
Gender Differences in Social Networks Based on Prevailing Kinship Norms in the Mosuo of China

 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
In summary, we propose that men and women are equally social, but their sociality is directed differently. To caricature, female sociality is dyadic, whereas male sociality is tribal. In other words, men seek social connection in a broad group with multiple people, particularly by competing for a good position in a status hierarchy; women, in contrast, seek social connection in close personal relationships based on mutual, dyadic intimacy.
2. Methods and Study Site
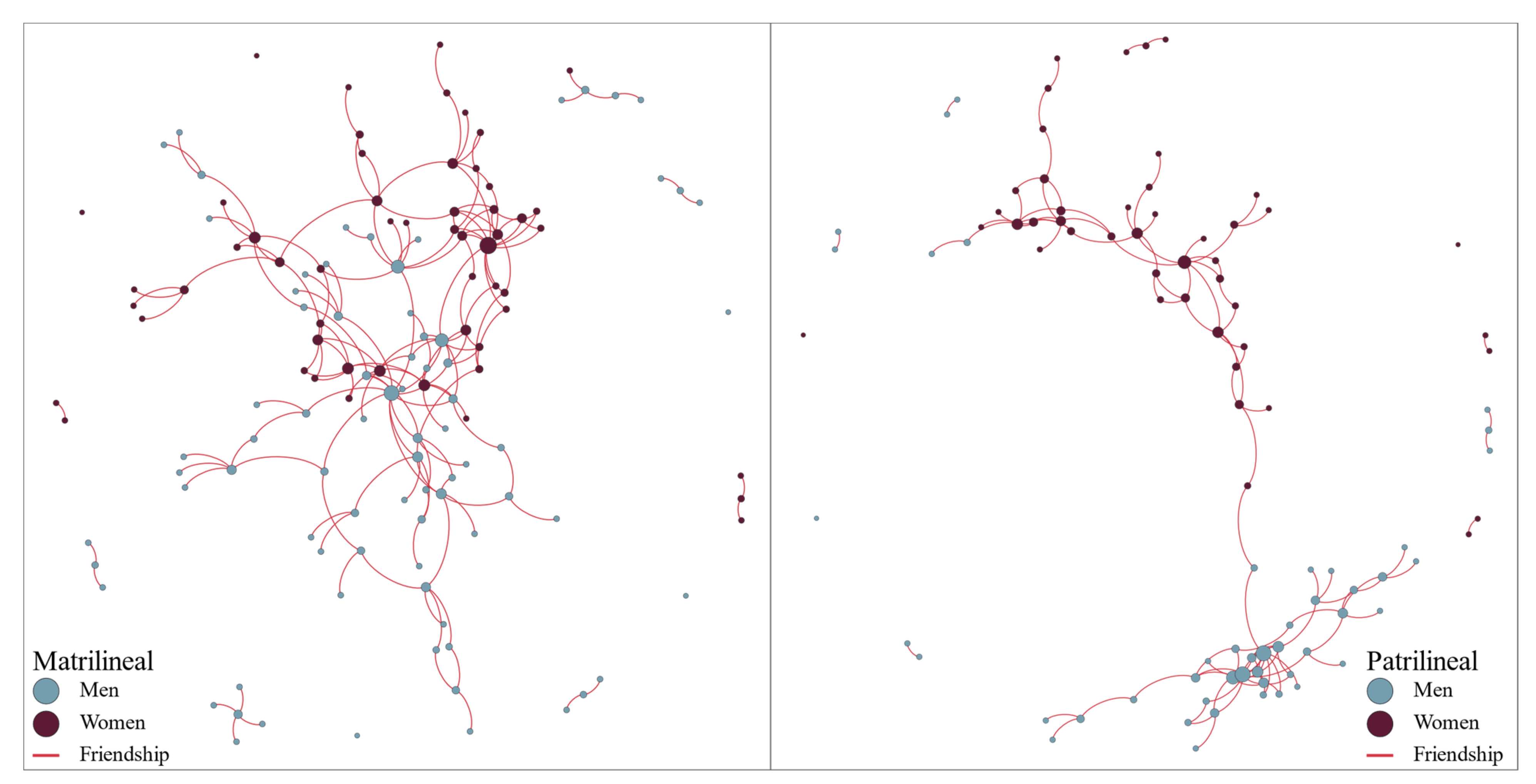
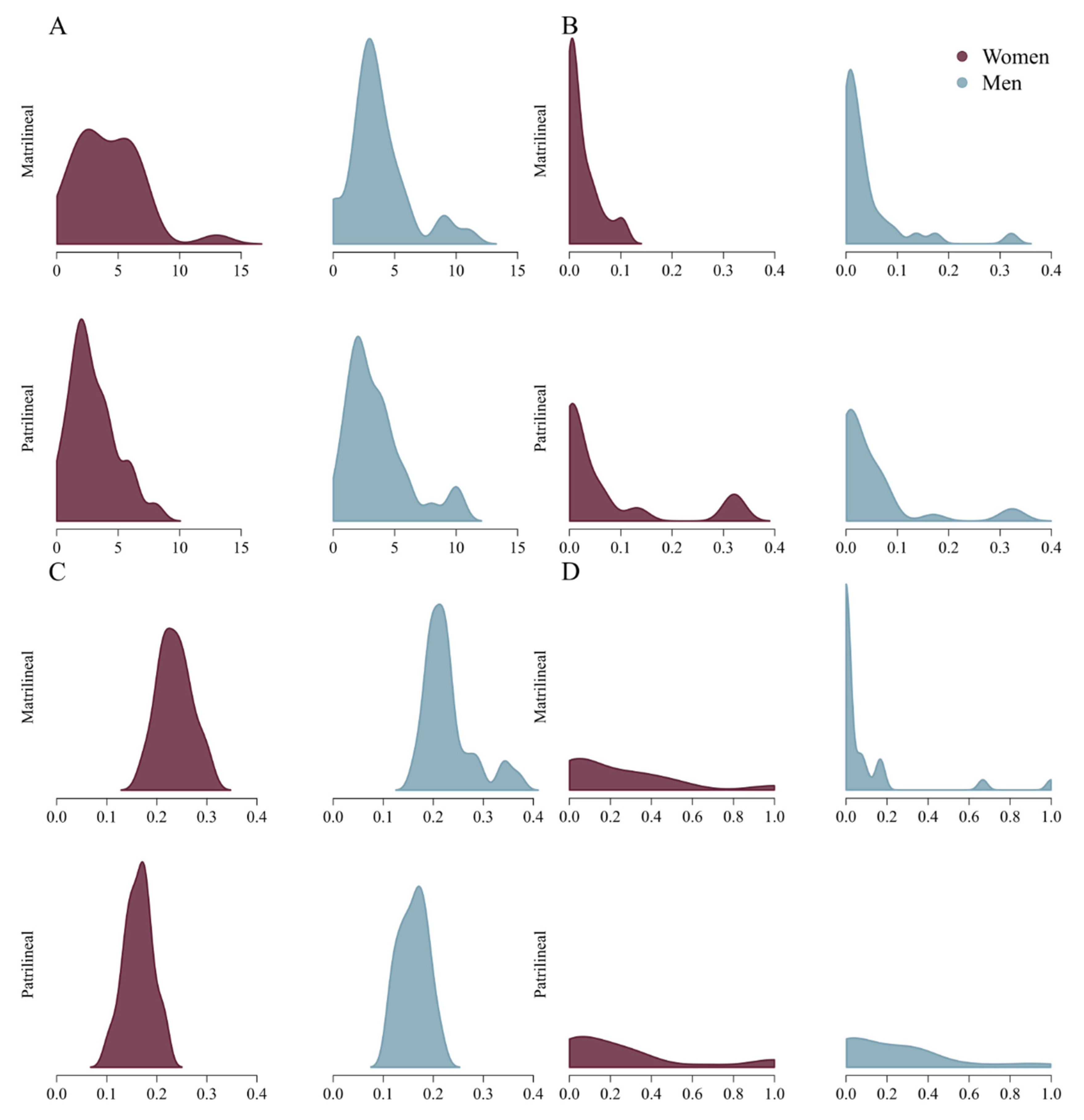
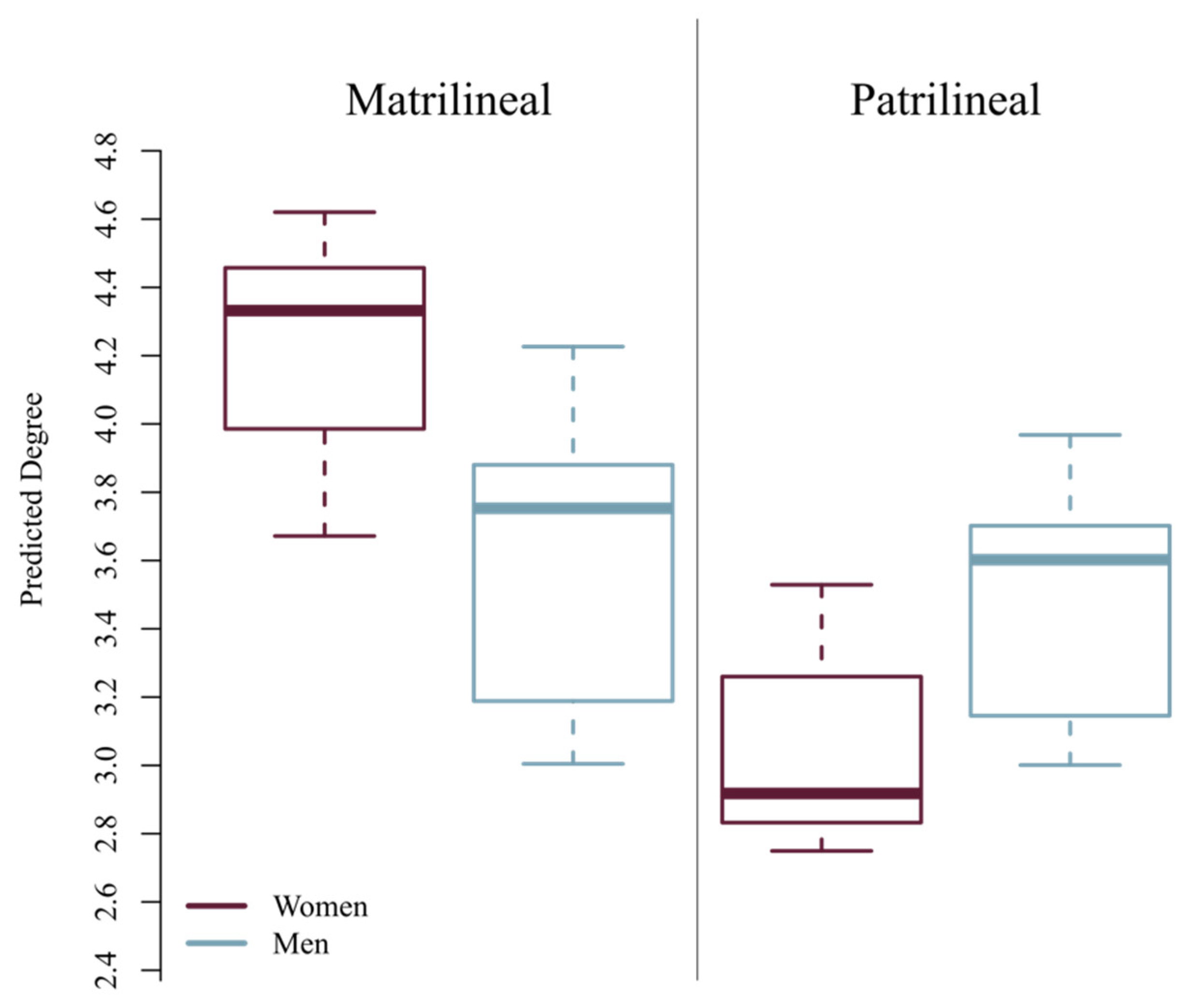
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
| 1 | This modification to standard use of the terms ego and alter is used in this study to reflect the fact that our sampling strategy resulted in individuals having systematically more or less opportunity to have their friendship edges represented in the network. |
References
- Ackerman, Joshua M., Douglas T. Kenrick, and Mark Schaller. 2007. Is friendship akin to kinship? Evolution and Human Behavior 28: 365–74. [Google Scholar] [CrossRef]
- Alami, Sarah, Christopher Von Rueden, Edmond Seabright, Thomas S. Kraft, Aaron D. Blackwell, Jonathan Stieglitz, Hillard Kaplan, and Michael Gurven. 2020. Mother’s social status is associated with child health in a horticulturalist population. Proceedings of the Royal Society B: Biological Sciences 287: 20192783. [Google Scholar] [CrossRef] [Green Version]
- Alesina, Alberto, Paola Giuliano, and Nathan Nunn. 2013. On the Origins of Gender Roles: Women and the Plough. The Quarterly Journal of Economics 128: 469–530. [Google Scholar] [CrossRef]
- Alvard, Michael. 2011. Genetic and Cultural Kinship among the Lamaleran Whale Hunters. Human Nature 22: 89. [Google Scholar] [CrossRef]
- Andersen, Steffen, Erwin Bulte, Uri Gneezy, and John A. List. 2008. Do Women Supply More Public Goods Than Men? Preliminary Experimental Evidence from Matrilineal and Patriarchal Societies. American Economic Review 98: 376–81. [Google Scholar] [CrossRef] [Green Version]
- Apicella, Coren L., Frank W. Marlowe, James H. Fowler, and Nicholas A. Christakis. 2012. Social networks and cooperation in hunter-gatherers. Nature 481: 497–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumeister, Roy F., and Kristin L. Sommer. 1997. What Do Men Want? Gender Differences and Two Spheres of Belongingness: Comment on Cross and Madson. Psychological Bulletin 122: 38–44. [Google Scholar] [CrossRef] [PubMed]
- Becker, Anke. 2019. On the Economic Origins of Restrictions on Women’s Sexuality. “On the Economic Origins of Restrictions on Women’s Sexuality.” SSRN Scholarly Paper ID 3432818. Rochester: Social Science Research Network, Available online: https://papers.ssrn.com/abstract=3432818 (accessed on 7 July 2020).
- Benenson, Joyce F. 1990. Gender differences in social networks. The Journal of Early Adolescence 10: 472–95. [Google Scholar] [CrossRef]
- Benenson, Joyce F. 2019. Sex differences in human peer relationships: A primate’s-eye view. Current Directions in Psychological Science 28: 124–30. [Google Scholar] [CrossRef]
