
A Review of Ant Nests and Their Implications for Architecture

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Biological Perspective
2.1. Underground Ant Nest
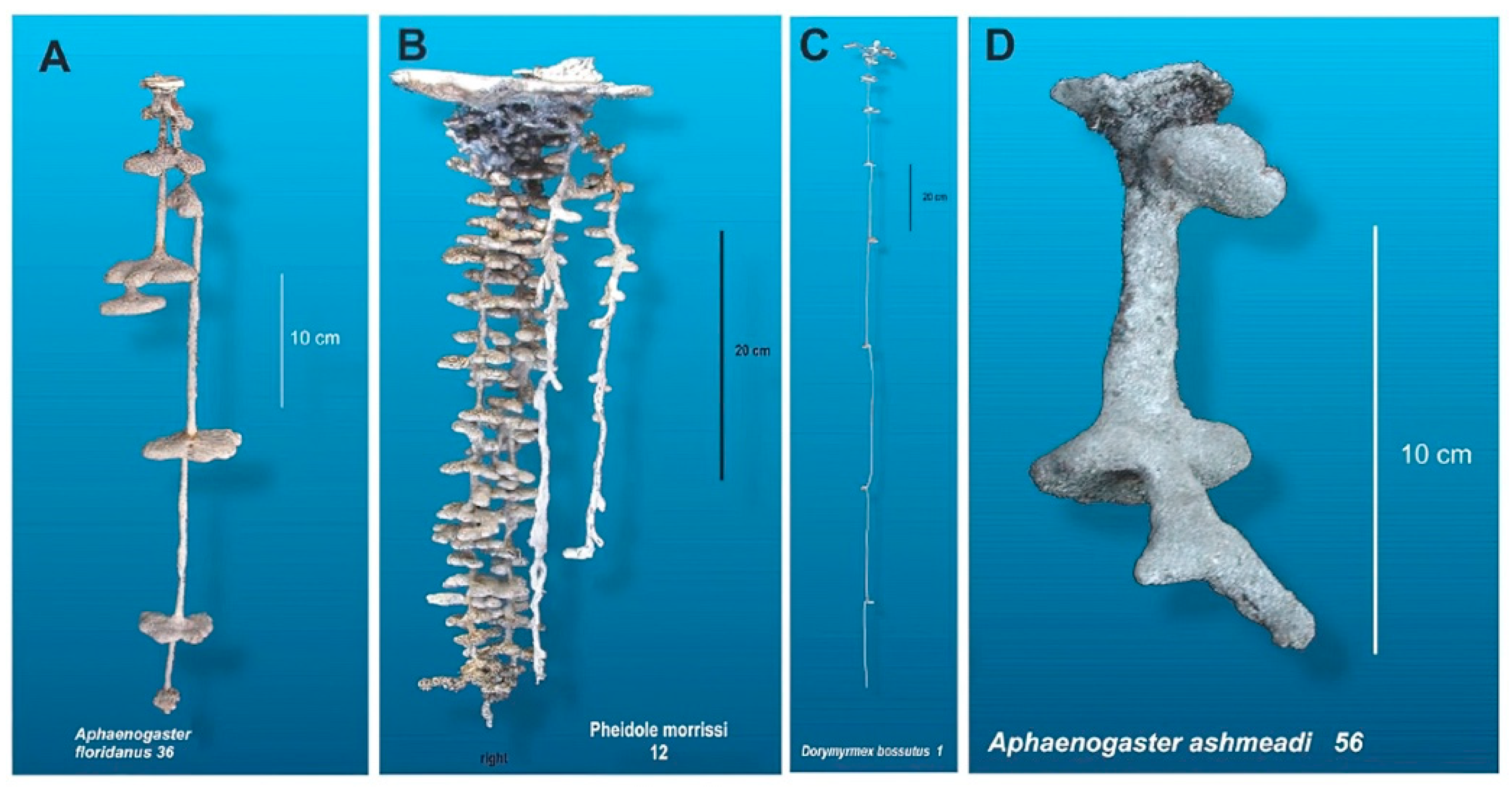
2.1.1. Research Methodology
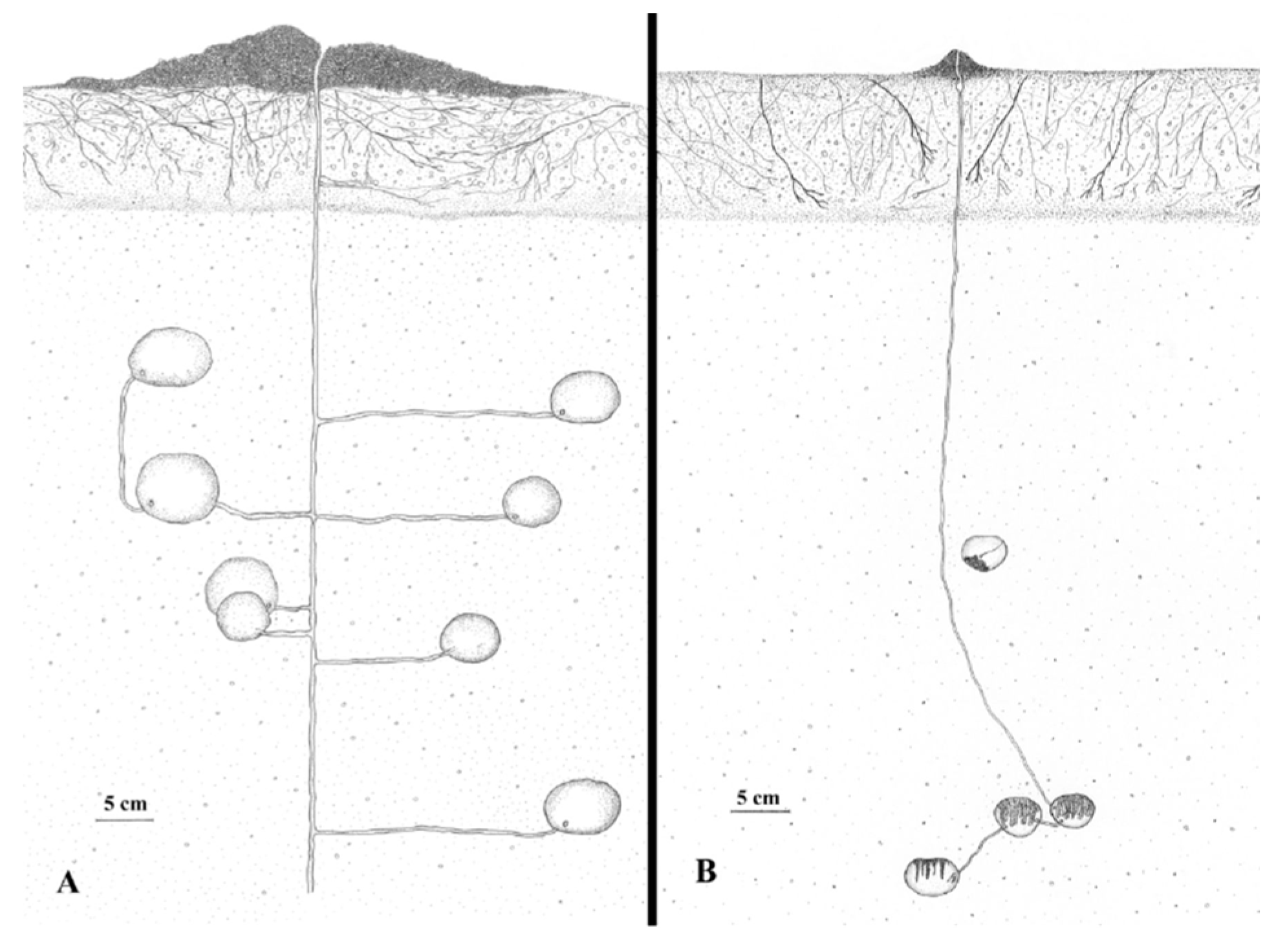
2.1.2. The Interaction between Structure and Individual
2.2. Constructional Bionics of Underground Ant Nest Structures
3. Architectural Perspective
3.1. Characteristics of Underground Ant Nest Structures
3.2. Internal Environmental Characteristics of Underground Ant Nest
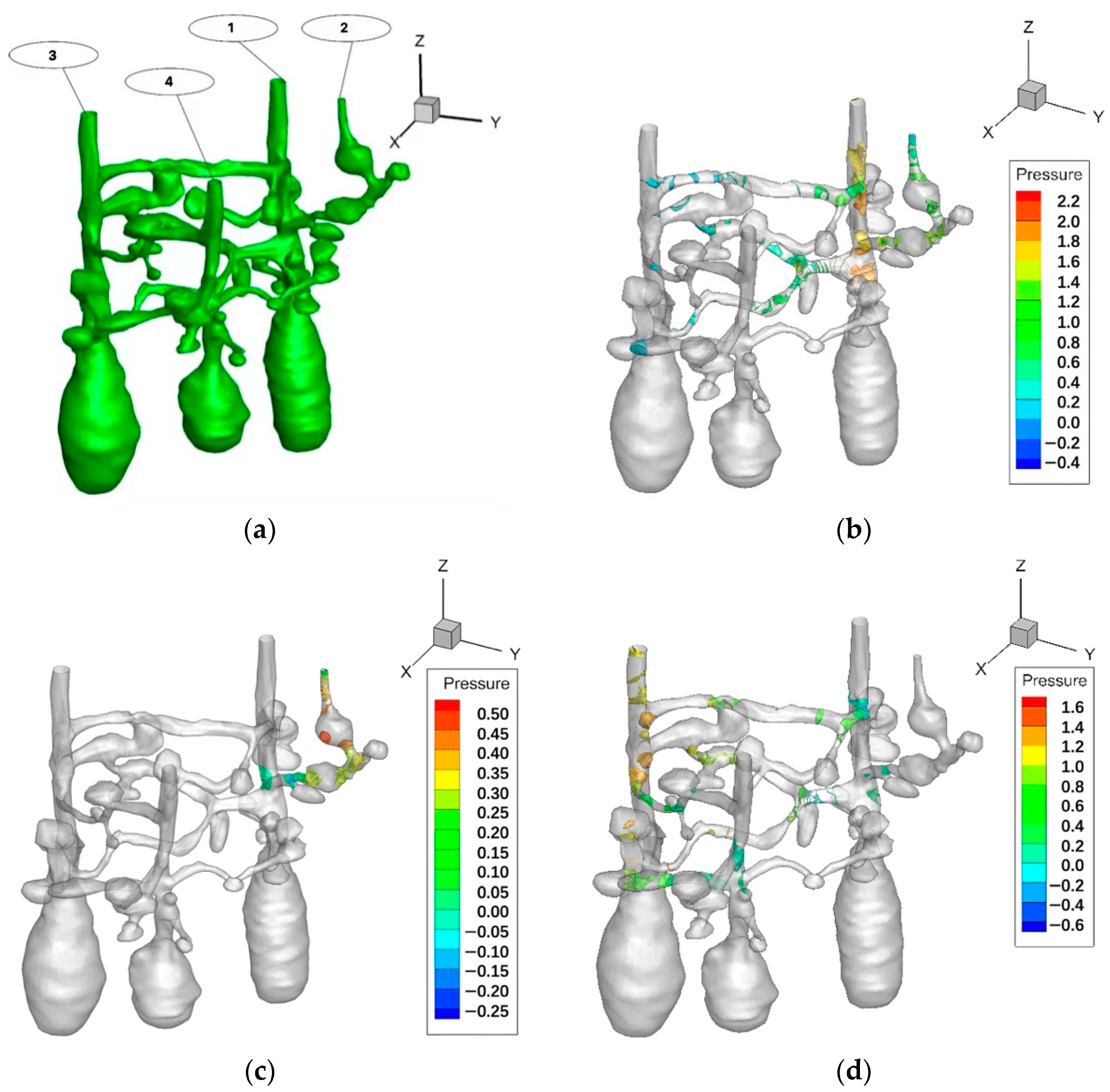
3.3. Ventilation Analysis of the Ant Nest
3.4. Morphological Bionics of Underground Ant Nest Structures
3.5. Functional Bionics of Underground Ant Nest Structures
4. Civil Engineering Perspective
4.1. The Effect of Underground Ant Nest on Soil Geotechnical Properties
4.1.1. Physical Properties
4.1.2. Hydraulic Properties
4.2. CANCP
4.3. Material Bionics of Underground Ant Nest Structures
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ben-Alon, L.; Sacks, R.; Grobman, Y.J. Similarities and Differences between Humans’ and Social Insects’ Building Processes and Building Behaviors. In Proceedings of the Construction Research Congress 2014: Construction in a Global Network, Atlanta, GA, USA, 19–21 May 2014; pp. 51–60. [Google Scholar]
- Camazine, S.; Deneubourg, J.-L.; Franks, N.R.; Sneyd, J.; Theraula, G.; Bonabeau, E. Self-Organization in Biological Systems; Princeton University Press: Princeton, NJ, USA, 2020. [Google Scholar]
- Detrain, C.; Deneubourg, J.-L. Self-organized structures in a superorganism: Do ants “behave” like molecules? Phys. Life Rev. 2006, 3, 162–187. [Google Scholar] [CrossRef]
- Anderson, C.; Theraulaz, G.; Deneubourg, J.-L. Self-assemblages in insect societies. Insectes Sociaux 2002, 49, 99–110. [Google Scholar] [CrossRef]
- Holbrook, C.T.; Clark, R.M.; Moore, D.; Overson, R.P.; Penick, C.A.; Smith, A.A. Social insects inspire human design. Biol. Lett. 2010, 6, 431–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabin, J.E.; Jones, P.L. Labstudio: Design Research between Architecture and Biology; Routledge: London, UK, 2017. [Google Scholar]
