
Host-Specific Diversity of Culturable Bacteria in the Gut Systems of Fungus-Growing Termites and Their Potential Functions towards Lignocellulose Bioconversion

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Lignocellulosic Substrates
2.1.2. Growth Media and Reagents
2.2. Methods
2.2.1. Termite Gut Collection and Sample Preparations
2.2.2. Growth and Isolation of the Bacteria from Termite Gut Systems
2.2.3. DNA Extraction, PCR Amplification, and Sequencing
2.2.4. Identification and Phylogenetic Analysis of Isolated Bacteria
2.2.5. Deposition of DNA Sequences
2.2.6. Screening of the Bacteria for Cellulase and Xylanase Production
2.2.7. Growth Profile of the Potential Bacteria in BSM Media Containing Avicel or Xylan
2.2.8. Rice Straw and Corn Stover Degradation
2.2.9. Lignocellulase Production by Potential Isolates
2.2.10. Statistical Analysis
3. Results
3.1. Identification of the Termites
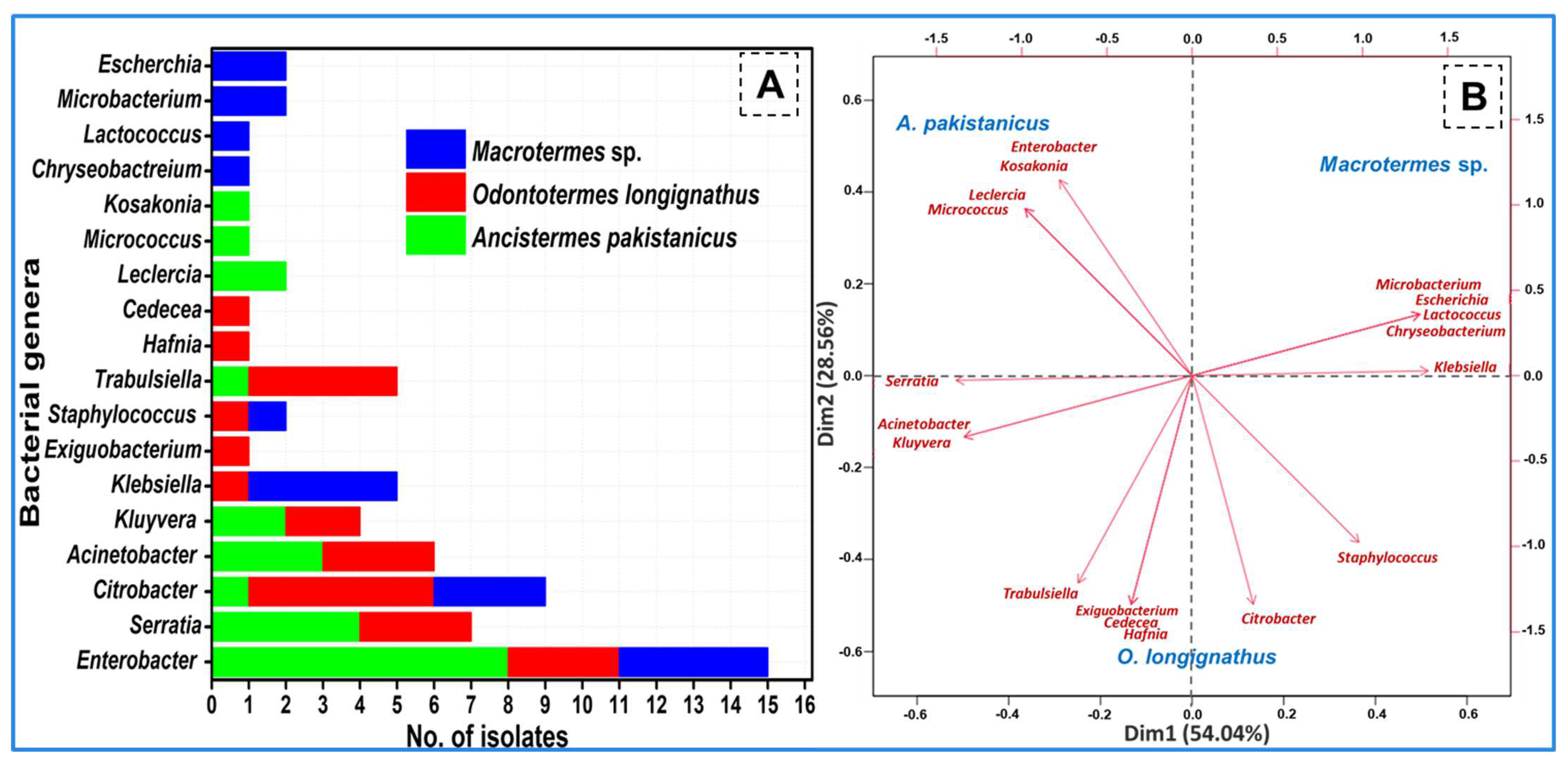
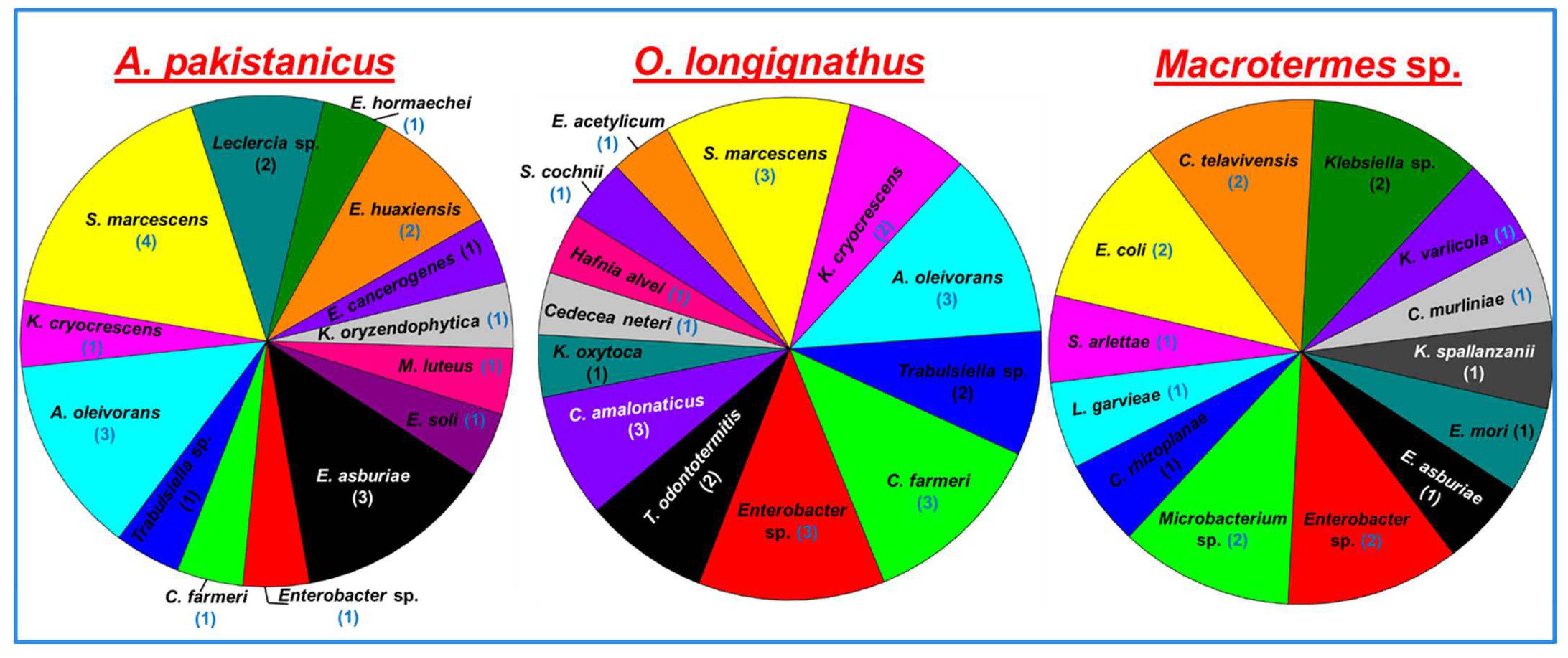
3.2. Isolation of Bacteria from the Guts of the Termites
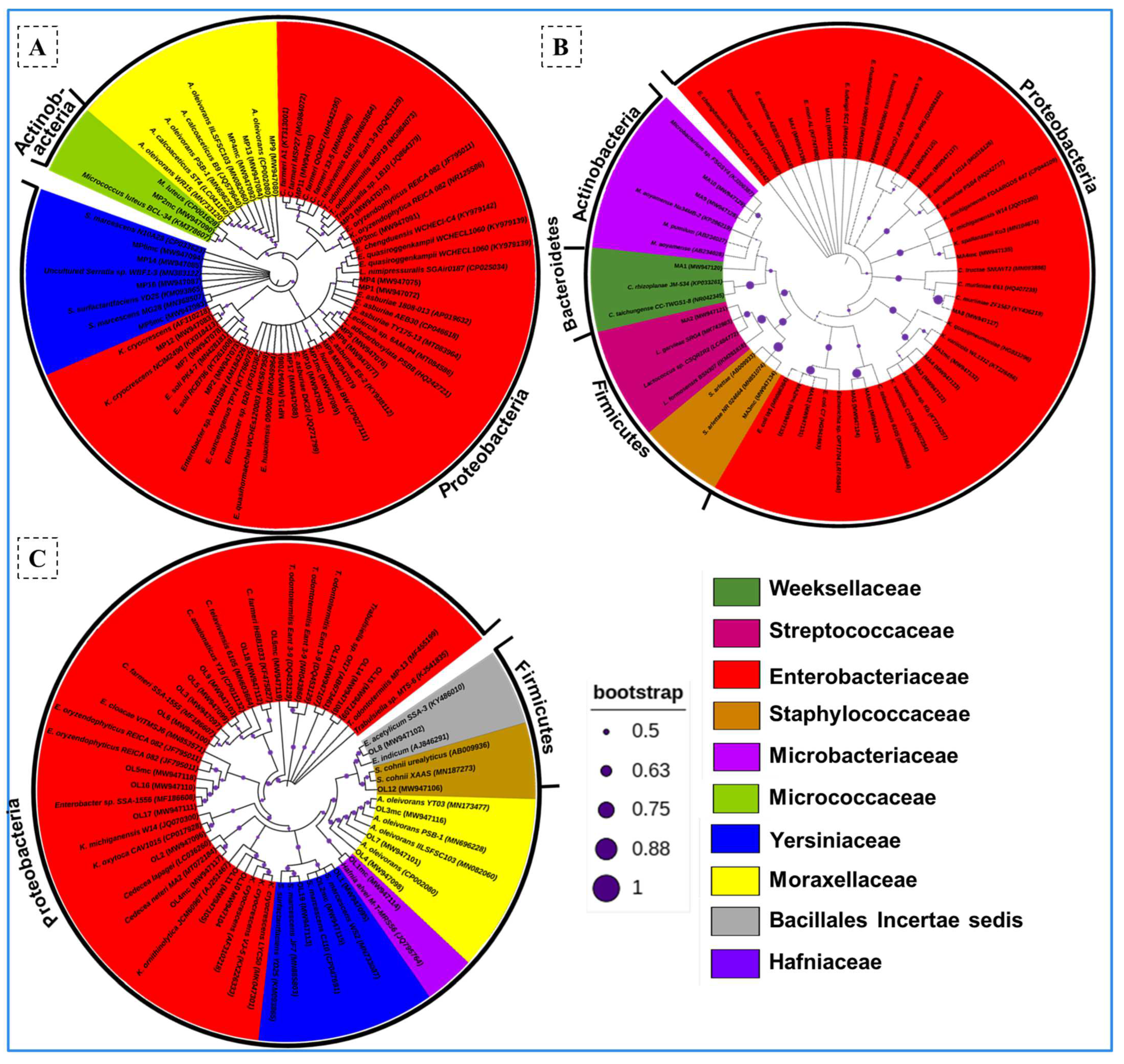
3.3. Identification and Phylogenetic Analysis of the Bacterial Isolates
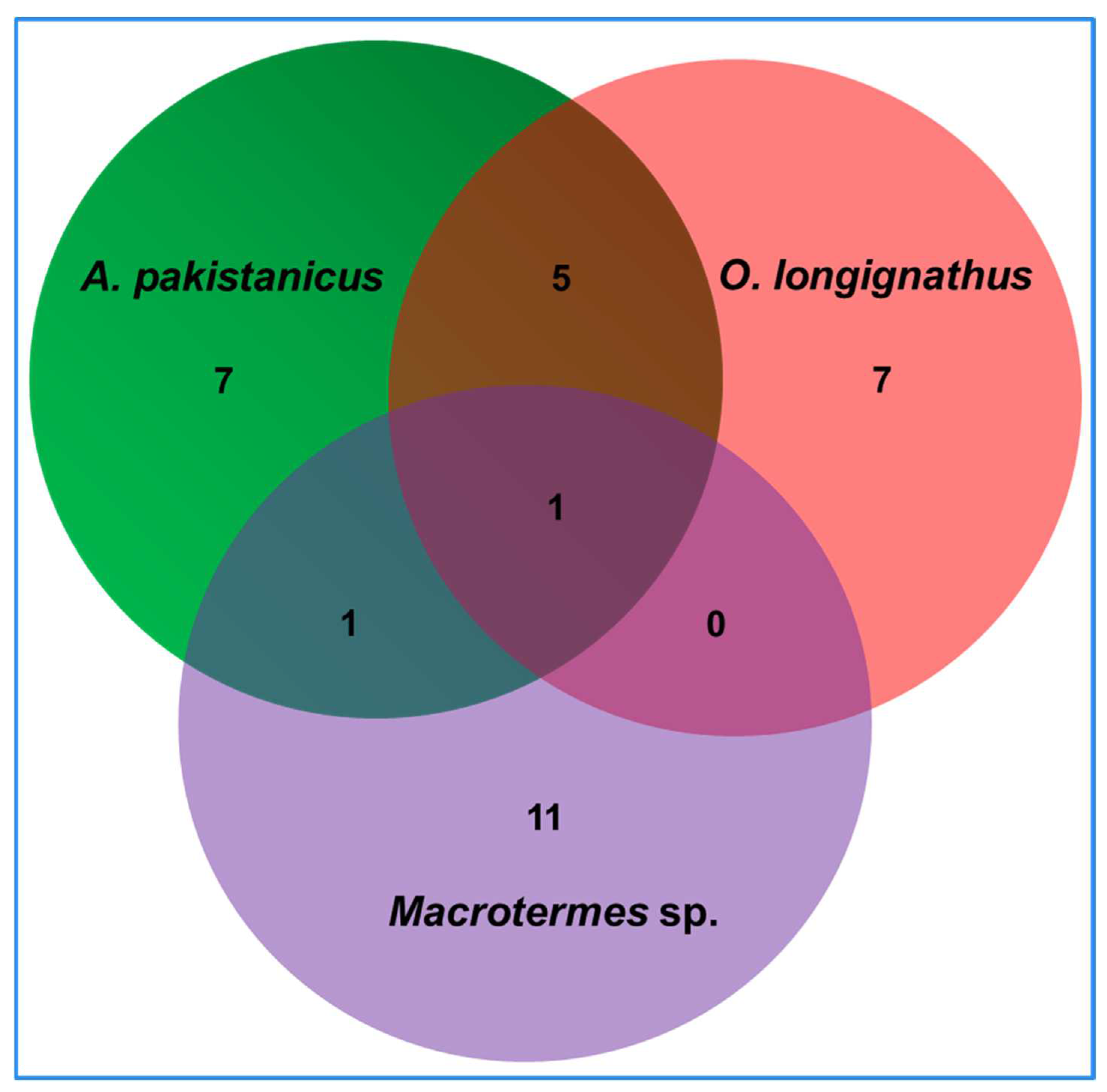
3.4. Termite-Specific Isolates
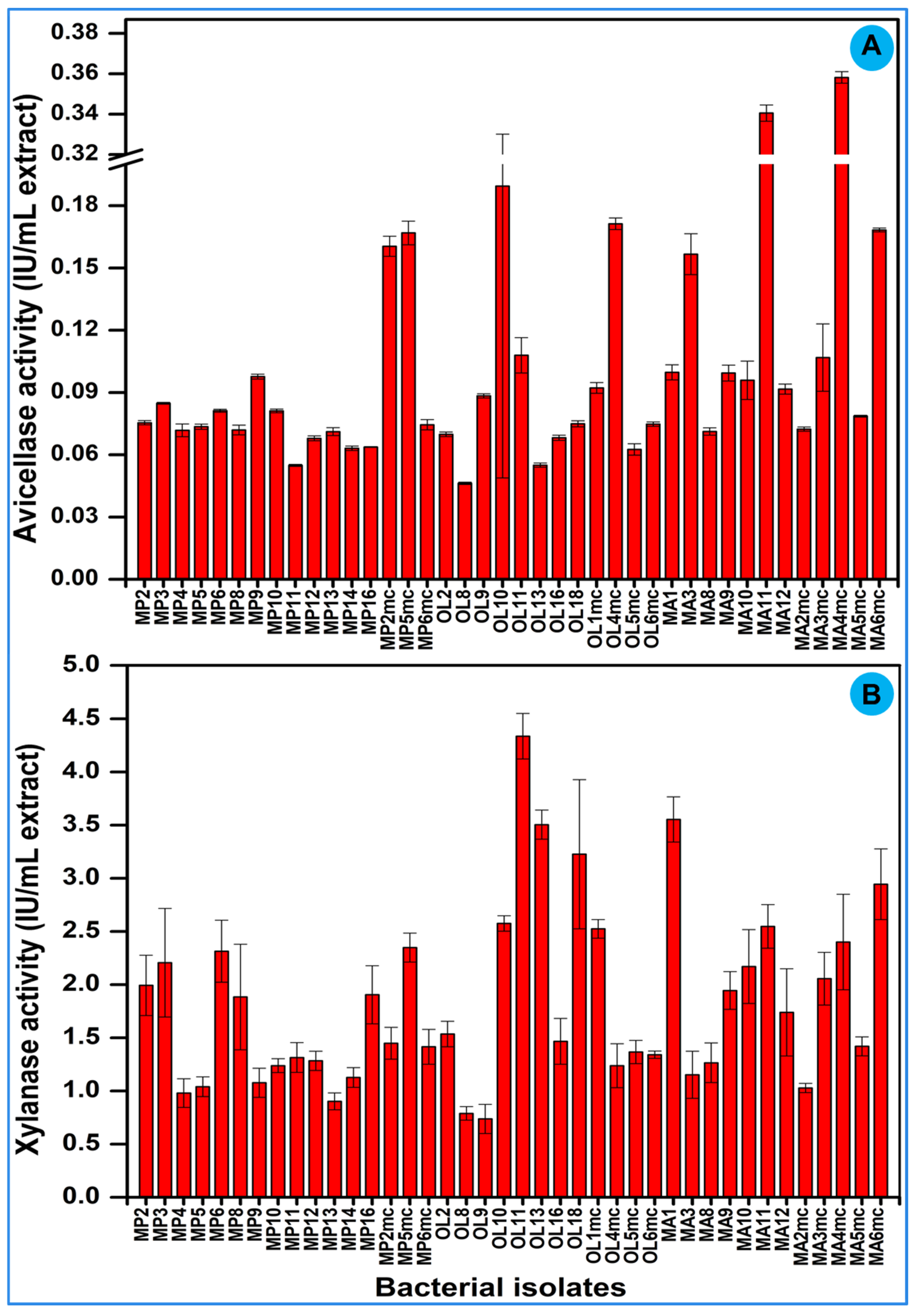
3.5. Bacterial Screening for Cellulase and Xylanase Production
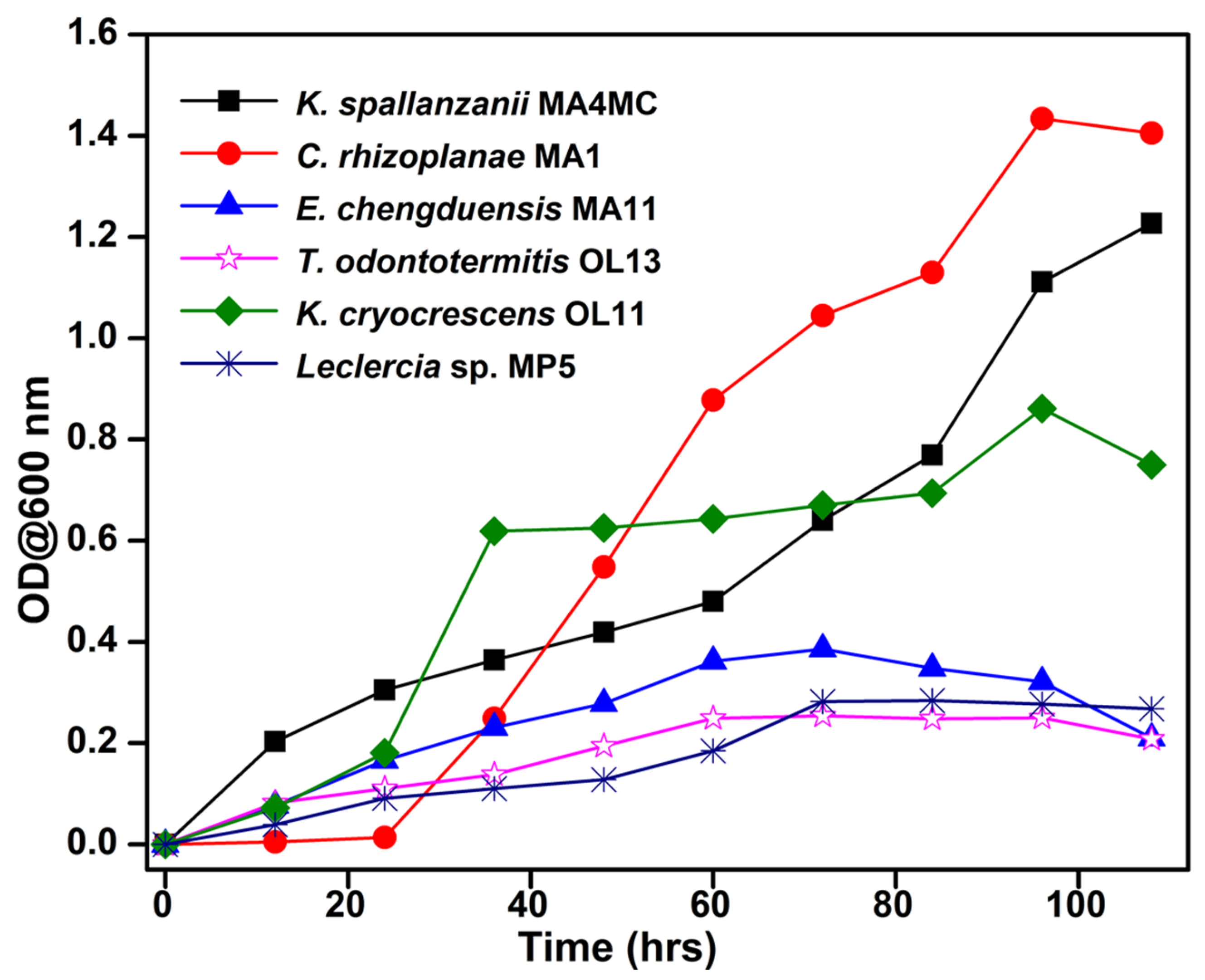
3.6. Growth of Potential Bacteria in the BSM Media Containing Avicel or Xylan
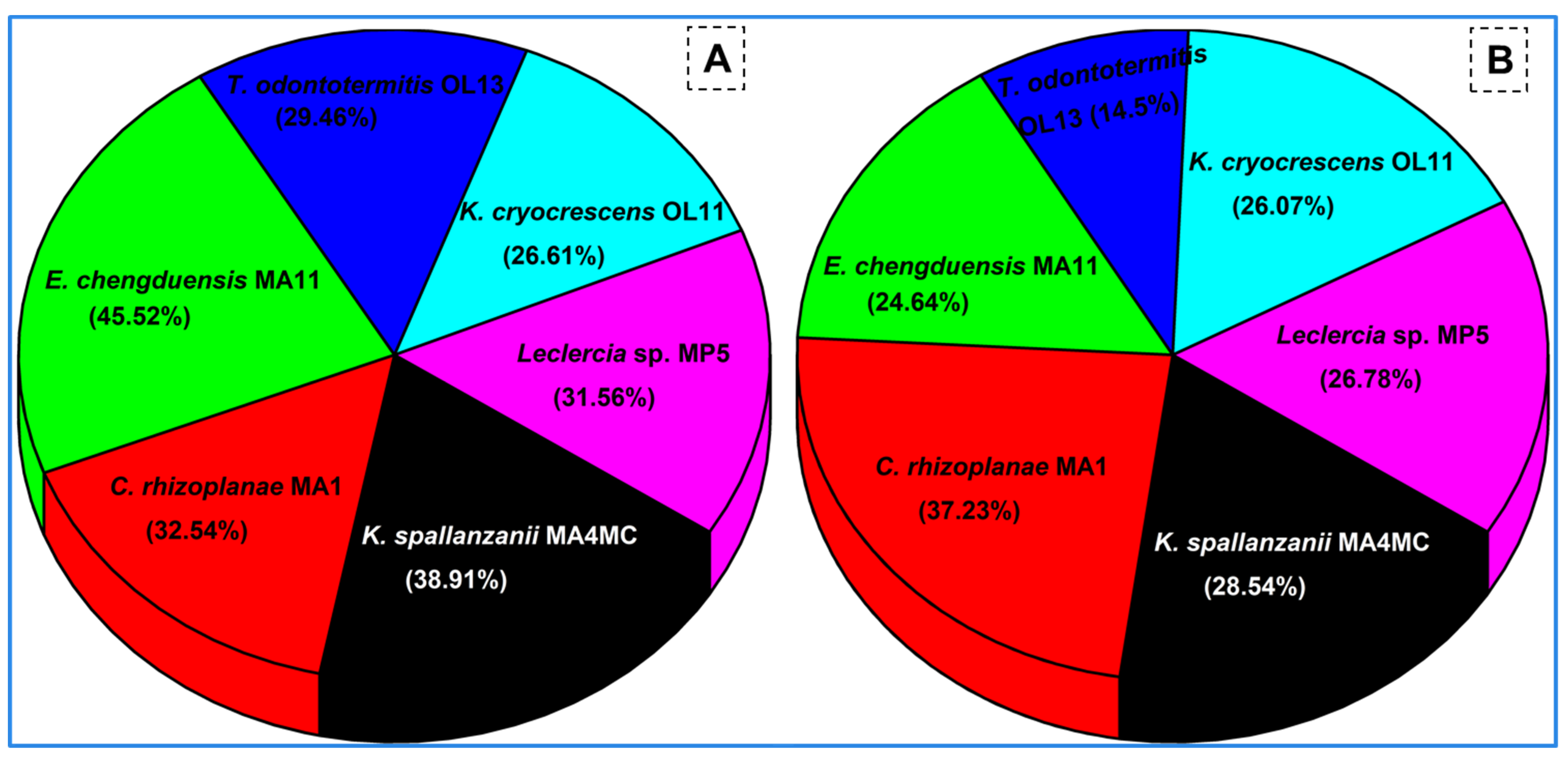
3.7. Rice Straw and Corn Stover Degradation
3.8. Lignocellulase Production by Potential Isolates
3.8.1. Endoglucanase Activity
3.8.2. Exoglucanase Activity
3.8.3. Xylanase Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Brune, A. Symbiotic digestion of lignocellulose in termite guts. Nat. Rev. Microbiol. 2014, 12, 168. [Google Scholar] [CrossRef]
- Rust, M.K.; Su, N.Y. Managing social insects of urban importance. Annu. Rev. Entomol. 2012, 57, 355–375. [Google Scholar] [CrossRef]
- Ahmad, F.; Yang, G.Y.; Liang, S.Y.; Zhou, Q.H.; Gaal, H.A.; Mo, J.C. Multipartite symbioses in fungus-growing termites (Blattodea: Termitidae, Macrotermitinae) for the degradation of lignocellulose. Insect Sci. 2021, 28, 1512–1529. [Google Scholar] [CrossRef]
- Brauman, A.; Majeed, M.Z.; Buatois, B.; Robert, A.; Pablo, A.L.; Miambi, E. Nitrous oxide (N2O) emissions by termites: Does the feeding guild matter? PLoS ONE 2015, 10, e0144340. [Google Scholar] [CrossRef]
- Pennisi, E. Africa’s soil engineers: Termites. Science 2015, 347, 596–597. [Google Scholar] [CrossRef]
- Tokuda, G.; Watanabe, H. Hidden cellulases in termites: Revision of an old hypothesis. Biol. Lett. 2007, 3, 336–339. [Google Scholar] [CrossRef]
- Bastien, G.; Arnal, G.; Bozonnet, S.; Laguerre, S.; Ferreira, F.; Fauré, R.; O’Donohue, M. Mining for hemicellulases in the fungus-growing termite Pseudacanthotermes militaris using functional metagenomics. Biotechnol. Biofuels 2013, 6, 78. [Google Scholar] [CrossRef]
- Li, H.; Yelle, D.J.; Li, C.; Yang, M.; Ke, J.; Zhang, R.; Mo, J. Lignocellulose pretreatment in a fungus-cultivating termite. Proc. Natl. Acad. Sci. USA 2017, 114, 4709–4714. [Google Scholar] [CrossRef]
- Bonachela, J.A.; Pringle, R.M.; Sheffer, E.; Coverdale, T.C.; Guyton, J.A.; Caylor, K.K.; Tarnita, C.E. Termite mounds can increase the robustness of dryland ecosystems to climatic change. Science 2015, 347, 651–655. [Google Scholar] [CrossRef]
- Evans, T.A.; Dawes, T.Z.; Ward, P.R.; Lo, N. Ants and termites increase crop yield in a dry climate. Nat. Commun. 2011, 2, 262. [Google Scholar] [CrossRef]
- Jouquet, P.; Traoré, S.; Choosai, C.; Hartmann, C.; Bignell, D. Influence of termites on ecosystem functioning. Ecosystem services provided by termites. Eur. J. Soil Biol. 2011, 47, 215–222. [Google Scholar] [CrossRef]