- Benenson, Joyce F., and Helen Abadzi. 2020. Contest versus scramble competition: Sex differences in the quest for status. Power, Status and Hierarchy 33: 62–68. [Google Scholar] [CrossRef]
- Benenson, Joyce F., and Athena Christakos. 2003. The greater fragility of females’ versus males’ closest same-sex friendships. Child Development 74: 1123–29. [Google Scholar] [CrossRef]
- Benenson, Joyce F., Maxwell M. White, Delfina Martinez Pandiani, Lindsay J. Hillyer, Sera Kantor, Henry Markovits, and Richard W. Wrangham. 2018. Competition elicits more physical affiliation between male than female friends. Scientific Reports 8: 1–8. [Google Scholar] [CrossRef] [PubMed]
- Berdahl, Jennifer L., and Cameron Anderson. 2005. Men, women, and leadership centralization in groups over time. Group Dynamics: Theory, Research, and Practice 9: 45. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, Kunal, Asim Ghosh, Daniel Monsivais, Robin I. M. Dunbar, and Kimmo Kaski. 2016. Sex differences in social focus across the life cycle in humans. Royal Society Open Science 3: 160097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bliege Bird, Rebecca, and Douglas W. Bird. 2008. Why Women Hunt: Risk and Contemporary Foraging in a Western Desert Aboriginal Community. Current Anthropology 49: 655–93. [Google Scholar] [CrossRef]
- Blumenfield, Tami, Chun-Yi Sum, Mary K. Shenk, and Siobhán M. Mattison. 2018. Poverty alleviation and mobility in southwest China: Examining effects of market transition and state policies in Mosuo communities. Urban Anthropology and Studies of Cultural Systems and World Economic Development 47: 259–99. [Google Scholar]
- Bradley, Brenda J., Diane M. Doran-Sheehy, Dieter Lukas, Christophe Boesch, and Linda Vigilant. 2004. Dispersed Male Networks in Western Gorillas. Current Biology 14: 510–13. [Google Scholar] [CrossRef] [Green Version]
- Brown, Gillian R., Kevin N. Laland, and Monique Borgerhoff Mulder. 2009. Bateman’s Principles and Human Sex Roles. Trends in Ecology & Evolution 24: 297–304. [Google Scholar]
- Brown, Judith K., and Virginia Kerns. 1985. In Her Prime: A New View of Middle-Aged Women. Greenwood: Bergin & Garvey Publishers. [Google Scholar]
- Campbell, Anne. 2013. A Mind of Her Own: The Evolutionary Psychology of Women. New York: OUP Oxford. [Google Scholar]
- Cheney, Dorothy L., Joan B. Silk, and Robert M. Seyfarth. 2016. Network connections, dyadic bonds and fitness in wild female baboons. Royal Society Open Science 3: 160255. [Google Scholar] [CrossRef] [Green Version]
- Cronk, Lee. 2000. Female-Biased Parental Investment and Growth Performance among the Mukogodo. In Adaptation and Human Behavior. London: Routledge, pp. 203–22. [Google Scholar]
- David-Barrett, Tamas, Anna Rotkirch, James Carney, Isabel Behncke Izquierdo, Jaimie A. Krems, Dylan Townley, Elinor McDaniell, Anna Byrne-Smith, and Robin IM Dunbar. 2015. Women Favour Dyadic Relationships, but Men Prefer Clubs: Cross-Cultural Evidence from Social Networking. PLoS ONE 10: e0118329. [Google Scholar] [CrossRef]
- David-Barrett, Tamas, Janos Kertesz, Anna Rotkirch, Asim Ghosh, Kunal Bhattacharya, Daniel Monsivais, and Kimmo Kaski. 2016. Communication with Family and Friends across the Life Course. PLoS ONE 11: e0165687. [Google Scholar] [CrossRef]