- Mertins, D. Where architecture meets biology: An interview with Detlef Mertins. In Departmental Papers (Architecture); University of Pennsylvania: Philadelphia, PA, USA, 2007; p. 7. [Google Scholar]
- Tim, I.; Simon, G. Architecture, space and information in constructions built by humans and social insects: A conceptual review. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2018, 373, 20170244. [Google Scholar]
- Tschinkel, W.R. The nest architecture of the ant, Camponotus socius. J. Insect Sci. 2005, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Tschinkel, W.R. The nest architecture of the Florida harvester ant, Pogonomyrmex badius. J. Insect Sci. 2004, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Moreira, A.; Forti, L.; Boaretto, M.; Andrade, A.; Lopes, J.; Ramos, V. External and internal structure of Atta bisphaerica Forel (Hymenoptera: Formicidae) nests. J. Appl. Entomol. 2004, 128, 204–211. [Google Scholar] [CrossRef]
- Tschinkel, W.R. Subterranean ant nests: Trace fossils past and future? Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 192, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Tschinkel, W.R. Sociometry and sociogenesis of colonies of the harvester ant, Pogonomyrmex badius: Worker characteristics in relation to colony size and season. Insectes Sociaux 1998, 45, 385–410. [Google Scholar] [CrossRef]
- Tschinkel, W.R. Sociometry and sociogenesis of colonies of the harvester ant, Pogonomyrmex badius: Distribution of workers, brood and seeds within the nest in relation to colony size and season. Ecol. Entomol. 1999, 24, 222–237. [Google Scholar] [CrossRef]
- Williams, D.F.; Lofgren, C.S.; Trager, J. Nest casting of some grounddwelling Florida ant species using dental labstone. In Advances in Myrmecology; Brill Publishers: Leiden, The Netherlands, 1988; Volume 433. [Google Scholar]
- Tschinkel, W.R. Ant Architecture: The Wonder, Beauty, and Science of Underground Nests; Princeton University Press: Princeton, NJ, USA, 2021. [Google Scholar]
