
Predicting the Current and Future Distributions of Frankliniella occidentalis (Pergande) Based on the MaxEnt Species Distribution Model

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data
2.1.1. Species Distribution Data
2.1.2. Bioclimatic Variables
2.2. Data Analysis
Bioclimatic Variable Selection
2.3. Evaluation Criteria
3. Results
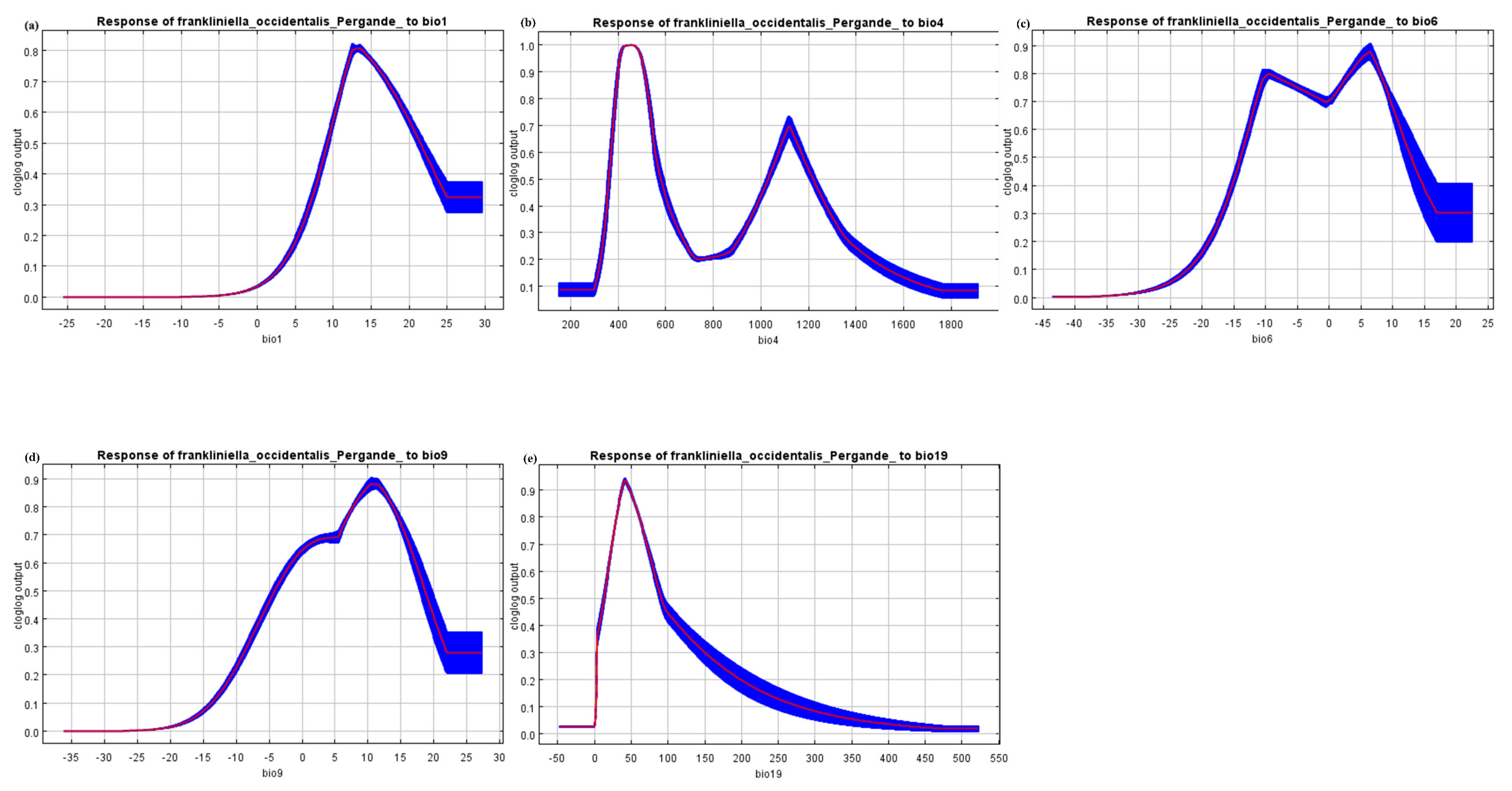
3.1. Key Environment Variable Selection and Model Performance
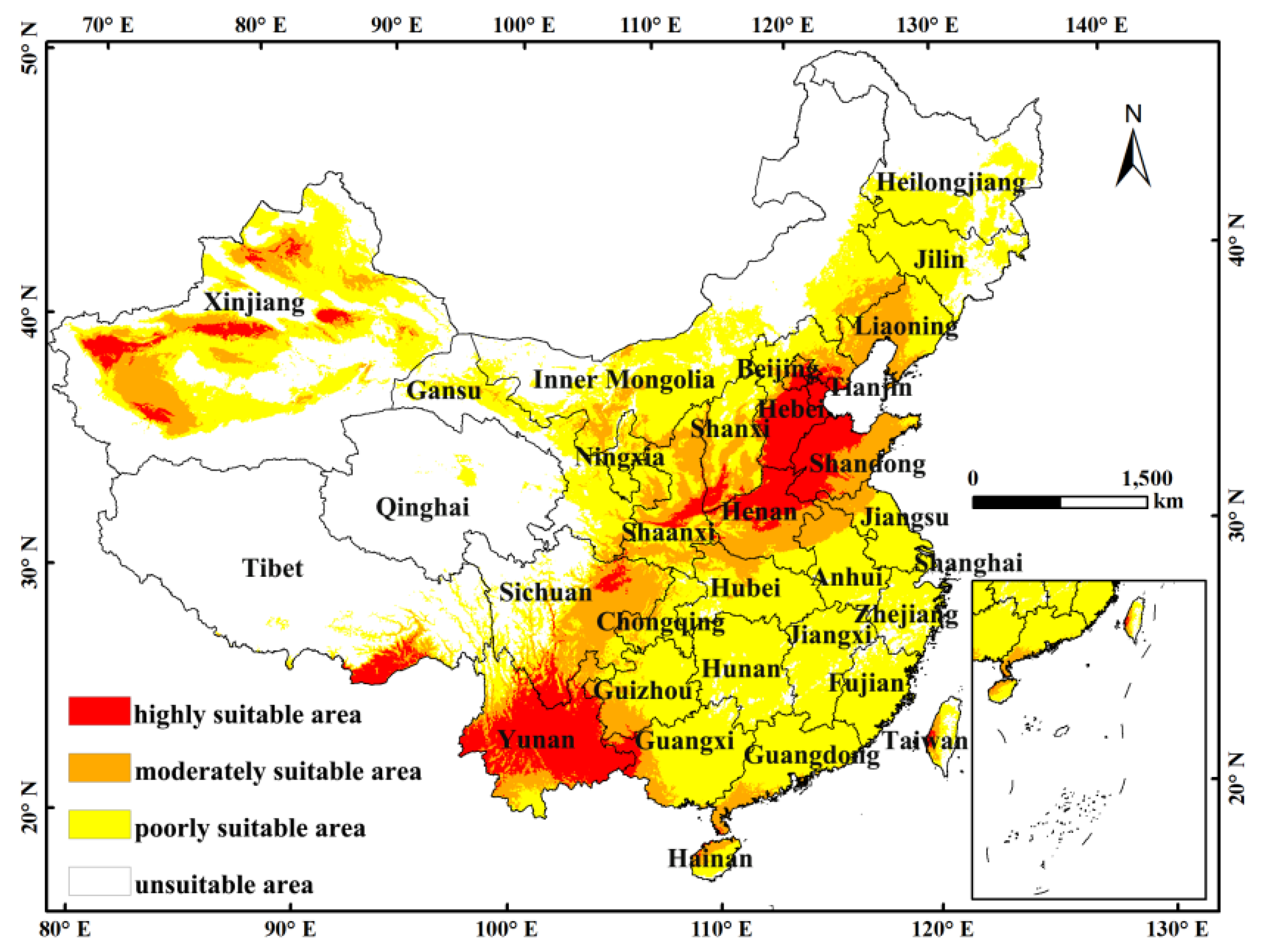
3.2. Species Response and Potential Habitat Suitability Distribution
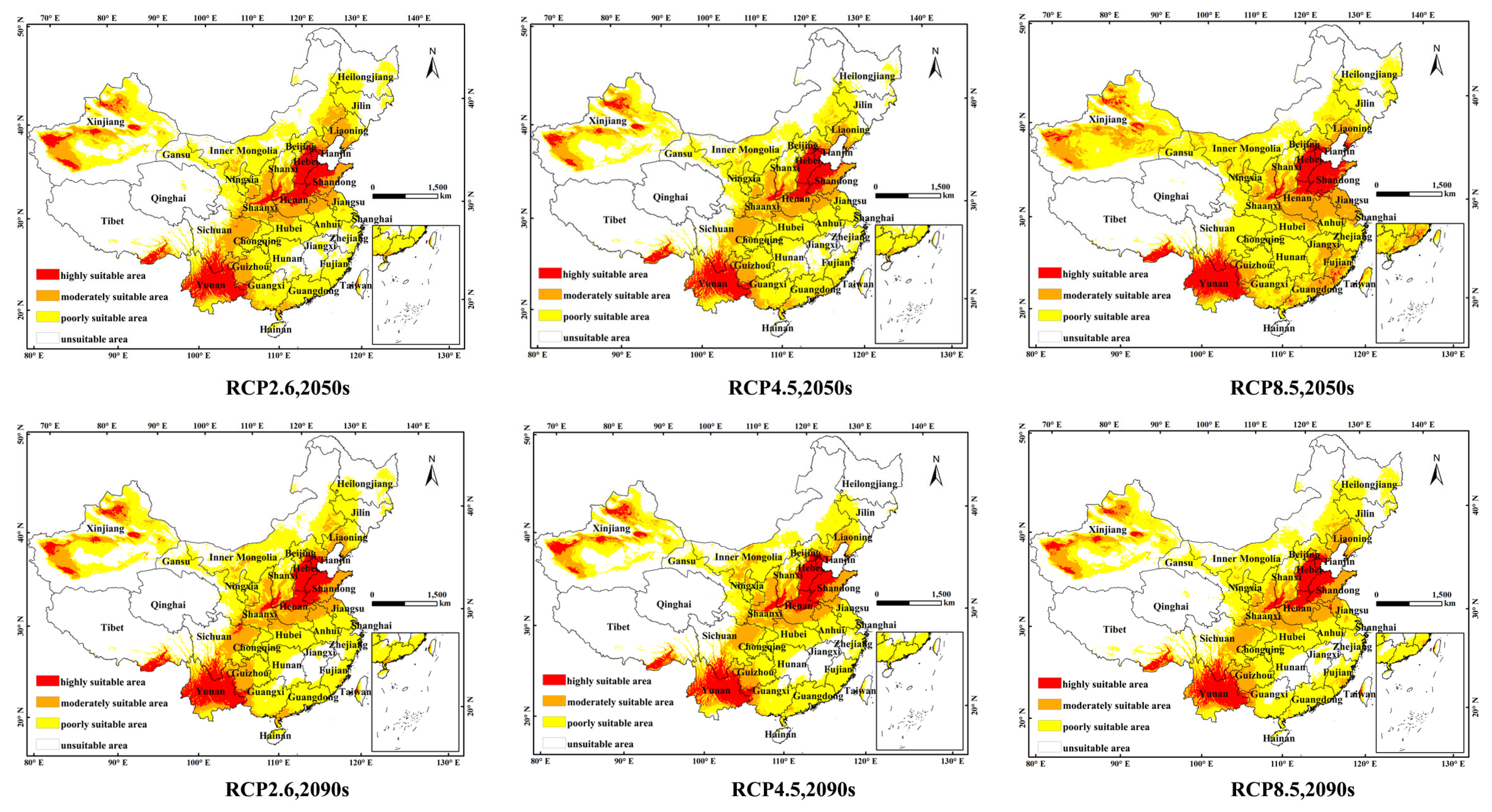
3.3. Environmental Adaptation under Current and Future Climate Scenarios
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sampson, C.; Bennison, J.; Kirk, W.D.J. Overwintering of the western flower thrips in outdoor strawberry crops. J. Pest Sci. 2021, 94, 143–152. [Google Scholar] [CrossRef]
- Dlamini, T.M.; Allsopp, E.; Malan, A.P. Application of Steinernema yirgalemense to control Frankliniella occidentalis (Thysanoptera: Thripidae) on blueberries. Crop Prot. 2020, 128, 105016. [Google Scholar] [CrossRef]
- Li, W.D.; Zhang, P.J.; Zhang, J.M.; Zhang, Z.J.; Huang, F.; Bei, Y.W.; Lin, W.C.; Lu, Y.B. An evaluation of Frankliniella occidentalis (Thysanoptera: Thripidae) and Frankliniella intonsa (Thysanoptera: Thripidae) performance on different plant leaves based on life history characteristics. J. Insect Sci. 2015, 15, 167. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Qian, W.; Qiao, X.; Xi, Y.; Wan, F. Invasion biology, ecology, and management of Frankliniella occidentalis in China. Arch. Insect Biochem. 2019, 102, e21613. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Yuan, J.; Wang, J.; Hua, D.; Zheng, X.; Tao, M.; Zhang, Z.; Wan, Y.; Wang, S.; Zhang, Y.; et al. Susceptibility levels of field populations of Frankliniella occidentalis (Thysanoptera: Thripidae) to seven insecticides in China. Crop Prot. 2022, 153, 105886. [Google Scholar] [CrossRef]