- David-Barrett, Tamas. 2019. Network Effects of Demographic Transition. Scientific Reports 9: 2361. [Google Scholar] [CrossRef]
- Dunbar, R., and A. Machin. 2014. Sex differences in relationship conflict and reconciliation. Journal of Evolutionary Psychology 12: 109–33. [Google Scholar] [CrossRef]
- Dyble, Mark, Gul Deniz Salali, Nikhil Chaudhary, Abigail Page, Daniel Smith, James Thompson, Lucio Vinicius, Ruth Mace, and Andrea Bamberg Migliano. 2015. Sex equality can explain the unique social structure of hunter-gatherer bands. Science 348: 796–98. [Google Scholar] [CrossRef] [PubMed]
- Emery Thompson, Melissa. 2019. How can non-human primates inform evolutionary perspectives on female-biased kinship in humans? Philosophical Transactions of the Royal Society B 374: 20180074. [Google Scholar] [CrossRef] [Green Version]
- Fortunato, Laura, and Marco Archetti. 2010. Evolution of monogamous marriage by maximization of inclusive fitness. Journal of Evolutionary Biology 23: 149–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortunato, Laura. 2012. The evolution of matrilineal kinship organization. Proceedings of the Royal Society B: Biological Sciences 279: 4939–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortunato, Laura. 2019. Lineal kinship organization in cross-specific perspective. Philosophical Transactions of the Royal Society B: Biological Sciences 374: 20190005. [Google Scholar] [CrossRef] [Green Version]
- Friebel, Guido, Marie Lalanne, Bernard Richter, Peter Schwardmann, and Paul Seabright. 2021. Gender differences in social interactions. Journal of Economic Behavior and Organization 186: 33–45. [Google Scholar] [CrossRef]
- Geary, David C. 2006. Sex differences in social behavior and cognition: Utility of sexual selection for hypothesis generation. Hormones and Behavior 49: 273–75. [Google Scholar] [CrossRef] [PubMed]
- Gerkey, Drew, and Lee Cronk. 2014. What is a group? Conceptual clarity can help integrate evolutionary and social scientific research on cooperation. Behavioral and Brain Sciences 37: 260–61. [Google Scholar] [CrossRef] [PubMed]
- Gong, Binglin, and Chun-Lei Yang. 2012. Gender differences in risk attitudes: Field experiments on the matrilineal Mosuo and the patriarchal Yi. Journal of Economic Behavior & Organization 83: 59–65. [Google Scholar] [CrossRef]
- Gong, Binglin, Huibin Yan, and Chun-Lei Yang. 2015. Gender differences in the dictator experiment: Evidence from the matrilineal Mosuo and the patriarchal Yi. Experimental Economics 18: 302–13. [Google Scholar] [CrossRef]
- He, Qiao-Qiao, Jia-Jia Wu, Ting Ji, Yi Tao, and Ruth Mace. 2016. Not leaving home: Grandmothers and male dispersal in a duolocal human society. Behavioral Ecology 27: 1343–52. [Google Scholar] [CrossRef] [PubMed]
- Holden, Clare Janaki, Rebecca Sear, and Ruth Mace. 2003. Matriliny as daughter-biased investment. Evolution and Human Behavior 24: 99–112. [Google Scholar] [CrossRef]
- Hrdy, Sarah Blaffer. 2000. The Optimal Number of Fathers: Evolution, Demography, and History in the Shaping of Female Mate Preferences. Annals of the New York Academy of Sciences 907: 75–96. [Google Scholar] [CrossRef]