- Joseph, G.S.; Seymour, C.L.; Cumming, G.S.; Cumming, D.H.; Mahlangu, Z. Termite mounds as islands: Woody plant assemblages relative to termitarium size and soil properties. J. Veg. Sci. 2013, 24, 702–711. [Google Scholar] [CrossRef]
- Erens, H.; Mujinya, B.B.; Mees, F.; Baert, G.; Boeckx, P.; Malaisse, F.; Van Ranst, E. The origin and implications of variations in soil-related properties within Macrotermes falciger mounds. Geoderma 2015, 249, 40–50. [Google Scholar] [CrossRef]
- Traoré, S.; Tigabu, M.; Jouquet, P.; Ouédraogo, S.J.; Guinko, S.; Lepage, M. Long-term effects of Macrotermes termites, herbivores and annual early fire on woody undergrowth community in Sudanian woodland, Burkina Faso. Flora–Morphol. Distrib. Funct. Ecol. Plants 2015, 211, 40–50. [Google Scholar] [CrossRef]
- Brune, A. Termite guts: The world’s smallest bioreactors. Trends Biotechnol. 1998, 16, 16–21. [Google Scholar] [CrossRef]
- Scharf, M. Termites as targets and models for biotechnology. Annu. Rev. Entomol. 2015, 60, 77–102. [Google Scholar] [CrossRef]
- Auer, L.; Lazuka, A.; Sillam-Dussès, D.; Miambi, E.; O’Donohue, M.; Hernandez-Raquet, G. Uncovering the potential of termite gut microbiome for lignocellulose bioconversion in anaerobic batch bioreactors. Front. Microbiol. 2018, 8, 2623. [Google Scholar] [CrossRef]
- Poulsen, M.; Hu, H.; Li, C.; Chen, Z.; Xu, L.; Otani, S.; Zhang, G. Complementary symbiont contributions to plant decomposition in a fungus-farming termite. Proc. Natl. Acad. Sci. USA 2014, 111, 14500–14505. [Google Scholar] [CrossRef]
- Liu, N.; Yan, X.; Zhang, M.; Xie, L.; Wang, Q.; Huang, Y.; Zhou, X.; Wang, S.; Zhou, Z. Microbiome of fungus-growing termites: A new reservoir for lignocellulase genes. Appl. Environ. Microbiol. 2011, 77, 48–56. [Google Scholar] [CrossRef]
- Hongoh, Y. Toward the functional analysis of uncultivable, symbiotic microorganisms in the termite gut. Cell. Mol. Life Sci. 2011, 68, 1311–1325. [Google Scholar] [CrossRef]
- Dillon, R.J.; Dillon, V.M. The gut bacteria of insects. Non-pathogenic interactions. Annu. Rev. Entomol. 2004, 49, 71–92. [Google Scholar] [CrossRef]
- Dar, M.A.; Dhole, N.P.; Xie, R.; Pawar, K.D.; Ullah, K.; Rahi, P.; Sun, J. Valorization potential of a novel bacterial strain, Bacillus altitudinis RSP75, towards lignocellulose bioconversion: An assessment of symbiotic bacteria from the stored grain pest, Tribolium castaneum. Microorganisms 2021, 9, 1952. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Ding, S.Y.; Doran-Peterson, J. Biomass and its biorefinery: Novel approaches from nature-inspired strategies and technology. In Biological Conversion of Biomass for Fuels and Chemicals: Explorations from Natural Utilization Systems; The Royal Society of Chemistry: London, UK, 2014; pp. 1–13. [Google Scholar] [CrossRef]
- Otani, S.; Mikaelyan, A.; Nobre, T.; Hansen, L.H.; Koné, N.A.; Sørensen, S.J.; Aanen, D.K.; Boomsma, J.J.; Brune, A.; Poulsen, M. Identifying the core microbial community in the gut of fungus-growing termites. Mol. Ecol. 2014, 23, 4631–4644. [Google Scholar] [CrossRef] [PubMed]
- Nobre, T.; Kone, N.A.; Konate, S.; Linsenmair, K.E.; Aanen, D.K. Dating the fungus-growing termites’ mutualism shows a mixture between ancient codiversification and recent symbiont dispersal across divergent hosts. Mol. Ecol. 2011, 20, 2619–2627. [Google Scholar] [CrossRef] [PubMed]
- Marynowska, M.; Goux, X.; Sillam-Dussès, D.; Rouland-Lefèvre, C.; Halder, R.; Wilmes, P.; Calusinska, M. Compositional and functional characterisation of biomass-degrading microbial communities in guts of plant fibre- and soil-feeding higher termites. Microbiome 2020, 8, 96. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Kirton, L.G.; Panandam, J.M.; Siraj, S.S.; Ng, K.K.; Tan, S.G. Evidence for a higher number of species of Odontotermes (Isoptera) than currently known from Peninsular Malaysia from mitochondrial DNA phylogenies. PLoS ONE 2011, 6, e20992. [Google Scholar] [CrossRef]