- Duan, H.; Yu, Y.; Zhang, A.; Guo, D.; Tao, Y.; Chu, D. Sudden Widespread Distribution of Frankliniella occidentalis (Thysanoptera: Thripidae) in Shandong Province, China. Fla. Entomol. 2013, 96, 933–940. [Google Scholar] [CrossRef]
- Garrick, T.A.; Liburd, O.E.; Funderburk, J. Effect of Humidity on Fecundity and Egg Incubation of Frankliniella bispinosa and Frankliniella occidentalis (Thysanoptera: Thripidae). Fla. Entomol. 2016, 99, 505–508. [Google Scholar] [CrossRef]
- Demirozer, O.; Tyler-Julian, K.; Funderburk, J.; Leppla, N.; Reitz, S. Frankliniella occidentalis (Pergande) integrated pest management programs for fruiting vegetables in Florida. Pest Manag. Sci. 2012, 68, 1537–1545. [Google Scholar] [CrossRef]
- Park, Y.; Lee, J. Life history characteristics of the western flower thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae), under fluctuating conditions of temperature or relative humidity. J. Asia-Pac. Entomol. 2020, 23, 606–611. [Google Scholar] [CrossRef]
- Zhang, W.; Jiao, Y.; Ding, C.; Shen, L.; Li, Y.; Yu, Y.; Huang, K.; Li, B.; Wang, F.; Yang, J. Rapid Detection of Tomato Spotted Wilt Virus With Cas13a in Tomato and Frankliniella occidentalis. Front. Microbiol. 2021, 12, 3118. [Google Scholar] [CrossRef]
- Luna-Espino, H.M.; Jiménez-Pérez, A.; Castrejón-Gómez, V.R. Assessment of Chrysoperla comanche (Banks) and Chrysoperla externa (Hagen) as Biological Control Agents of Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae) on Tomato (Solanum lycopersicum) under Glasshouse Conditions. Insects 2020, 11, 87. [Google Scholar] [CrossRef] [PubMed]