- Tschinkel, W.R. Methods for casting subterranean ant nests. J. Insect Sci. 2010, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Pinter-Wollman, N.; Penn, A.; Theraulaz, G.; Fiore, S.M. Interdisciplinary approaches for uncovering the impacts of architecture on collective behaviour. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mersch, D.P.; Crespi, A.; Keller, L. Tracking individuals shows spatial fidelity is a key regulator of ant social organization. Science 2013, 340, 1090–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prabhakar, B.; Dektar, K.N.; Gordon, D.M. The regulation of ant colony foraging activity without spatial information. PLoS Comput. Biol. 2012, 8, e1002670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinter-Wollman, N. Nest architecture shapes the collective behaviour of harvester ants. Biol. Lett. 2015, 11, 20150695. [Google Scholar] [CrossRef] [Green Version]
- Gordon, D.M. Ant encounters. In Ant Encounters; Princeton University Press: Princeton, NJ, USA, 2010. [Google Scholar]
- Pinter-Wollman, N.; Bala, A.; Merrell, A.; Queirolo, J.; Stumpe, M.C.; Holmes, S.; Gordon, D.M. Harvester ants use interactions to regulate forager activation and availability. Anim. Behav. 2013, 86, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Pinter-Wollman, N.; Wollman, R.; Guetz, A.; Holmes, S.; Gordon, D.M. The effect of individual variation on the structure and function of interaction networks in harvester ants. J. R. Soc. Interface 2011, 8, 1562–1573. [Google Scholar] [CrossRef]
- Korb, J. Thermoregulation and ventilation of termite mounds. Naturwissenschaften 2003, 90, 212–219. [Google Scholar] [CrossRef]
- Grassé, P.-P. La reconstruction du nid et les coordinations interindividuelles chezBellicositermes natalensis etCubitermes sp. la théorie de la stigmergie: Essai d’interprétation du comportement des termites constructeurs. Insectes Sociaux 1959, 6, 41–80. [Google Scholar] [CrossRef]
- Tschinkel, W.R. Nest relocation and excavation in the Florida harvester ant, Pogonomyrmex badius. PLoS ONE 2014, 9, e112981. [Google Scholar] [CrossRef]
- Khuong, A.; Gautrais, J.; Perna, A.; Sbaï, C.; Combe, M.; Kuntz, P.; Jost, C.; Theraulaz, G. Stigmergic construction and topochemical information shape ant nest architecture. Proc. Natl. Acad. Sci. USA 2016, 113, 1303–1308. [Google Scholar] [CrossRef] [PubMed]