- Bourguignon, T.; Lo, N.; Dietrich, C.; Šobotník, J.; Sidek, S.; Roisin, Y.; Brune, A.; Evan, T.A. Rampant Host Switch. Shaped Termit. Gut Microbiome. Curr. Biol. 2018, 28, 649–654. [Google Scholar]
- Sommer, F.; Bäckhed, F. The gut microbiota--masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef]
- Brune, A.; Dietrich, C. The gut microbiota of termites: Digesting the diversity in the light of ecology and evolution. Annu. Rev. Microbiol. 2015, 69, 145–166. [Google Scholar] [CrossRef]
- Ohkuma, M.; Brune, A. Diversity, structure, and evolution of the termite gut microbial community. In Biology of Termites: A Modern Synthesis; Bignell, D.E., Roisin, Y., Lo, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 413–438. [Google Scholar]
- Dietrich, C.; Ko¨hler, T.; Brune, A. The cockroach origin of the termite gut microbiota: Patterns in bacterial community structure reflect major evolutionary events. Appl. Environ. Microbiol. 2014, 80, 2261–2269. [Google Scholar] [CrossRef]
- Tschech, A.; Pfennig, N. Growth yield increase linked to caffeate reduction in Acetobacterium woodii. Arch. Microbiol. 1984, 137, 163–167. [Google Scholar] [CrossRef]
- Widdel, F.; Pfennig, N. Studies on dissimilatory sulfate-reducing bacteria that decompose fatty acids. I. Isolation of a new sulfate-reducer enriched with acetate from saline environments. Description of Desulfobacter postgatei gen. nov., sp. nov. Arch. Microbiol. 1981, 129, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Dar, M.A.; Shaikh, A.A.; Pawar, K.D.; Pandit, R.S. Exploring the gut of Helicoverpa armigera for cellulose-degrading bacteria and evaluation of a potential strain for lignocellulosic biomass deconstruction. Process Biochem. 2018, 73, 142–153. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Nitisinprasert, S.; Temmes, A. The characteristics of a new non-spore-forming cellulolytic mesophillic anaerobe strain CMC126 isolated from municipal sewage sludge. J. Appl. Bacteriol. 1991, 71, 154–161. [Google Scholar] [CrossRef]
- R Studio Team. RStudio: Integrated Development for R. RStudio, PBC, Boston, MA, USA. 2022. Available online: https://www.rstudio.com/ (accessed on 20 December 2022).
- Dar, M.A.; Xie, R.; Pandit, R.S.; Danso, B.; ChenChen, D.; Sun, J. Exploring the region-wise diversity and functions of symbiotic bacteria in the gut-system of wood-feeding termite, Coptotermes formosanus, towards lignocellulose degradation. Insect Sci. 2022, 29, 19. [Google Scholar] [CrossRef]
- Ali, H.R.K.; Hemeda, N.F.; Abdelaliem, Y.F. Symbiotic cellulolytic bacteria from the gut of the subterranean termite Psammotermes hypostoma Desneux and their role in cellulose digestion. AMB Express 2019, 9, 111. [Google Scholar] [CrossRef]
- Ferbiyanto, A.; Rusmana, I.; Raffiudin, R. Characterization and identification of cellulolytic bacteria from gut of worker Macrotermes gilvus. HAYATI J. Biosci. 2015, 22, 197–200. [Google Scholar] [CrossRef]
- Manjula, A.; Pushpanathan, M.; Sathyavathi, S.; Gunasekaran, P.; Rajendhran, J. Comparative analysis of microbial diversity in termite gut and termite nest using ion sequencing. Curr. Microbiol. 2016, 72, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Butera, G.; Ferraro, C.; Alonzo, G.; Colazza, S.; Quatrini, P. The gut microbiota of the wood-feeding termite Reticulitermes lucifugus (Isoptera; Rhinotermitidae). Ann. Microbiol. 2016, 66, 253–260. [Google Scholar] [CrossRef]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Lampert, N.; Mikaelyan, A.; Brune, A. Diet is not the primary driver of bacterial community structure in the gut of litter-feeding cockroaches. BMC Microbiol. 2019, 19, 238. [Google Scholar] [CrossRef]