- Hrdy, Sarah Blaffer. 2009. Mothers and Others: The Evolutionary Origins of Mutual Understanding. Cambridge: Belknap Press of Harvard University Press. [Google Scholar]
- Ji, Ting, Jia-Jia Wu, Qiao-Qiao He, Jing-Jing Xu, Ruth Mace, and Yi Tao. 2013. Reproductive competition between females in the matrilineal Mosuo of southwestern China. Philosophical Transactions of the Royal Society B: Biological Sciences 368: 20130081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokko, Hanna, and Michael D. Jennions. 2008. Parental investment, sexual selection and sex ratios. Journal of Evolutionary Biology 21: 919–48. [Google Scholar] [CrossRef]
- Koster, Jeremy, Dieter Lukas, David Nolin, Eleanor Power, Alexandra Alvergne, Ruth Mace, Cody T. Ross, Karen Kramer, Russell Greaves, Mark Caudell, and et al. 2019. Kinship ties across the lifespan in human communities. Philosophical Transactions of the Royal Society B: Biological Sciences 374: 20180069. [Google Scholar] [CrossRef] [Green Version]
- Kramer, Karen L. 2010. Cooperative breeding and its significance to the demographic success of humans. Annual Review of Anthropology 39: 417–36. [Google Scholar] [CrossRef] [Green Version]
- Kramer, Karen L., and Russell D. Greaves. 2011. Postmarital Residence and Bilateral Kin Associations among Hunter-Gatherers. Human Nature 22: 41–63. [Google Scholar] [CrossRef]
- Langergraber, Kevin, John Mitani, and Linda Vigilant. 2009. Kinship and social bonds in female chimpanzees (Pan troglodytes). American Journal of Primatology 71: 840–51. [Google Scholar] [CrossRef] [PubMed]
- Lederman, Rena. 1990. Big men, large and small? Towards a comparative perspective. Ethnology 29: 3–15. [Google Scholar] [CrossRef]
- Leonetti, D., Dilip Nath, N. Hemam, and D. Neill. 2005. Kinship organization and grandmother’s impact on reproductive success among the matrilineal Khasi and patrilineal Bengali of N.E. India. In Grandparenthood—The Second Half of Life. Edited by E. Voland. New Brunswick: Rutgers University Press. [Google Scholar]
- Lindenlaub, Ilse, and Anja Prummer. 2021. Network structure and performance. The Economic Journal 131: 851–98. [Google Scholar] [CrossRef]
- Liu, Elaine M., and Sharon Xuejing Zuo. 2019. Measuring the impact of interaction between children of a matrilineal and a patriarchal culture on gender differences in risk aversion. Proceedings of the National Academy of Sciences 116: 6713–19. [Google Scholar] [CrossRef] [Green Version]
- Low, Bobbi S. 2005. Women’s lives there, here, then, now: A review of women’s ecological and demographic constraints cross-culturally. Evolution and Human Behavior 26: 64–87. [Google Scholar] [CrossRef]
- Marsden, Peter V. 2005. Recent developments in network measurement. Models and Methods in Social Network Analysis 8: 30. [Google Scholar]
- Mathieu, Christine. 2003. A History and Anthropological Study of the Ancient Kingdoms of the Sino-Tibetan Borderland—Naxi and Mosuo. Lewiston: The Edwin Mellen Press. [Google Scholar]
- Mattison, Siobhán M. 2010. Economic impacts of tourism and erosion of the visiting system among the Mosuo of Lugu Lake. The Asia Pacific Journal of Anthropology 11: 159–76. [Google Scholar] [CrossRef]