- Rabeling, C.; Verhaagh, M.; Engels, W. Comparative study of nest architecture and colony structure of the fungus-growing ants, Mycocepurus goeldii and M. smithii. J. Insect Sci. 2007, 7, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tschinkel, W.R. The nest architecture of three species of north Florida Aphaenogaster ants. J. Insect Sci. 2011, 11, 105. [Google Scholar] [CrossRef] [Green Version]
- Verza, S.; Forti, L.C.; Lopes, J.; Hughes, W. Nest architecture of the leaf-cutting ant Acromyrmex rugosus rugosus. Insectes Sociaux 2007, 54, 303–309. [Google Scholar] [CrossRef]
- Mikheyev, A.S.; Tschinkel, W. Nest architecture of the ant Formica pallidefulva: Structure, costs and rules of excavation. Insectes Sociaux 2004, 51, 30–36. [Google Scholar] [CrossRef]
- Cerquera, L.M.; Tschinkel, W.R. The nest architecture of the ant Odontomachus brunneus. J. Insect Sci. 2010, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forti, L.C.; Protti de Andrade, A.P.; Camargo, R.d.S.; Caldato, N.; Moreira, A.A. Discovering the giant nest architecture of grass-cutting ants, Atta capiguara (Hymenoptera, Formicidae). Insects 2017, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Li, D.; Dong, G.; Zhao, Y.F. Crack path-engineered 2D octet-truss lattice with bio-inspired crack deflection. Addit. Manuf. 2020, 36, 101539. [Google Scholar] [CrossRef]
- Tiwary, C.S.; Kishore, S.; Sarkar, S.; Mahapatra, D.R.; Ajayan, P.M.; Chattopadhyay, K. Morphogenesis and mechanostabilization of complex natural and 3D printed shapes. Sci. Adv. 2015, 1, e1400052. [Google Scholar] [CrossRef] [Green Version]
- Ambekar, R.S.; Oliveira, E.F.; Kushwaha, B.; Pal, V.; Ajayan, P.M.; Roy, A.K.; Galvao, D.S.; Tiwary, C.S. Flexure resistant 3D printed zeolite-inspired structures. Addit. Manuf. 2021, 47, 102297. [Google Scholar] [CrossRef]
- Liu, J.; Gaynor, A.T.; Chen, S.; Kang, Z.; Suresh, K.; Takezawa, A.; Li, L.; Kato, J.; Tang, J.; Wang, C.C. Current and future trends in topology optimization for additive manufacturing. Struct. Multidiscip. Optim. 2018, 57, 2457–2483. [Google Scholar] [CrossRef]
- Kushwaha, B.; Kumar, A.; Ambekar, R.S.; Arya, V.; Negedu, S.D.; Bakshi, D.; Olu, F.E.; Sastri Ayyagari, R.; Pal, V.; Sadasivuni, K.K. Understanding the mechanics of complex topology of the 3D printed Anthill architecture. Oxf. Open Mater. Sci. 2022, 2, itac003. [Google Scholar] [CrossRef]
- Jones, J.C.; Oldroyd, B.P. Nest thermoregulation in social insects. Adv. Insect Physiol. 2006, 33, 153–191. [Google Scholar]