- Hongoh, Y.; Ohkuma, M.; Kudo, T. Molecular analysis of bacterial microbiota in the gut of the termite Reticulitermes speratus (Isoptera; Rhinotermitidae). FEMS Microb. Ecol. 2003, 44, 231–242. [Google Scholar] [CrossRef]
- Shinzato, N.; Muramatsu, M.; Matsui, T.; Watanabe, Y. Molecular phylogenetic diversity of the bacterial community in the gut of the termite Coptotermes formosanus. Biosci. Biotechnol. Biochem. 2005, 69, 1145–1155. [Google Scholar] [CrossRef]
- Dar, M.A.; Pawar, K.D.; Jadhav, J.P.; Pandit, R.S. Isolation of cellulolytic bacteria from the gastro-intestinal tract of Achatina fulica (Gastropoda: Pulmonata) and their evaluation for cellulose biodegradation. Int. Biodeterior. Biodegrad. 2015, 98, 73–80. [Google Scholar] [CrossRef]
- Dar, M.A.; Syed, R.; Pawar, K.D.; Dhole, N.P.; Xie, R.; Pandit, R.S.; Sun, J. Evaluation and characterization of the cellulolytic bacterium, Bacillus pumilus SL8 isolated from the gut of oriental leafworm, Spodoptera litura: An assessment of its potential value for lignocellulose bioconversion. Environ. Technol. Innov. 2022, 27, 102459. [Google Scholar] [CrossRef]
- Barnes, M.J.; Powrie, F. The gut’s Clostridium cocktail. Science 2011, 331, 289–290. [Google Scholar] [CrossRef]
- Su, L.; Yang, L.; Huang, S.; Su, X.; Li, Y.; Wang, F.; Wang, E.; Kang, N.; Xu, J.; Song, A. Comparative gut microbiomes of four species representing the higher and the lower termites. J. Insect Sci. 2016, 16, 97. [Google Scholar] [CrossRef]
- Pourramezan, Z.; Ghezelbash, G.R.; Romani, B.; Ziaei, S.; Hedayatkhah, A. Screening and identification of newly isolated cellulose degrading bacteria from the gut of xylophagous termite Microcerotermes diversus (Silvestri). Microbiology 2012, 81, 736–742. [Google Scholar] [CrossRef]
- Adams, L.; Boopathy, R. Isolation and characterization of enteric bacteria from the hindgut of formosan termite. Bioresour. Technol. 2005, 96, 1592–1598. [Google Scholar] [CrossRef]
- Rizzi, A.; Crotti, E.; Borruso, L.; Jucker, C.; Lupi, D.; Colombo, M.; Daffonchio, D. Characterization of the bacterial community associated with larvae and adults of Anoplophora chinensis collected in Italy by culture and culture-independent methods. BioMed. Res. Int. 2013, 2013, 420287. [Google Scholar] [CrossRef]
- Ohkuma, M. Termite symbiotic systems: Efficient biorecycling of lignocellulose. Appl. Microbiol. Biotechnol. 2003, 61, 1–9. [Google Scholar] [CrossRef]
- Liu, N.; Zhang, L.; Zhou, H.; Zhang, M.; Yan, X.; Wang, Q.; Long, Y.; Xie, L.; Wang, S.; Huang, Y.; et al. Metagenomic insights into metabolic capacities of the gut microbiota in a fungus-cultivating termite (Odontotermes yunnanensis). PLoS ONE 2013, 8, e69184. [Google Scholar] [CrossRef]
- Tokuda, G.; Lo, N.; Watanabe, H.; Arakawa, G.; Matsumoto, T.; Noda, H. Major alteration of the expression site of endogenous cellulases in members of an apical termite lineage. Mol. Ecol. 2004, 13, 3219–3228. [Google Scholar] [CrossRef]
- Hu, H.; da Costa, R.R.; Pilgaard, B.; Schiøtt, M.; Lange, L.; Poulsen, M. Fungiculture in Termites Is Associated with a Mycolytic Gut Bacterial Community. mSphere 2019, 4, e00165-19. [Google Scholar] [CrossRef]
- Liu, N.; Li, H.; Chevrette, M.G.; Zhang, L.; Cao, L.; Zhou, H.; Zhou, X.; Zhou, Z.; Pope, P.B.; Currie, C.R.; et al. Functional metagenomics reveals abundant polysaccharide degrading gene clusters and cellobiose utilization pathways within gut microbiota of a wood-feeding higher termite. ISME J. 2019, 13, 104–117. [Google Scholar] [CrossRef]
- Dar, M.A.; Shaikh, A.F.; Pawar, K.D.; Xie, R.; Sun, J.; Kandasamy, S.; Pandit, R.S. Evaluation of cellulose degrading bacteria isolated from the gut system of cotton bollworm, Helicoverpa armigera and their potential values in biomass conversion. PeerJ 2021, 9, e11254. [Google Scholar] [CrossRef]