- Mattison, Siobhán M. 2011. Evolutionary contributions to solving the “matrilineal puzzle”: A test of Holden, Sear, and Mace’s model. Human Nature 22: 64–88. [Google Scholar] [CrossRef] [PubMed]
- Mattison, Siobhán M. 2016. Male-provisioning hypothesis. In Encyclopedia of Evolutionary Psychological Science. Edited by V. Weekes-Shackelford, Todd K. Shackelford and V. A. Weekes-Shackelford. Cham: Springer International Publishing, pp. 1–6. [Google Scholar] [CrossRef]
- Mattison, Siobhán M., and Rebecca Sear. 2016. Modernizing evolutionary anthropology. Human Nature 27: 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattison, Siobhán M., Bret Beheim, Bridget Chak, and Peter Buston. 2016. Offspring sex preferences among patrilineal and matrilineal Mosuo in Southwest China revealed by differences in parity progression. Royal Society Open Science 3: 160526. [Google Scholar] [CrossRef] [Green Version]
- Mattison, Siobhán M., Brooke Scelza, and Tami Blumenfield. 2014. Paternal investment and the positive effects of fathers among the matrilineal Mosuo of Southwest China. American Anthropologist 116: 591–610. [Google Scholar] [CrossRef]
- Mattison, Siobhán M., Robert J. Quinlan, and Darragh Hare. 2019. The expendable male hypothesis. Philosophical Transactions of the Royal Society B: Biological Sciences 374: 20180080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattison, Siobhán, Chun-Yi Sum, Adam Reynolds, Gabrielle Baca, Tami Blumenfield, Sara Niedbalski, Ruizhe Liu, Meng Zhang, Lige Liu, Lin Wei, and et al. 2021. Using evolutionary theory to hypothesize a transition from patriliny to matriliny and back again among the ethnic Mosuo of Southwest China. Matrix: A Journal of Matricultural Studies 2: 90–117. [Google Scholar]
- Mengel, Friederike. 2020. Gender differences in networking. The Economic Journal 130: 1842–73. [Google Scholar] [CrossRef]
- Mitani, John C. 2009. Male Chimpanzees Form Enduring and Equitable Social Bonds. Animal Behaviour 77: 633–40. [Google Scholar] [CrossRef]
- Murdock, George P., and Douglas R. White. 1969. Standard Cross-Cultural Sample. Ethnology 8: 329–69. [Google Scholar] [CrossRef]
- Newton-Fisher, Nicholas E. 2006. Female coalitions against male aggression in wild chimpanzees of the Budongo Forest. International Journal of Primatology 27: 1589–99. [Google Scholar] [CrossRef]
- Nolin, David A. 2012. Food-sharing networks in Lamalera, Indonesia: Status, sharing, and signaling. Evolution and Human Behavior 33: 334–45. [Google Scholar] [CrossRef] [Green Version]
- Page, Abigail E., Nikhil Chaudhary, Sylvain Viguier, Mark Dyble, James Thompson, Daniel Smith, Gul D. Salali, Ruth Mace, and Andrea Bamberg Migliano. 2017. Hunter-Gatherer Social Networks and Reproductive Success. Scientific Reports 7: 1153. [Google Scholar] [CrossRef] [Green Version]
- Palchykov, Vasyl, Kimmo Kaski, Janos Kertész, Albert-László Barabási, and Robin I. M. Dunbar. 2012. Sex differences in intimate relationships. Scientific Reports 2: 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkin, Robert. 2021. The Fragility of Marriage in Matrilineal Societies. Book Review, 224. Available online: https://www.anthro.ox.ac.uk/sites/default/files/anthro/documents/media/jaso12_2_2020_224_237.pdf (accessed on 30 June 2021).