- Kleineidam, C.; Ernst, R.; Roces, F. Wind-induced ventilation of the giant nests of the leaf-cutting ant Atta vollenweideri. Naturwissenschaften 2001, 88, 301–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollazzi, M.; Roces, F. Control of nest water losses through building behavior in leaf-cutting ants (Acromyrmex heyeri). Insectes Sociaux 2010, 57, 267–273. [Google Scholar] [CrossRef]
- Bollazzi, M.; Roces, F. Leaf-cutting ant workers (Acromyrmex heyeri) trade off nest thermoregulation for humidity control. J. Ethol. 2010, 28, 399–403. [Google Scholar] [CrossRef]
- Anderson, K.E.; Munger, J.C. Effect of temperature on brood relocation in Pogonomyrmex salinus (Hymenoptera: Formicidae). West. N. Am. Nat. 2003, 63, 122–128. [Google Scholar]
- Bollazzi, M.; Roces, F. Thermal preference for fungus culturing and brood location by workers of the thatching grass-cutting ant Acromyrmex heyeri. Insectes Sociaux 2002, 49, 153–157. [Google Scholar] [CrossRef]
- The Global Health Community (Termite’s Nest-Based Apartment Complex Design) Guangzhou, China. World Archit. Rev. 2010, 25, 92–97.
- Drager, K.I.; Hirmas, D.R.; Hasiotis, S.T. Effects of Ant (Formica subsericea) Nests on Physical and Hydrological Properties of a Fine-Textured Soil. Soil Sci. Soc. Am. J. 2016, 80, 364–375. [Google Scholar] [CrossRef]
- Jiang, J.; Luo, G. Experimental investigations on correlativity between liquidity index and other parameters of silty clay in Su-Tong Bridge subgrade. Rock Soil Mech. 2008, 29, 1669–1674. [Google Scholar]
- Brian, M.V.; Brian, M. Production Ecology of Ants and Termites; Cambridge University Press: Cambridge, UK, 1978; Volume 13. [Google Scholar]
- Jungerius, P.D.; Ancker, J.A.M.v.d.; Mücher, H.J. The contribution of termites to the microgranular structure of soils on the Uasin Gishu Plateau, Kenya. Catena 1999, 34, 349–363. [Google Scholar] [CrossRef]
- Majer, J.; Walker, T.; Berlandier, F. The role of ants in degraded soils within Dryandra state forest. Mulga Res. J. 1987, 9, 15–16. [Google Scholar]
- Baxter, F.P.; Hole, F.D. Ant (Formica cinerea) pedoturbation in a prairie soil. Soil Sci. Soc. Am. J. 1967, 31, 425–428. [Google Scholar] [CrossRef]