- Huang, S.; Sheng, P.; Zhang, H. Isolation and identification of cellulolytic bacteria from the gut of Holotrichia parallela larvae (Coleoptera: Scarabaeidae). Int. J. Mol. Sci. 2012, 13, 2563–2577. [Google Scholar] [CrossRef]
- Shao, Y.; Arias-Cordero, E.; Guo, H.; Bartram, S.; Boland, W. In vivo Pyro-SIP assessing active gut microbiota of the cotton leafworm, Spodoptera littoralis. PLoS ONE 2014, 9, e85948. [Google Scholar] [CrossRef]
- Ramin, M.; Alimon, A.R.; Abdullah, N. Identification of cellulolytic bacteria isolated from the termite Coptotermes curvignathus (holmgren). J. Rapid Methods Autom. Microbiol. 2009, 17, 103–116. [Google Scholar] [CrossRef]
- Konig, H. Bacillus species in the intestine of termites and other soil invertebrates. J. Appl. Microbiol. 2006, 101, 620–627. [Google Scholar] [CrossRef]
- Korsa, G.; Masi, C.; Konwarh, R.; Tafesse, M. Harnessing the potential use of cellulolytic Klebsiella oxytoca (M21WG) and Klebsiella sp. (Z6WG) isolated from the guts of termites (Isoptera). Ann. Microbiol. 2022, 72, 5. [Google Scholar] [CrossRef]
- Adams, A.S.; Jordan, M.S.; Adams, S.M.; Suen, G.; Goodwin, L.A.; Davenport, K.W.; Currie, C.R.; Raffa, K.F. Cellulose-degrading bacteria associated with the invasive woodwasp Sirex noctilio. ISME J. 2011, 5, 1323–1331. [Google Scholar] [CrossRef]
- Dantur, K.I.; Enrique, R.; Welin, B.; Castagnaro, A.P. Isolation of cellulolytic bacteria from the intestine of Diatraea saccharalis larvae and evaluation of their capacity to degrade sugarcane biomass. AMB Express 2015, 5, 15. [Google Scholar] [CrossRef]
- Banerjee, S.; Maiti, T.K.; Roy, R.N. Production, purification, and characterization of cellulase from Acinetobacter junii GAC 16.2, a novel cellulolytic gut isolate of Gryllotalpa africana, and its effects on cotton fiber and sawdust. Ann. Microbiol. 2020, 70, 28. [Google Scholar] [CrossRef]
- Arakawa, G.; Watanabe, H.; Yamasaki, H.; Maekawa, H.; Tokuda, G. Purification and molecular cloning of xylanases from the wood-feeding termite, Coptotermes formosanus Shiraki. Biosci. Biotechnol. Biochem. 2009, 73, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Tokuda, G. Lignocellulose-degrading enzymes from termites and their symbiotic microbiota. Biotechnol. Adv. 2013, 31, 838–850. [Google Scholar] [CrossRef]
- Wai, A.C.H.; Lai, G.K.K.; Griffin, S.D.J.; Leung, F.C.C. Complete genome sequence of Kluyvera sp. CRP, a cellulolytic strain isolated from red panda feces (Ailurus fulgens). Microbiol. Resour. Announc. 2022, 11, e00067-22. [Google Scholar] [CrossRef]
- Sapountzis, P.; Gruntjes, T.; Otani, S.; Estevez, J.; da Costa, R.R.; Plunkett, G.; Perna, N.T.; Poulsen, M. The enterobacterium Trabulsiella odontotermitis presents novel adaptations related to its association with fungus-growing termites. Appl. Environ. Microbiol. 2015, 81, 6577–6588. [Google Scholar] [CrossRef]
- Chou, J.H.; Chen, W.M.; Arun, A.B.; Young, C.C. Trabulsiella odontotermitis sp. nov., isolated from the gut of the termite odontotermes formosanus shiraki. Int. J. Syst. Evol. Microbiol. 2007, 57, 696–700. [Google Scholar] [CrossRef]
- Tikhe, C.V.; Sethi, A.; Delatte, J.; Husseneder, C. Isolation and assessment of gut bacteria from the Formosan subterranean termite, Coptotermes formosanus (Isoptera: Rhinotermitidae), for paratransgenesis research and application. Insect Sci. 2015, 24, 93–102. [Google Scholar] [CrossRef]
- Arevalos-Sánchez, M.M.; Maynez-Perez, A.O.; Rodríguez-Almeida, F.A.; Martínez-Quintana, J.A.; Sanchez-Flores, F.A.; Felix-Portillo, M.; Chavéz-Martínez, A.; Olvera-García, M.E.; Ruiz-Barrera, O.; Corral-Luna, A. In vitro assessment of two novel Cellulases from Trabulsiella odontotermitis for agricultural waste utilization. BMC Biotechnol. 2021, 21, 26. [Google Scholar] [CrossRef]