- Pearce, Eiluned, Anna Machin, and Robin I. M. Dunbar. 2021. Sex Differences in Intimacy Levels in Best Friendships and Romantic Partnerships. Adaptive Human Behavior and Physiology 7: 1–16. [Google Scholar] [CrossRef]
- Peperkoorn, Leonard S., D. Vaughn Becker, Daniel Balliet, Simon Columbus, Catherine Molho, and Paul A. M. Van Lange. 2020. The prevalence of dyads in social life. PLoS ONE 15: e0244188. [Google Scholar] [CrossRef]
- Power, Eleanor A., and Elspeth Ready. 2019. Cooperation beyond conanguinity: Post-marital residence, delineations of kin, and social support among South Indian Tamils. Philosophical Transactions of the Royal Society B: Biological Sciences 374: 20180070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psylla, Ioanna, Piotr Sapiezynski, Enys Mones, and Sune Lehmann. 2017. The role of gender in social network organization. PLoS ONE 12: e0189873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, Adam Z., Katherine Wander, Chun-Yi Sum, Mingjie Su, Melissa Emery Thompson, Paul L. Hooper, Hui Li, Mary K. Shenk, Kathrine E. Starkweather, Tami Blumenfield, and et al. 2020. Matriliny reverses gender disparities in inflammation and hypertension among the Mosuo of China. Proceedings of the National Academy of Sciences 117: 30324–27. [Google Scholar] [CrossRef]
- Roberts, Sam B. G., and R. I. M. Dunbar. 2015. Managing relationship decay. Human Nature 26: 426–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodseth, Lars, Richard W. Wrangham, Alisa M. Harrigan, Barbara B. Smuts, Ron Dare, Robin Fox, Barbara J. King, P. C. Lee, R. A. Foley, J. C. Muller, and et al. 1991. The Human Community as a Primate Society [and Comments]. Current Anthropology 32: 221–54. [Google Scholar] [CrossRef]
- Rodseth, Lars. 2012. From Bachelor Threat to Fraternal Security: Male Associations and Modular Organization in Human Societies. International Journal of Primatology 33: 1194–1214. [Google Scholar] [CrossRef]
- Rose, Amanda J., and Karen D. Rudolph. 2006. A review of sex differences in peer relationship processes: Potential trade-offs for the emotional and behavioral development of girls and boys. Psychological Bulletin 132: 98–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scelza, Brooke A. 2011. The Place of Proximity. Human Nature 22: 108–27. [Google Scholar] [CrossRef]
- Schneider, David M. 1961. The distinctive features of matrilineal descent groups. In Matrilineal Kinship. Edited by D. M. Schneider and K. Gough. Oakland: University of California Press, pp. 1–29. [Google Scholar]
- Seabright, Paul. 2012. The War of the Sexes: How Conflict and Cooperation Have Shaped Men and Women from Prehistory to the Present. Princeton. Princeton: Princeton University Press. [Google Scholar]
- Shenk, Mary K., and Siobhán M. Mattison. 2011. The rebirth of kinship: Evolutionary and quantitative approaches in the revitalization of a dying field. Human Nature 22: 1–15. [Google Scholar] [CrossRef]
- Shenk, Mary K., Ryan O. Begley, David A. Nolin, and Andrew Swiatek. 2019. Why does matriliny fail? Causes and frequency of matrilineal transitions estimated from a de novo coding of a cross-cultural sample. Philosophical Transactions of the Royal Society B: Biological Sciences, 20190006. [Google Scholar] [CrossRef] [Green Version]
- Shih, Chuan-kang, and Mark R. Jenike. 2002. cultural-historical perspective on the depressed fertility among the matrilineal Moso in Southwest China. Human Ecology 30: 21–47. [Google Scholar] [CrossRef]
- Shih, Chuan-kang. 1993. The Yongning Moso: Sexual Union, Household Organization, Gender and Ethnicity in a Matrilineal Duolocal Society in Southwest China. Ph.D. dissertation, Stanford University, Stanford, CA, USA. [Google Scholar]
- Shih, Chuan-kang. 2010. Quest for Harmony: The Moso Traditions of Sexual Union & Family Life. Stanford: Stanford University Press. [Google Scholar]