- Ferrand, L.; Almeida, M.M.S.; Orílio, A.F.; Dal Bó, E.; Resende, R.O.; García, M.L. Biological and molecular characterization of tomato spotted wilt virus (TSWV) resistance-breaking isolates from Argentina. Plant Pathol. 2019, 68, 1587–1601. [Google Scholar] [CrossRef]
- Batuman, O.; Turini, T.A.; LeStrange, M.; Stoddard, S.; Miyao, G.; Aegerter, B.J.; Chen, L.; McRoberts, N.; Ullman, D.E.; Gilbertson, R.L. Development of an IPM Strategy for Thrips and Tomato spotted wilt virus in Processing Tomatoes in the Central Valley of California. Pathogens 2020, 9, 636. [Google Scholar] [CrossRef] [PubMed]
- Ogada, P.A.; Kiirika, L.M.; Lorenz, C.; Senkler, J.; Braun, H.; Poehling, H. Differential proteomics analysis of Frankliniella occidentalis immune response after infection with Tomato spotted wilt virus (Tospovirus). Dev. Comp. Immunol. 2017, 67, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Funderburk, J.; Frantz, G.; Mellinger, C.; Tyler-Julian, K.; Srivastava, M. Biotic resistance limits the invasiveness of the western flower thrips, Frankliniella occidentalis (Thysanoptera: Thripidae), in Florida. Insect Sci. 2016, 23, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Bystrova, D.; Poggiato, G.; Bektaş, B.; Arbel, J.; Clark, J.S.; Guglielmi, A.; Thuiller, W. Clustering Species With Residual Covariance Matrix in Joint Species Distribution Models. Front. Ecol. Evol. 2021, 9, 601384. [Google Scholar] [CrossRef]
- Yu, H.; Cooper, A.R.; Infante, D.M. Improving species distribution model predictive accuracy using species abundance: Application with boosted regression trees. Ecol. Model. 2020, 432, 109202. [Google Scholar] [CrossRef]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. Bioclim: The first species distribution modelling package, its early applications and relevance to most currentMaxEnt studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef]