- Dugas, J.E.; Zurek, L.; Paster, B.J.; Keddie, B.A. Isolation and characterization of a Chryseobacterium strain from the gut of the American cockroach, Periplaneta americana. Arch. Microbiol. 2001, 175, 259–262. [Google Scholar] [CrossRef]
- Cho, M.J.; Kim, H.Y.; Shin, K.; Kim, Y.K.; Kim, Y.S.; Kim, T.J. Symbiotic adaptation of bacteria in the gut of Reticulitermes speratus: Low endo-b-1, 4-glucanase activity. Biochem. Biophys. Res. Commun. 2010, 395, 432–435. [Google Scholar] [CrossRef]
- Ventorino, V.; Aliberti, A.; Faraco, V.; Robertiello, A.; Giacobbe, S.; Ercolini, D.; Amore, A.; Fagnano, M.; Pepe, O. Exploring the microbiota dynamics related to vegetable biomasses degradation and study of lignocellulose-degrading bacteria for industrial biotechnological application. Sci. Rep. 2015, 5, 8161. [Google Scholar] [CrossRef]
- Tokuda, G.; Lo, N.; Watanabe, H. Marked variations in patterns of cellulase activity against crystalline- vs. carboxymethyl-cellulose in the digestive systems of diverse, wood-feeding termites. Physiol. Entomol. 2005, 30, 372–380. [Google Scholar] [CrossRef]
- Watanabe, H.; Nakamura, M.; Tokuda, G.; Yamaoka, I.; Scrivener, A.M.; Noda, H. Site of secretion and properties of endogenous endo-b-1,4-glucanase components from Reticulitermes speratus (Kolbe), a Japanese subterranean termite. Insect Biochem. Mol. Biol. 1997, 4, 305–313. [Google Scholar] [CrossRef]
- Inoue, T.; Murashima, K.; Azuma, J.I.; Sugimoto, A.; Slaytor, M. Cellulose and xylan utilisation in the lower termite Reticulitermes speratus. J. Insect Physiol. 1997, 43, 235–242. [Google Scholar] [CrossRef]
- Breznak, J.A. Ecology of prokaryotic microbes in the guts of wood- and litter-feeding termites. In Termites: Evolution, Sociality, Symbioses, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 209–231. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Termite | Number of Isolates (%) | Number of Isolates Obtained from Xylan (%) | Number of Isolates Obtained from Avicel (%) | Activity Showing Isolates (%) |
|---|---|---|---|---|
| Ancistrotermes pakistanicus | 23 (34.8) | 17 (74) | 6 (26) | 16 (69.5) |
| Odontotermes longignathus | 25 (37.8) | 19 (76) | 6 (24) | 12 (48) |
| Macrotermes sp. | 18 (27.2) | 12 (66.6) | 6 (33.3) | 12 (66.6) |
| Total | 66 | 48 (72.7) | 18 (27.3) | 40 (60) |
| Sr. No. | Isolate Code | Termite Species | Accession Number | NCBI Likely Species (Accession Number) | Similarity (%) | Query Cover (%) | Carbon Source Used for Isolation | Activity (IU/mL Extract) |
|---|---|---|---|---|---|---|---|---|
| 1 | MA1 | Macrotermes sp. | MW947120 | Chryseobacterium rhizoplanae (KP033261) | 97.11 | 100 | Xylan | 3.55 |
| 2 | MA3 | MW947122 | Klebsiella sp. (KT716257) | 100 | 100 | 1.15 | ||
| 3 | MA8 | MW947127 | Citrobacter murliniae (KY436219) | 100 | 100 | 1.26 | ||
| 4 | MA9 | MW947128 | Microbacterium sp. (KJ200387) | 99.95 | 96 | 1.94 | ||
| 5 | MA10 | MW947129 | 99.93 | 99 | 2.17 | |||
| 6 | MA11 | MW947130 | Enterobacter chengduensis WCHECl-C4 | 99.85 | 100 | 2.54 | ||
| 7 | MA12 | MW947131 | Escherichia coli C7 (HG941663) | 99.93 | 100 | 1.73 | ||
| 8 | MA2MC | MW947133 | Escherichia coli W5 (MN086364) | 100 | 100 | Avicel | 0.07 | |
| 9 | MA3MC | MW947134 | Staphylococcus arlettae (MN851074) | 100 | 100 | 0.10 | ||
| 10 | MA4MC | MW947135 | K. spallanzanii Ko3 (MN104674) | 99.85 | 100 | 0.35 | ||
| 11 | MA5MC | MW947136 | Citrobacter telavivensis 6105 (MN603664) | 98.93 | 100 | 0.07 | ||
| 12 | MA6MC | MW947137 | Enterobacter asburiae (MG516126) | 99.50 | 98 | 0.16 | ||
| 13 | OL2 | Odontotermes longignathus | MW947096 | Klebsiella oxytoca (CP017928) | 99.64 | 100 | 1.53 | |
| 14 | OL8 | MW947102 | Exiguobacterium acetylicum (KY486010) | 100 | 100 | 0.78 | ||