- Silk, Joan B. 2007. Social components of fitness in primate groups. Science 317: 1347–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silk, Joan B., Jacinta C. Beehner, Thore J. Bergman, Catherine Crockford, Anne L. Engh, Liza R. Moscovice, Roman M. Wittig, Robert M. Seyfarth, and Dorothy L. Cheney. 2010. Strong and Consistent Social Bonds Enhance the Longevity of Female Baboons. Current Biology 20: 1359–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silk, Joan B., Susan C. Alberts, and Jeanne Altmann. 2003. Social Bonds of Female Baboons Enhance Infant Survival. Science 302: 1231–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, Jeffrey A., and James Moody. 2013. Structural effects of network sampling coverage I: Nodes missing at random. Social Networks 35: 652–68. [Google Scholar] [CrossRef] [Green Version]
- Smith, Jeffrey A., James Moody, and Jonathan H. Morgan. 2017. Network sampling coverage II: The effect of non-random missing data on network measurement. Social Networks 48: 78–99. [Google Scholar] [CrossRef] [Green Version]
- Smuts, Barbara. 1994. The origins of patriarchy: An evolutionary perspective. In Origins of Gender Inequality. Edited by A. Zagarell. Kalamazoo: New Issues Press. [Google Scholar]
- Starkweather, Kathrine E., Mary K. Shenk, and Richard McElreath. 2020. Biological constraints and socioecological influences on women’s pursuit of risk and the sexual division of labor. Evolutionary Human Sciences 2: 1–24. [Google Scholar] [CrossRef]
- Surowiec, Alexandra, Kate T. Snyder, and Nicole Creanza. 2019. A worldwide view of matriliny: Using genetic data to shed light on the evolution of human kinship systems. Philosophical Transactions of the Royal Society B: Biological Sciences 374: 20180077. [Google Scholar] [CrossRef] [Green Version]
- Szell, Michael, and Stefan Thurner. 2013. How women organize social networks different from men. Scientific Reports 3: 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, Matthew Gwynfryn, Ting Ji, Jiajia Wu, QiaoQiao He, Yi Tao, and Ruth Mace. 2018. Kinship underlies costly cooperation in Mosuo villages. Royal Society Open Science 5: 171535. [Google Scholar] [CrossRef] [Green Version]
- Tokuyama, Nahoko, and Takeshi Furuichi. 2016. Do friends help each other? Patterns of female coalition formation in wild bonobos at Wamba. Animal Behaviour 119: 27–35. [Google Scholar] [CrossRef]
- Trivers, Robert L., ed. 1972. Parental investment and sexual selection. In Sexual Selection and the Descent of Man. (Reprinted from B. Campbell (Ed.). New York: Aldine, pp. 136–79. [Google Scholar]
- Vigil, Jacob M. 2007. Asymmetries in the Friendship Preferences and Social Styles of Men and Women. Human Nature 18: 143–61. [Google Scholar] [CrossRef] [PubMed]
- von Rueden, Chris, Sarah Alami, Hillard Kaplan, and Michael Gurven. 2018. Sex differences in political leadership in an egalitarian society. Evolution and Human Behavior 39: 402–11. [Google Scholar] [CrossRef]
- Walsh, Eileen Rose. 2001. The Mosuo—Beyond the Myths of Matriarchy: Gender Transformation and Economic Development. Ph.D. dissertation, Temple University, Setagaya City, Tokyo, Japan. [Google Scholar]
- Wasserman, Stanley, and Katherine Faust. 1994. Social Network Analysis: Methods and Applications. New York: Cambridge University Press. [Google Scholar]
- Werner, Dennis. 1984. Child care and influence among the Mekranoti of Central Brazil. Sex Roles 10: 395–404. [Google Scholar] [CrossRef]
- Winterhalder, Bruce, and Eric Alden Smith. 2000. Analyzing adaptive strategies: Human behavioral ecology at twenty-five. Evolutionary Anthropology 9: 51–72. [Google Scholar] [CrossRef]
- Wood, Brian M., and Frank W. Marlowe. 2011. Dynamics of Postmarital Residence among the Hadza. Human Nature 22: 128–38. [Google Scholar] [CrossRef]
- Wrangham, Richard W. 1980. An Ecological Model of Female-Bonded Primate Groups. Behaviour 75: 262–300. [Google Scholar] [CrossRef]
- Wu, Jia-Jia, Ting Ji, Qiao-Qiao He, Juan Du, and Ruth Mace. 2015. Cooperation is related to dispersal patterns in Sino-Tibetan populations. Nature Communications 6: 8693. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| ALL RESPONDENTS | Matriliny | Patriliny | ||
| Women | Men | Women | Men | |
| N | 31 | 32 | 29 | 29 |
| Individual characteristics: mean (standard deviation) | ||||
| Age (years) | 48.9 (9.1) | 45.9 (14.4) | 43.5 (11.4) | 47.4 (13.3) |