- Waldock, C.; Smith, R.D.S.; Albouy, C.; Cheung, W.W.L.; Edgar, G.J.; Mouillot, D.; Tjiputra, J.; Pellissier, L. A quantitative review of abundance-based species distribution models. Ecography 2021, 2022. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M.; et al. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- West, A.M.; Kumar, S.; Brown, C.S.; Stohlgren, T.J.; Bromberg, J. Field validation of an invasive species Maxent model. Ecol. Inform. 2016, 36, 126–134. [Google Scholar] [CrossRef]
- He, Z.; Guo, J.F.; Reitz, S.R.; Lei, Z.R.; Wu, S.Y. A global invasion by the thrip, Frankliniella occidentalis: Current virus vector status and its management. Insect Sci. 2020, 27, 626–645. [Google Scholar] [CrossRef]
- Santos, A.A.; Hogendoorn, K.; Ramos, R.S.; Picanco, M.C. Distribution models for Ascia monuste and the host Brassica oleracea var. capitata. J. Appl. Entomol. 2019, 143, 1043–1051. [Google Scholar] [CrossRef]
- Merow, C.; Silander, J.A. A comparison of Maxlike and Maxent for modelling species distributions. Methods Ecol. Evol. 2014, 5, 215–225. [Google Scholar] [CrossRef]
- Wittmann, M.E.; Barnes, M.A.; Jerde, C.L.; Jones, L.A.; Lodge, D.M. Confronting species distribution model predictions with species functional traits. Ecol. Evol. 2016, 6, 873–879. [Google Scholar] [CrossRef]
- Jian, C.W.; Bin, Z.; Hong, G.L.; Jun, P.W.; Chang, Y.Z. Effects of Exposure to High Temperature on Frankliniella occidentalis (Thysanoptera: Thripidae), under Arrhenotoky and Sexual Reproduction Conditions. Fla. Entomol. 2014, 97, 504–510. [Google Scholar]