| 15 | OL9 | MW947103 | Citrobacter amalonaticus (CP011132) | 99.50 | 100 | 0.73 | ||
| 16 | OL10 | MW947104 | Kluyvera cryocrescens (AF310218) | 99.78 | 100 | 2.57 | ||
| 17 | OL11 | MW947105 | 99.86 | 100 | 4.33 | |||
| 18 | OL13 | MW947107 | Trabulsiella odontotermitis Of17 (AB673461) | 100 | 3.50 | |||
| 19 | OL16 | MW947110 | Enterobacter oryzendophyticus REICA_082 (JF795011) | 98.86 | 100 | 1.46 | ||
| 20 | OL18 | MW947112 | Citrobacter farmeri (KF475827) | 99.72 | 100 | 3.22 | ||
| 21 | OL1MC | MW947114 | Hafnia alvei (JQ795764) | 100 | 100 | Avicel | 0.09 | |
| 22 | OL4MC | MW947117 | Cedecea neteri (MT072184) | 100 | 100 | 0.17 | ||
| 23 | OL5MC | MW947118 | Enterobacter oryzendophyticus REICA_082 (JF795011) | 98.72 | 100 | 0.06 | ||
| 24 | OL6MC | MW947119 | Trabulsiella odontotermitis strain Eant 3–9 (DQ453129) | 98.83 | 99 | 0.07 | ||
| 25 | MP2 | Ancistrotermes pakistanicus | MW947073 | K. cryocrescens (KX0184413) | 99.93 | 100 | Xylan | 1.99 |
| 26 | MP3 | MW947074 | Trabulsiella sp. LB10 (JQ864379) | 99.5 | 99 | 2.20 | ||
| 27 | MP4 | MW947075 | Enterobacter asburiae (AP019632) | 99.93 | 99 | 0.98 | ||
| 28 | MP5 | MW947076 | Leclercia sp. 6AM.I94 (MT084586) | 99.93 | 100 | 1.04 | ||
| 29 | MP6 | MW947077 | Leclercia decarboxylata (HQ242721) | 99.93 | 99 | 2.31 | ||
| 30 | MP8 | MW947079 | Enterobacter asburiae (KY938112) | 99.93 | 100 | 1.88 | ||
| 31 | MP9 | MW947080 | Acinetobacter oleivorans (CP002080) | 100 | 100 | 1.07 | ||
| 32 | MP10 | MW947081 | Enterobacter cancerogenes (KT766075) | 99.93 | 99 | 1.23 | ||
| 33 | MP11 | MW947082 | Citrobacter farmeri (MH542295) | 100 | 100 | 1.31 | ||
| 34 | MP12 | MW947083 | Kluyvera cryocrescens (KX018413) | 99.72 | 100 | 1.28 | ||
| 35 | MP13 | MW947084 | Acinetobacter oleivorans (CP002080) | 100 | 100 | 0.90 | ||
| 36 | MP14 | MW947085 | Serratia marcescens (CP033623) | 99.93 | 100 | 1.12 | ||
| 37 | MP16 | MW947087 | Serratia surfactantfaciens YD25 (KM093865) | 99.78 | 100 | 1.90 | ||
| 38 | MP2MC | MW947090 | Micrococcus luteus (KM378607) | 100 | 100 | Avicel | 0.16 | |
| 39 | MP5MC | MW947093 | Serratia marcescens (MN368507) | 100 | 100 | 0.16 | ||
| 40 | MP6MC | MW947094 | Serratia marcescens (CP033623) | 100 | 100 | 0.07 |
| Enzyme Activity (IU/mL Extract) | Incubation (Days) | K. michiganensis MA4MC | C. rhizoplanae MA1 | E. chengduensis MA11 | T. odontotermitis OL13 | K. cryocrescens OL11 | Leclercia sp. MP5 |
|---|---|---|---|---|---|---|---|
| Endoglucanase (CMCase) | 1 | 0.25 ± 0.0 | 0.33 ± 0.0 | 0.15 ± 0.0 | 0.37 ± 0.0 | 0.47 ± 0.0 | 0.27 ± 0.0 |
| 2 | 0.10 ± 0.0 | 0.10 ± 0.0 | 0.09 ± 0.0 | 0.08 ± 0.0 | 0.09 ± 0.0 | 0.13 ± 0.0 | |
| 3 | 0.09 ± 0.0 | 0.10 ± 0.0 | 0.11 ± 0.0 | 0.09 ± 0.0 | 0.13 ± 0.0 | 0.12 ± 0.0 | |
| 4 | 0.06 ± 0.0 | 0.07 ± 0.0 | 0.07 ± 0.0 | 0.07 ± 0.0 | 0.06 ± 0.0 | 0.07 ± 0.0 | |
| 5 | 0.09 ± 0.0 | 0.10 ± 0.0 | 0.10 ± 0.0 | 0.09 ± 0.0 | 0.09 ± 0.0 | 0.15 ± 0.0 | |
| 6 | 0.07 ± 0.0 | 0.17 ± 0.0 | 0.08 ± 0.0 | 0.07 ± 0.0 | 0.16 ± 0.0 | 0.36 ± 0.1 | |
| 7 | 0.09 ± 0.0 | 0.24 ± 0.0 | 0.09 ± 0.0 | 0.17 ± 0.0 | 0.10 ± 0.0 | 0.35 ± 0.1 | |
| Exoglucanase (avicellase) | 1 | 0.61 ± 0.1 | 0.52 ± 0.1 | 0.38 ± 0.0 | 0.52 ± 0.2 | 0.62 ± 0.4 | 0.46 ± 0.0 |
| 2 | 0.26 ± 0.0 | 0.64 ± 0.2 | 0.37 ± 0.0 | 0.63 ± 0.2 | 0.45 ± 0.1 | 0.29 ± 0.0 | |
| 3 | 0.32 ± 0.0 | 0.16 ± 0.0 | 0.38 ± 0.1 | 0.38 ± 0.1 | 0.14 ± 0.0 | 0.42 ± 0.1 | |
| 4 | 0.21 ± 0.0 | 0.18 ± 0.0 | 0.48 ± 0.4 | 0.26 ± 0.1 | 0.30 ± 0.1 | 0.22 ± 0.0 | |
| 5 | 0.27 ± 0.0 | 0.27 ± 0.0 | 0.32 ± 0.0 | 0.30 ± 0.0 | 0.25 ± 0.0 | 0.30 ± 0.0 | |
| 6 | 0.19 ± 0.0 | 0.20 ± 0.0 | 0.32 ± 0.0 | 0.31 ± 0.1 | 0.20 ± 0.0 | 0.26 ± 0.0 | |
| 7 | 0.19 ± 0.0 | 0.28 ± 0.1 | 0.32 ± 0.0 | 0.17 ± 0.0 | 0.24 ± 0.0 | 0.20 ± 0.0 | |
| Xylanase | 1 | 1.73 ± 0.3 | 0.98 ± 0.0 | 2.11 ± 0.2 | 1.75 ± 0.1 | 0.45 ± 0.0 | 1.94 ± 0.1 |