| Education (years) | 1.7 (3.5) | 4.3 (4.2) | 3.5 (4.2) | 5.2 (4.1) |
| Network centralities: median (interquartile range) | ||||
| Degree | 4 (2, 6) | 3 (2, 4.25) | 2 (2, 4) | 3 (2, 4) |
| Betweenness | 0.012 (0.001, 0.037) | 0.016 (0.001, 0.039) | 0.017 (0.000, 0.063) | 0.019 (0.000, 0.066) |
| Closeness | 0.23 (0.21, 0.26) | 0.22 (0.19, 0.23) | 0.16 (0.14, 0.18) | 0.17 (0.14, 0.18) |
| Transitivity | 0.10 (0.00, 0.33) | 0.00 (0.00, 0.067) | 0.17 (0.00, 0.33) | 0.17 (0.00, 0.33) |
| ALL NETWORK NODES | Matriliny | Patriliny | ||
| Women | Men | Women | Men | |
| N | 55 | 78 | 45 | 47 |
| Individual characteristics: mean (standard deviation) | ||||
| Age (years) | 50.0 (12.5) | 42.8 (14.5) | 46.8 (12.9) | 47.8 (12.9) |
| Education (years) | 2.3 (4.2) | 4.0 (4.1) | 2.9 (3.9) | 5.4 (4.0) |
| Network centralities: median (interquartile range) | ||||
| Degree | 2 (1, 4.5) | 1 (1, 3) | 2 (1, 3) | 2 (1, 3.5) |
| Betweenness | 0.001 (0.000, 0.015) | 0.000 (0.000, 0.013) | 0.000 (0.000, 0.034) | 0.000 (0.000, 0.031) |
| Closeness | 0.24 (0.21, 0.26) | 0.23 (0.20, 0.26) | 0.16 (0.15, 0.18) | 0.16 (0.14, 0.17) |
| Transitivity | 0.23 (0.00, 0.63) | 0.00 (0.00, 0.00) | 0.17 (0.00, 0.33) | 0.17 (0.00, 0.33) |
| Distributions Being Compared | Mean 1 | Mean 2 | p | |
|---|---|---|---|---|
| Degree | ||||
| Mat. Women | Mat. Men | 3.018 | 2.205 | 0.019 * |
| Pat. Men | Pat. Women | 2.596 | 2.356 | 0.315 |
| Pat. Men | Mat. Men | 2.596 | 2.205 | 0.170 |
| Mat. Women | Pat. Women | 3.018 | 2.356 | 0.069 † |
| Betweenness | ||||
| Mat. Men | Mat. Women | 0.017 | 0.014 | 0.351 |
| Pat. Women | Pat. Men | 0.045 | 0.031 | 0.214 |
| Pat. Men | Mat. Men | 0.031 | 0.017 | 0.096 † |
| Pat. Women | Mat. Women | 0.045 | 0.014 | 0.009 * |
| Closeness | ||||
| Mat. Women | Mat. Men | 0.239 | 0.233 | 0.224 |
| Pat. Women | Pat. Men | 0.163 | 0.158 | 0.221 |
| Mat. Men | Pat. Men | 0.233 | 0.158 | 0.000 * |
| Mat. Women | Pat. Women | 0.239 | 0.163 | 0.000 * |
| Transitivity | ||||
| Mat. Women | Mat. Men | 0.368 | 0.067 | 0.000 * |
| Pat. Women | Pat. Men | 0.258 | 0.247 | 0.498 |
| Pat. Men | Mat. Men | 0.247 | 0.067 | 0.001 * |
| Mat. Women | Pat. Women | 0.368 | 0.258 | 0.211 |
| Variable | Coefficient | Standard Error | 95% CI | p |
|---|---|---|---|---|
| Intercept | 0.04 | 0.20 | (−0.35, 0.42) | 0.84 |
| Control for primary respondent | 1.11 | 0.11 | (0.90, 1.33) | <0.0001 * |
| Control for secondary respondent | −0.19 | 0.10 | (−0.39, 0.01) | 0.065 † |
| Age | 0.00 | 0.00 | (−0.00, 0.01) | 0.37 |
| Gender (man) | −0.04 | 0.09 | (−0.21, 0.12) | 0.62 |
| Community (matrilineal) | 0.15 | 0.09 | (−0.02, 0.32) | 0.095 † |
| Intercept | 0.00 | 0.20 | (−0.40, 0.40) | 1.0 |
| Control for primary respondent | 1.10 | 0.11 | (0.89, 1.31) | <0.0001 * |
| Control for secondary respondent | −0.17 | 0.10 | (−0.37, 0.03) | 0.11 |
| Age | 0.00 | 0.00 | (−0.00, 0.00) | 0.45 |
| Gender (man) | 0.09 | 0.14 | (−0.18, 0.35) | 0.53 |
| Community (matrilineal) | 0.26 | 0.13 | (0.01, 0.52) | 0.04 * |
| Gender (man) * Community (matrilineal) | −0.21 | 0.18 | (−0.56, 0.13) | 0.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mattison, S.M.; MacLaren, N.G.; Liu, R.; Reynolds, A.Z.; Baca, G.D.; Mattison, P.M.; Zhang, M.; Sum, C.-Y.; Shenk, M.K.; Blumenfield, T.; et al. Gender Differences in Social Networks Based on Prevailing Kinship Norms in the Mosuo of China. Soc. Sci. 2021, 10, 253. https://doi.org/10.3390/socsci10070253
Mattison SM, MacLaren NG, Liu R, Reynolds AZ, Baca GD, Mattison PM, Zhang M, Sum C-Y, Shenk MK, Blumenfield T, et al. Gender Differences in Social Networks Based on Prevailing Kinship Norms in the Mosuo of China. Social Sciences. 2021; 10(7):253. https://doi.org/10.3390/socsci10070253
Chicago/Turabian StyleMattison, Siobhán M., Neil G. MacLaren, Ruizhe Liu, Adam Z. Reynolds, Gabrielle D. Baca, Peter M. Mattison, Meng Zhang, Chun-Yi Sum, Mary K. Shenk, Tami Blumenfield, and et al. 2021. "Gender Differences in Social Networks Based on Prevailing Kinship Norms in the Mosuo of China" Social Sciences 10, no. 7: 253. https://doi.org/10.3390/socsci10070253