- McGarry, D.; Bridge, B.; Radford, B. Contrasting soil physical properties after zero and traditional tillage of an alluvial soil in the semi-arid subtropics. Soil Tillage Res. 2000, 53, 105–115. [Google Scholar] [CrossRef]
- Mandel, R.D.; Sorenson, C.J. The role of the western harvester ant (Pogonomyrmex occidentalis) in soil formation. Soil Sci. Soc. Am. J. 1982, 46, 785–788. [Google Scholar] [CrossRef]
- Dauber, J.; Rommeler, A.; Wolters, V. The ant Lasius flavus alters the viable seed bank in pastures. Eur. J. Soil Biol. 2006, 42, S157–S163. [Google Scholar] [CrossRef]
- Jílková, V.; Šebek, O.; Frouz, J. Mechanisms of pH change in wood ant (Formica polyctena) nests. Pedobiologia 2012, 55, 247–251. [Google Scholar] [CrossRef]
- Geiger, S.; Manu, A.; Bationo, A. Changes in a sandy Sahelian soil following crop residue and fertilizer additions. Soil Sci. Soc. Am. J. 1992, 56, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Garnier-Sillam, E.; Toutain, F. Distribution of polysaccharides within the humic compounds of soils subjected to a humivorous termite Thoracotermes macrothorax Sjostedt. Pedobiologia 1995, 39, 462–469. [Google Scholar]
- Jílková, V.; Frouz, J. Contribution of ant and microbial respiration to CO2 emission from wood ant (Formica polyctena) nests. Eur. J. Soil Biol. 2014, 60, 44–48. [Google Scholar] [CrossRef]
- Abbadie, L.; Lepage, M. The role of subterranean fungus comb chambers (Isoptera, Macrotermitinae) in soil nitrogen cycling in a preforest savanna (Cote dIvoire). Soil Biol. Biochem. 1989, 21, 1067–1071. [Google Scholar] [CrossRef]
- Lilburn, T.; Kim, K.; Ostrom, N.; Byzek, K.; Leadbetter, J.; Breznak, J. Nitrogen fixation by symbiotic and free-living spirochetes. Science 2001, 292, 2495–2498. [Google Scholar] [CrossRef] [PubMed]
- Elinwa, A. Experimental characterization of Portland cement-calcined soldier-ant mound clay cement mortar and concrete. Constr. Build. Mater. 2006, 20, 754–760. [Google Scholar] [CrossRef]
- Barger, G.S.; Hansen, E.R.; Wood, M.R.; Neary, T.; Beech, D.J.; Jaquier, D. Production and use of calcined natural pozzolans in concrete. Cem. Concr. Aggreg. 2001, 23, 73–80. [Google Scholar]
- Tironi, A.; Trezza, M.A.; Scian, A.N.; Irassar, E.F. Assessment of pozzolanic activity of different calcined clays. Cem. Concr. Compos. 2013, 37, 319–327. [Google Scholar] [CrossRef]
