like vesicles derived by Schistosoma japonicum adult worms mediates M1 type immune- activity of macrophage. Parasitol. Res. 2015, 114, 1865–1873. [Google Scholar] [CrossRef]
- Zhu, S.; Wang, S.; Lin, Y.; Jiang, P.; Cui, X.; Wang, X.; Zhang, Y.; Pan, W. Release of extracellular vesicles containing small RNAs from the eggs of Schistosoma japonicum. Parasites Vectors 2016, 9, 574. [Google Scholar] [CrossRef] [Green Version]
- Nowacki, F.C.; Swain, M.T.; Klychnikov, O.I.; Niazi, U.; Ivens, A.; Quintana, J.F.; Hensbergen, P.; Hokke, C.H.; Buck, A.; Hoffmann, K.F. Protein and small non-coding RNA-enriched extracellular vesicles are released by the pathogenic blood fluke Schistosoma mansoni. J. Extracell. Vesicles 2015, 4, 28665. [Google Scholar] [CrossRef] [Green Version]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Regev-Rudzki, N.; Wilson, D.; Carvalho, T.; Sisquella, X.; Coleman, B.M.; Rug, M.; Bursac, D.; Angrisano, F.; Gee, M.; Hill, A.; et al. Cell-Cell Communication between Malaria-Infected Red Blood Cells via Exosome-like Vesicles. Cell 2013, 153, 1120–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tritten, L.; Geary, T.G. Helminth extracellular vesicles in host–parasite interactions. Curr. Opin. Microbiol. 2018, 46, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Sotillo, J.; Robinson, M.W.; Kimber, M.J.; Cucher, M.; Ancarola, M.E.; Nejsum, P.; Marcilla, A.; Eichenberger, R.M.; Tritten, L. The protein and microRNA cargo of extracellular vesicles from parasitic helminths—Current status and research priorities. Int. J. Parasitol. 2020, 50, 635–645. [Google Scholar] [CrossRef] [PubMed]
- De La Torre-Escudero, E.; Gerlach, J.; Bennett, A.P.S.; Cwiklinski, K.; Jewhurst, H.L.; Huson, K.M.; Joshi, L.; Kilcoyne, M.; O’Neill, S.; Dalton, J.P.; et al. Surface molecules of extracellular vesicles secreted by the helminth pathogen Fasciola hepatica direct their internalisation by host cells. PLoS Negl. Trop. Dis. 2019, 13, e0007087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, C.; Pazos, R.; Royo, F.; González, E.; Roura-Ferrer, M.; Martinez, A.; Gamiz, J.; Reichardt, N.-C.; Falcón-Pérez, J.M. Assessing the role of surface glycans of extracellular vesicles on cellular uptake. Sci. Rep. 2019, 9, 11920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, C.; Royo, F.; Aizpurua-Olaizola, O.; Pazos, R.; Boons, G.-J.; Reichardt, N.-C.; Falcon-Perez, J.M. Glycosylation of extracellular vesicles: Current knowledge, tools and clinical perspectives. J. Extracell. Vesicles 2018, 7, 1442985. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, A.; Tahara, Y.; Sawada, S.-I.; Sasaki, Y.; Akiyoshi, K. Glycan profiling analysis using evanescent-field fluorescence-assisted lectin array: Importance of sugar recognition for cellular uptake of exosomes from mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2017, 491, 701–707. [Google Scholar] [CrossRef]
- Kuipers, M.E.; Nolte-’t Hoen, E.N.; van der Ham, A.J.; Ozir-Fazalalikhan, A.; Nguyen, D.L.; de Korne, C.M.; Koning, R.I.; Tomes, J.J.; Hoffmann, K.F.; Smits, H.H.; et al. DC-SIGN mediated internalisation of glycosylated extracellular vesicles from Schistosoma mansoni increases activation of monocyte-derived dendritic cells. J. Extracell. Vesicles 2020, 9, 1753420. [Google Scholar] [CrossRef] [PubMed]
- Schauer, R.; Shukla, A.K.; Schroder, C.; Müller, E. The anti-recognition function of sialic acids: Studies with erythrocytes and macrophages. Pure Appl. Chem. 1984, 56, 907–921. [Google Scholar] [CrossRef] [Green Version]
- Vimr, E.; Lichtensteiger, C. To sialylate, or not to sialylate: That is the question. Trends Microbiol. 2002, 10, 254–257. [Google Scholar] [CrossRef]