- Fatnassi, H.; Pizzol, J.; Senoussi, R.; Biondi, A.; Desneux, N.; Poncet, C.; Boulard, T. Within-Crop Air Temperature and Humidity Outcomes on Spatio-Temporal Distribution of the Key Rose Pest Frankliniella occidentalis. PLoS ONE 2015, 10, e126655. [Google Scholar] [CrossRef]
- Ullah, M.S.; Lim, U.T. Life History Characteristics of Frankliniella occidentalis and Frankliniella intonsa (Thysanoptera: Thripidae) in Constant and Fluctuating Temperatures. J. Econ. Entomol. 2015, 108, 1000–1009. [Google Scholar] [CrossRef]
- Cao, Y.; Yang, H.; Li, J.; Zhang, G.; Wang, Y.; Li, C.; Gao, Y. Population development of Frankliniella occidentalis and Thrips hawaiiensis in constant and fluctuating temperatures. J. Appl. Entomol. 2019, 143, 49–57. [Google Scholar] [CrossRef]
- ShuQi, H.; Ying, L.; Lei, Q.; ZhiHua, L.; Chao, X.; Lu, Y.; FuRong, G. The Influence of Elevated CO2 Concentration on the Fitness Traits of Frankliniella occidentalis and Frankliniella intonsa (Thysanoptera: Thripidae). Environ. Entomol. 2017, 46, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Chen, F.; Liu, J.; He, S.; Liu, J.; Li, Z.; Gui, F. Effects of elevated CO2 on life-history traits of three successive generations of Frankliniella occidentalis andF. intonsa on kidney bean, Phaseolus vulgaris. Entomol. Exp. Appl. 2017, 165, 50–61. [Google Scholar] [CrossRef]
- Nielsen, M.C.; Teulon, D.A.J.; Chapman, R.B.; Butler, R.C.; Drayton, G.M.; Phillipsen, H. Effects of temperature on survival, oviposition, and development rate of ‘greenhouse’ and ‘lupin’ strains of western flower thrips, Frankliniella occidentalis. Entomol. Exp. Appl. 2021, 169, 480–490. [Google Scholar] [CrossRef]
- Wang, Z.; Gong, Y.; Jin, G.; Li, B.; Chen, J.; Kang, Z.; Zhu, L.; Gao, Y.; Reitz, S.; Wei, S. Field-evolved resistance to insecticides in the invasive western flower thrips Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae) in China. Pest Manag. Sci. 2016, 72, 1440–1444. [Google Scholar] [CrossRef]