- Shao, Y.; Lefort, T.; Moras, S.; Rodriguez, D. Studies on concrete containing ground waste glass. Cem. Concr. Res. 2000, 30, 91–100. [Google Scholar] [CrossRef]
- Ay, N.; Ünal, M. The use of waste ceramic tile in cement production. Cem. Concr. Res. 2000, 30, 497–499. [Google Scholar] [CrossRef]
- O’Farrell, M.; Wild, S.; Sabir, B. Pore size distribution and compressive strength of waste clay brick mortar. Cem. Concr. Compos. 2001, 23, 81–91. [Google Scholar] [CrossRef]
- Mohammed, S. Processing, effect and reactivity assessment of artificial pozzolans obtained from clays and clay wastes: A review. Constr. Build. Mater. 2017, 140, 10–19. [Google Scholar] [CrossRef]
- Zhou, W.; Zhu, P.; Qu, W. Basic Properties of calcined underground ant nest materials and its influence on the compressive strength of concrete. Materials 2019, 12, 1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Xu, J. Application of microbial precipitation in self-healing concrete: A review on the protection strategies for bacteria. Constr. Build. Mater. 2021, 306, 124950. [Google Scholar] [CrossRef]
- Zhou, W.; Zhu, P.; Qu, W.; Yao, W.; Wu, S. Study on the influence of calcined underground ant nest powder on the durability of concrete. Materials 2020, 13, 2119. [Google Scholar] [CrossRef]
- Jiang, Y. Out of the Eastgate. Build. Sci. 2011, 27, 1–5. [Google Scholar]
- Zhao, J.; Xu, Y. Ecological Wisdom Inspired from Termite Mounds—Analysis on Biometric Design of Zimbabwe Eastgate Center. Build. Sci. 2010, 26, 19–23. [Google Scholar]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.; Zhou, W.; Qu, W.; Yao, W.; Zhu, P.; Xu, J. A Review of Ant Nests and Their Implications for Architecture. Buildings 2022, 12, 2225. https://doi.org/10.3390/buildings12122225
Yang G, Zhou W, Qu W, Yao W, Zhu P, Xu J. A Review of Ant Nests and Their Implications for Architecture. Buildings. 2022; 12(12):2225. https://doi.org/10.3390/buildings12122225
Chicago/Turabian StyleYang, Guanghong, Wei Zhou, Wenjun Qu, Wu Yao, Peng Zhu, and Jing Xu. 2022. "A Review of Ant Nests and Their Implications for Architecture" Buildings 12, no. 12: 2225. https://doi.org/10.3390/buildings12122225