- Hokke, C.H.; van Diepen, A. Helminth glycomics–glycan repertoires and host-parasite interactions. Mol. Biochem. Parasitol. 2017, 215, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Wendt, G.R.; Collins, J.N.; Pei, J.; Pearson, M.S.; Bennett, H.M.; Loukas, A.; Berriman, M.; Grishin, N.V.; Collins, J.J., III. Flatworm-specific transcriptional regulators promote the specification of tegumental progenitors in Schistosoma mansoni. Elife 2018, 7, e33221. [Google Scholar] [CrossRef] [Green Version]
- Marcilla, A.; Trelis, M.; Cortés, A.; Sotillo, J.; Cantalapiedra, F.; Minguez, M.T.; Valero, M.L.; del Pino, M.S.; Muñoz-Antoli, C.; Toledo, R.; et al. Extracellular Vesicles from Parasitic Helminths Contain Specific Excretory/Secretory Proteins and Are Internalized in Intestinal Host Cells. PLoS ONE 2012, 7, e45974. [Google Scholar] [CrossRef]
- Cwiklinski, K.; de la Torre-Escudero, E.; Trelis, M.; Bernal, D.; Dufresne, P.J.; Brennan, G.P.; O’Neill, S.; Tort, J.F.; Paterson, S.; Marcilla, A.; et al. The Extracellular Vesicles of the Helminth Pathogen, Fasciola hepatica: Biogenesis Pathways and Cargo Molecules Involved in Parasite Pathogenesis. Mol. Cell. Proteom. 2015, 14, 3258–3273. [Google Scholar] [CrossRef] [Green Version]
- Kifle, D.W.; Pearson, M.S.; Becker, L.; Pickering, D.; Loukas, A.; Sotillo, J. Proteomic analysis of two populations of Schistosoma mansoni-derived extracellular vesicles: 15k pellet and 120k pellet vesicles. Mol. Biochem. Parasitol. 2020, 236, 111264. [Google Scholar] [CrossRef]
- Vallejo, M.C.; Nakayasu, E.S.; Longo, L.V.; Ganiko, L.; Lopes, F.G.; Matsuo, A.L.; Almeida, I.C.; Puccia, R. Lipidomic analysis of extracellular vesicles from the pathogenic phase of Paracoccidioides brasiliensis. PLoS ONE 2012, 7, e39463. [Google Scholar] [CrossRef]
- Royo, F.; Cossío, U.; de Angulo, A.R.; Llop, J.; Falcon-Perez, J.M. Modification of the glycosylation of extracellular vesicles alters their biodistribution in mice. Nanoscale 2019, 11, 1531–1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McVeigh, P.; Cwiklinski, K.; Garcia-Campos, A.; Mulcahy, G.; O’Neill, S.M.; Maule, A.G.; Dalton, J.P. In silico analyses of protein glycosylating genes in the helminth Fasciola hepatica (liver fluke) predict protein-linked glycan simplicity and reveal temporally-dynamic expression profiles. Sci. Rep. 2018, 8, 11700. [Google Scholar] [CrossRef] [PubMed]
- Khoo, K.-H.; Nieto, A.; Morris, H.R.; Dell, A. Structural characterization of the N-glycans from Echinococcus granulosus hydatid cyst membrane and protoscoleces. Mol. Biochem. Parasitol. 1997, 86, 237–248. [Google Scholar] [CrossRef]
- Lee, J.J.; Dissanayake, S.; Panico, M.; Morris, H.R.; Dell, A.; Haslam, S.M. Mass spectrometric characterisation of Taenia crassiceps metacestode N-glycans. Mol. Biochem. Parasitol. 2005, 143, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Ravidà, A.; Aldridge, A.M.; Driessen, N.N.; Heus, F.A.H.; Hokke, C.H.; O’Neill, S.M. Fasciola hepatica Surface Coat Glycoproteins Contain Mannosylated and Phosphorylated N-glycans and Exhibit Immune Modulatory Properties Independent of the Mannose Receptor. PLoS Negl. Trop. Dis. 2016, 10, e0004601. [Google Scholar] [CrossRef] [Green Version]
- Saunderson, S.; Dunn, A.C.; Crocker, P.; McLellan, A.D. CD169 mediates the capture of exosomes in spleen and lymph node. Blood 2014, 123, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Kornilov, R.; Puhka, M.; Mannerström, B.; Hiidenmaa, H.; Peltoniemi, H.; Siljander, P.; Seppänen-Kaijansinkko, R.; Kaur, S. Efficient ultrafiltration-based protocol to deplete extracellular vesicles from fetal bovine serum. J. Extracell. Vesicles 2018, 7, 1422674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerlach, J.Q.; Kilcoyne, M.; Joshi, L. Microarray evaluation of the effects of lectin and glycoprotein orientation and data filtering on glycoform discrimination. Anal. Methods 2013, 6, 440–449. [Google Scholar] [CrossRef] [Green Version]