- Chu, Z.; Huan, Z. Research progress of Frankliniella occidentalis (Pergande) control. Mod. Agric. 2022, 7–9. [Google Scholar] [CrossRef]
- Zengzhi, L. History, Progress and Current Status of the Application of Fungi against Pest Insects in China. Chin. J. Biol. Control. 2015, 31, 699–711. [Google Scholar]
- Mei, L.; Changrong, Z.; Xiaoli, S.; Jun, G.; Guang, Z.; Lejuan, S. Control of Frankliniella occidentalis on Gerbera jamesoni by Orius similis. Chin. J. Biol. Control 2020, 36, 992–996. [Google Scholar]
- Songsong, P.; Xuan, W.; Ruijun, L.; Dan, Z.; Xiujun, L.; Wei, G. Screening of Efficient Metarhizium anisopliae Strains for Frankliniella occidentalis and Its Application in Peanut Field. Chin. J. Biol. Control 2021, 37, 732–739. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Bio 1 | Bio 4 | Bio 6 | Bio 9 |
|---|---|---|---|---|
| Bio 4 | 0.704 ** | |||
| Bio 6 | −0.076 | −0.755 ** | ||
| Bio 9 | 0.575 ** | −0.139 * | 0.712 ** | |
| Bio 19 | 0.729 ** | 0.356 ** | 0.201 ** | 0.605 ** |
| Province | High-Suitable Area (104 km2) | Total (104 km2) * | Percentage of High-Suitable Area in the Province (%) | Percentage of High-Suitable Area in China (%) |
|---|---|---|---|---|
| Tianjin | 1.18 | 1.20 | 98.3 | 1.8 |
| Shandong | 8.35 | 15.58 | 53.6 | 12.6 |
| Beijing | 0.84 | 1.64 | 51.2 | 1.3 |
| Hebei | 9.27 | 18.88 | 49.1 | 14.0 |
| Henan | 6.40 | 16.70 | 38.3 | 9.7 |
| Shanxi | 1.40 | 15.67 | 8.9 | 2.1 |
| Shaanxi | 1.61 | 20.56 | 7.8 | 2.4 |
| Sichuan | 3.65 | 48.60 | 7.5 | 5.5 |
| Taiwan | 0.20 | 3.60 | 5.6 | 0.3 |
| Yunnan | 2.13 | 39.41 | 5.4 | 3.2 |
| Guangxi | 1.20 | 23.76 | 5.1 | 1.8 |
| Xinjiang | 7.16 | 166.49 | 4.3 | 10.8 |