| 2 | 2.12 ± 0.1 | 1.93 ± 0.2 | 2.43 ± 0.1 | 1.77 ± 0.5 | 1.82 ± 0.3 | 2.13 ± 0.0 | |
| 3 | 1.53 ± 0.7 | 1.78 ± 0.3 | 2.14 ± 0.2 | 1.61 ± 0.2 | 1.95 ± 0.1 | 1.80 ± 0.3 | |
| 4 | 2.99 ± 0.0 | 2.54 ± 0.1 | 2.86 ± 0.2 | 2.37 ± 0.3 | 2.91 ± 0.2 | 2.39 ± 0.4 | |
| 5 | 1.59 ± 0.2 | 1.70 ± 0.9 | 2.05 ± 0.2 | 1.28 ± 0.4 | 1.95 ± 0.3 | 2.06 ± 0.1 | |
| 6 | 2.94 ± 0.2 | 2.28 ± 0.1 | 2.76 ± 0.1 | 2.62 ± 0.3 | 2.67 ± 0.0 | 2.85 ± 0.2 | |
| 7 | 2.18 ± 0.1 | 1.79 ± 0.2 | 2.40 ± 0.2 | 2.15 ± 0.2 | 2.12 ± 0.2 | 2.23 ± 0.2 |
| Enzyme Activity (IU/mL Extract) | Incubation (Days) | K. michiganensis MA4MC | C. rhizoplanae MA1 | E. chengduensis MA11 | T. odontotermitis OL13 | K. cryocrescens OL11 | Leclercia sp. MP5 |
|---|---|---|---|---|---|---|---|
| Endoglucanase (CMCase) | 1 | 0.15 ± 0.01 | 0.19 ± 0.01 | 0.11 ± 0.03 | 0.10 ± 0.01 | 0.11 ± 0.01 | 0.14 ± 0.0 |
| 2 | 0.10 ± 0.01 | 0.10 ± 0.01 | 0.10 ± 0.0 | 0.09 ± 0.01 | 0.09 ± 0.0 | 0.09 ± 0.0 | |
| 3 | 0.10 ± 0.01 | 0.09 ± 0.0 | 0.09 ± 0.0 | 0.08 ± 0.01 | 0.09 ± 0.0 | 0.13 ± 0.0 | |
| 4 | 0.07 ± 0.0 | 0.06 ± 0.01 | 0.06 ± 0.01 | 0.06 ± 0.0 | 0.06 ± 0.0 | 0.05 ± 0.0 | |
| 5 | 0.09 ± 0.01 | 0.08 ± 0.02 | 0.09 ± 0.02 | 0.07 ± 0.01 | 0.08 ± 0.0 | 0.11 ± 0.0 | |
| 6 | 0.08 ± 0.0 | 0.09 ± 0.02 | 0.07 ± 0.0 | 0.08 ± 0.0 | 0.09 ± 0.0 | 0.08 ± 0.0 | |
| 7 | 0.13 ± 0.07 | 0.08 ± 0.01 | 0.13 ± 0.01 | 0.08 ± 0.0 | 0.07 ± 0.0 | 0.08 ± 0.0 | |
| Exoglucanase (avicellase) | 1 | 0.46 ± 0.02 | 0.32 ± 0.06 | 0.16 ± 0.01 | 0.17 ± 0.05 | 0.20 ± 0.06 | 0.29 ± 0.1 |
| 2 | 0.16 ± 0.02 | 0.33 ± 0.1 | 0.36 ± 0.05 | 0.39 ± 0.08 | 0.22 ± 0.03 | 0.23 ± 0.07 | |
| 3 | 0.32 ± 0.1 | 0.15 ± 0.01 | 0.21 ± 0.08 | 0.14 ± 0.04 | 0.25 ± 0.02 | 0.38 ± 0.2 | |
| 4 | 0.16 ± 0.02 | 0.16 ± 0.02 | 0.14 ± 0.02 | 0.31 ± 0.2 | 0.19 ± 0.05 | 0.22 ± 0.1 | |
| 5 | 0.32 ± 0.05 | 0.20 ± 0.07 | 0.15 ± 0.04 | 0.15 ± 0.01 | 0.16 ± 0.02 | 0.15 ± 0.02 | |
| 6 | 0.26 ± 0.02 | 0.20 ± 0.03 | 0.26 ± 0.04 | 0.20 ± 0.08 | 0.14 ± 0.04 | 0.13 ± 0.06 | |
| 7 | 0.16 ± 0.02 | 0.17 ± 0.04 | 0.14 ± 0.03 | 0.15 ± 0.05 | 0.16 ± 0.06 | 0.15 ± 0.06 | |
| Xylanase | 1 | 0.74 ± 0.05 | 0.61 ± 0.06 | 0.71 ± 0.05 | 0.91 ± 0.4 | 0.53 ± 0.2 | 0.75 ± 0.4 |
| 2 | 0.91 ± 0.08 | 0.97 ± 0.2 | 0.78 ± 0.4 | 1.80 ± 0.07 | 1.12 ± 0.2 | 1.05 ± 0.08 | |
| 3 | 0.48 ± 0.2 | 0.67 ± 0.2 | 0.87 ± 0.3 | 0.77 ± 0.1 | 0.67 ± 0.2 | 1.09 ± 0.01 | |
| 4 | 0.44 ± 0.4 | 1.23 ± 0.2 | 0.99 ± 0.2 | 0.95 ± 0.1 | 0.80 ± 0.1 | 1.70 ± 0.06 | |
| 5 | 0.53 ± 0.06 | 0.84 ± 0.04 | 0.71 ± 0.2 | 0.82 ± 0.07 | 0.68 ± 0.2 | 1.21 ± 0.4 | |
| 6 | 0.55 ± 0.1 | 0.33 ± 0.1 | 0.42 ± 0.2 | 0.69 ± 0.4 | 0.57 ± 0.2 | 0.54 ± 0.2 | |
| 7 | 0.26 ± 0.1 | 0.65 ± 0.1 | 0.76 ± 0.2 | 0.92 ± 0.1 | 0.72 ± 0.1 | 0.91 ± 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, R.; Dong, C.; Wang, S.; Danso, B.; Dar, M.A.; Pandit, R.S.; Pawar, K.D.; Geng, A.; Zhu, D.; Li, X.; et al. Host-Specific Diversity of Culturable Bacteria in the Gut Systems of Fungus-Growing Termites and Their Potential Functions towards Lignocellulose Bioconversion. Insects 2023, 14, 403. https://doi.org/10.3390/insects14040403
Xie R, Dong C, Wang S, Danso B, Dar MA, Pandit RS, Pawar KD, Geng A, Zhu D, Li X, et al. Host-Specific Diversity of Culturable Bacteria in the Gut Systems of Fungus-Growing Termites and Their Potential Functions towards Lignocellulose Bioconversion. Insects. 2023; 14(4):403. https://doi.org/10.3390/insects14040403
Chicago/Turabian StyleXie, Rongrong, Chenchen Dong, Shengjie Wang, Blessing Danso, Mudasir A. Dar, Radhakrishna S. Pandit, Kiran D. Pawar, Alei Geng, Daochen Zhu, Xia Li, and et al. 2023. "Host-Specific Diversity of Culturable Bacteria in the Gut Systems of Fungus-Growing Termites and Their Potential Functions towards Lignocellulose Bioconversion" Insects 14, no. 4: 403. https://doi.org/10.3390/insects14040403