- Gerlach, J.Q.; Maguire, C.M.; Krüger, A.; Joshi, L.; Prina-Mello, A.; Griffin, M.D. Urinary nanovesicles captured by lectins or antibodies demonstrate variations in size and surface glycosylation profile. Nanomedicine 2017, 12, 1217–1229. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, J.Q.; Krüger, A.; Gallogly, S.; Hanley, S.A.; Hogan, M.C.; Ward, C.J.; Joshi, L.; Griffin, M.D. Surface Glycosylation Profiles of Urine Extracellular Vesicles. PLoS ONE 2013, 8, e74801. [Google Scholar] [CrossRef]
- Collins, J.J., III; Wang, B.; Lambrus, B.G.; Tharp, M.; Iyer, H.; Newmark, P.A. Adult somatic stem cells in the human parasite Schistosoma mansoni. Nature 2013, 494, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.N.; Collins, J.J., III. Tissue degeneration following loss of Schistosoma mansoni cbp1 is associated with increased stem cell proliferation and parasite death in vivo. PLoS Pathog. 2016, 12, e1005963. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lectin | Organism | Common Name | Specificity |
|---|---|---|---|
| SNA-II | Sambucus nigra | Sambucus lectin-II | Gal/GalNAc |
| DSA | Datura stramonium | Jimson weed lectin | GlcNAc |
| LEL | Lycopersicum eculentum | Tomato lectin | GlcNAc-β-(1,4)-GlcNAc |
| Calsepa | Calystegia sepium | Bindweed lectin | Man/Maltose |
| NPA | Narcissus pseudonarcissus | Daffodil lectin | α-(1,6)-Man |
| GNA | Galanthus nivalis | Snowdrop lectin | Man-α(1,3)- |
| HHA | Hippeastrum hybrid | Amarylis agglutinin | Man-α(1,3)-Man-α(1,6)- |
| WGA | Triticum vulgaris | Wheat germ agglutinin | NeuAc/GlcNAc |
| SNA-I | Sambucus nigra | Sambucus lectin-II | Sialic acid-α-(2,6)-Gal(NAc) |
| PHA-E | Phaseolus vulgaris | Kidney bean erythroagglutinin | biantennary, bisecting GlcNAc,β-Gal/Gal-β-(1,4-)GlcNAc |
| RCA-I/120 | Ricinus communis | Castor bean lectin | Gal-β-(1,4)-GlcNAc |
| CAA | Caragana arborescens | Pea tree lectin | Gal-β-(1,4)-GlcNAc |
| Difference 1 | p Value | Significance 2 | +/−? 1 | Lectin | Top Ligands |
|---|---|---|---|---|---|
| 11,386.58 | 0.000285 | *** | Up | SNA-II | Gal/GalNAc |
| 2829.56 | 0.028967 | * | Down | LEL | GlcNAc-β-(1,4)-GlcNAc |
| 775.70 | 0.000462 | *** | Down | Calsepa | Man/Maltose |
| 5911.46 | 0.001086 | *** | Down | WGA | NeuAc/GlcNAc |
| 8346.68 | 0.000001 | **** | Down | SNA-I | Sialic acid-α-(2,6)-Gal(NAc) |
| 1610.86 | 0.003570 | ** | Down | PHA-E | biantennary, bisecting GlcNAc,β-Gal/Gal-β-(1,4-)GlcNAc |
| 8081.83 | 0.009170 | ** | Up | RCA-I | Gal-β-(1,4)-GlcNAc |
| 858.01 | 0.002417 | ** | Up | ECA | Gal-β-(1,4)-GlcNAc oligomers |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dagenais, M.; Gerlach, J.Q.; Wendt, G.R.; Collins, J.J., III; Atkinson, L.E.; Mousley, A.; Geary, T.G.; Long, T. Analysis of Schistosoma mansoni Extracellular Vesicles Surface Glycans Reveals Potential Immune Evasion Mechanism and New Insights on Their Origins of Biogenesis. Pathogens 2021, 10, 1401. https://doi.org/10.3390/pathogens10111401
Dagenais M, Gerlach JQ, Wendt GR, Collins JJ III, Atkinson LE, Mousley A, Geary TG, Long T. Analysis of Schistosoma mansoni Extracellular Vesicles Surface Glycans Reveals Potential Immune Evasion Mechanism and New Insights on Their Origins of Biogenesis. Pathogens. 2021; 10(11):1401. https://doi.org/10.3390/pathogens10111401
Chicago/Turabian StyleDagenais, Maude, Jared Q. Gerlach, George R. Wendt, James J. Collins, III, Louise E. Atkinson, Angela Mousley, Timothy G. Geary, and Thavy Long. 2021. "Analysis of Schistosoma mansoni Extracellular Vesicles Surface Glycans Reveals Potential Immune Evasion Mechanism and New Insights on Their Origins of Biogenesis" Pathogens 10, no. 11: 1401. https://doi.org/10.3390/pathogens10111401