| Hainan | 0.09 | 3.54 | 2.5 | 0.1 |
| Tibet | 3.05 | 122.84 | 2.5 | 4.6 |
| Guizhou | 0.36 | 17.62 | 2.1 | 0.5 |
| Jiangsu | 0.19 | 10.72 | 1.8 | 0.3 |
| Guangdong | 0.06 | 17.97 | 0.3 | 0.1 |
| Anhui | 0.02 | 14.01 | 0.1 | 0.0 |
| Gansu | 0.003 | 42.58 | 0.0 | 0.0 |
| China | 47.16 | / | / | 0.1 |
| Decade | Scenarios | Predicted Area (104 km2) | Comparison with Current Distribution (%) | ||||
|---|---|---|---|---|---|---|---|
| High-Suitable Area | Moderate-Suitable Area | Low-Suitable Area | High-Suitable Area | Moderate-Suitable Area | Low-Suitable Area | ||
| Current | 66.28 | 121.93 | 346.63 | ||||
| 2050s | RCP2.6 | 66.15 | 132.87 | 308.74 | −0.2 | 9.0 | −10.9 |
| RCP4.5 | 60.72 | 118.14 | 308.83 | 8.4 | −3.1 | −10.9 | |
| RCP8.5 | 68.65 | 150.51 | 360.55 | 3.6 | 23.4 | 4.0 | |
| 2090s | RCP2.6 | 67.32 | 118.19 | 324.75 | 1.6 | −3.1 | −6.3 |
| RCP4.5 | 66.37 | 115.99 | 282.98 | 0.1 | −4.9 | −18.4 | |
| RCP8.5 | 59.40 | 124.11 | 323.72 | −10.4 | 1.8 | −6.6 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Xu, D.; Liao, W.; Xu, Y.; Zhuo, Z. Predicting the Current and Future Distributions of Frankliniella occidentalis (Pergande) Based on the MaxEnt Species Distribution Model. Insects 2023, 14, 458. https://doi.org/10.3390/insects14050458
Wang Z, Xu D, Liao W, Xu Y, Zhuo Z. Predicting the Current and Future Distributions of Frankliniella occidentalis (Pergande) Based on the MaxEnt Species Distribution Model. Insects. 2023; 14(5):458. https://doi.org/10.3390/insects14050458
Chicago/Turabian StyleWang, Zhiling, Danping Xu, Wenkai Liao, Yan Xu, and Zhihang Zhuo. 2023. "Predicting the Current and Future Distributions of Frankliniella occidentalis (Pergande) Based on the MaxEnt Species Distribution Model" Insects 14, no. 5: 458. https://doi.org/10.3390/insects14050458