
The Dynamic Distribution of Wolbachia and Rickettsia in AsiaII1 Bemisia tabaci
by
 and
and
Ning Lv
1,
Jing Peng
2,
Zi-Qi He
2,
Qin Wen
2,
Zheng-Qin Su
2,
Shaukat Ali
2,
Chang-Zhong Liu
1,* and
Bao-Li Qiu
2,3,4,* 1
Biocontrol Engineering Laboratory of Crop Diseases and Pests of Gansu Province, College of Plant Protection, Gansu Agricultural University, Lanzhou 730070, China
2
Key Laboratory of Bio-Pesticide Innovation and Application of Guangdong Province, South China Agricultural University, Guangzhou 510642, China
3
Engineering Research Center of Biocontrol, Ministry of Education, Guangzhou 510642, China
4
Chongqing Key Laboratory of Vector Insects, College of Life Sciences, Chongqing Normal University, Chongqing 401331, China
*
Authors to whom correspondence should be addressed.
Insects 2023, 14(4), 401; https://doi.org/10.3390/insects14040401
Submission received: 22 March 2023
/
Revised: 16 April 2023
/
Accepted: 17 April 2023
/
Published: 21 April 2023
Abstract
:Simple Summary
Bacterial endosymbionts play important roles in the life history of herbivorous insects, including supplying nutrients allowing exploitation of unbalanced diets, increasing the survivorship and fecundity of hosts, protecting hosts against entomopathogenic fungi and parasitoid wasps, ameliorating the detrimental effects of heat, and broadening the range of suitable food plants. The important function of bacterial endosymbionts is that they can manipulate the reproduction of their hosts. However, all functions of endosymbionts must rely on vertical transmission to spread within the population, such as Wolbachia, Rickettsia, and Cardinium. So, we studied the spatial and temporal distribution of Wolbachia and Rickettsia in Asia II 1 Bemisia tabaci. The results showed that the titers of Wolbachia and Rickettsia in the 3–120 h old eggs changed in a “w” pattern, and the location of Wolbachia and Rickettsia in the egg changed from egg stalk to egg base, and then from the egg base to egg posterior, and finally back to the middle of egg in a Rickettsia and Wolbachia coinfected whitefly host. Our study helps to explain the vertical transmission mechanism of bacterial endosymbionts and the distribution of bacterial endosymbionts in different tissues in the host.
Abstract
Wolbachia and Rickettsia are bacterial endosymbionts that can induce a number of reproductive abnormalities in their arthropod hosts. We screened and established the co-infection of Wolbachia and Rickettsia in Bemisia tabaci and compared the spatial and temporal distribution of Wolbachia and Rickettsia in eggs (3–120 h after spawning), nymphs, and adults of B. tabaci by qPCR quantification and fluorescent in situ hybridization (FISH). The results show that the titer of Wolbachia and Rickettsia in the 3–120 h old eggs showed a “w” patterned fluctuation, while the titers of Wolbachia and Rickettsia had a “descending–ascending descending–ascending” change process. The titers of Rickettsia and Wolbachia nymphal and the adult life stages of Asia II1 B. tabaci generally increased with the development of whiteflies. However, the location of Wolbachia and Rickettsia in the egg changed from egg stalk to egg base, and then from egg base to egg posterior, and finally back to the middle of the egg. These results will provide basic information on the quantity and localization of Wolbachia and Rickettsia within different life stages of B. tabaci. These findings help to understand the dynamics of the vertical transmission of symbiotic bacteria.
1. Introduction
The symbiotic relationship between insects and endosymbionts is widely prevalent [1]. Obligate endosymbionts are critical determinants of host physiology by complementing the incomplete or absent eukaryotic metabolic pathways required for the synthesis of essential nutrients with limited availability in the host diets [2]. Plant-sap-sucking insects are always dependent on their primary prokaryotic endosymbionts for survival, such as Buchnera in aphids and Carsonella in psyllids [3,4]. Additionally, these host insects may have one or more facultative endosymbionts, which are not necessary for the host’s survival or reproduction, but they can have important effects on the biology and ecology of the hosts, including physiology, evolution, reproduction, immune homeostasis, and defense [5,6,7]. Intracellular bacteria of insect hosts are vertically transmitted in different ways. Bacteria are released from the bacteriocytes and transferred to the ovary in most insects, but bacteriocytes migrate to each egg, and bacteria are released from bacteriocytes in whiteflies [8]. The vertical transmission mechanism of symbiotic bacteria can reshape bacteriocytes during the developmental transition from nymph to adulthood, including the loss of cell–cell adhesion, high division rates to constant cell size, and the onset of cell mobility, so that the bacteriocytes can crawl to the ovary [9].
Bemisia tabaci (Gennadius, s1889) (Hemiptera: Aleyrodidae) is a complex of cryptic species, among which at least 40 cryptic species are morphologically indistinguishable but biologically different, and has become one of the most economically important crop pests worldwide [10,11]. The B. tabaci species complex shares a long-term and intimate association with only one obligate bacterial symbiont (Portiera) and facultative bacterial endosymbionts (belonging to seven genera Arsenophonus, Cardinium, Hamiltonella, Hemipteriphilus, Wolbachia, Fritschea, and Rickettsia) [12]. Wolbachia and Rickettsia are the most studied endosymbionts. Both of them can affect some aspects of host fitness, such as weight, fecundity, longevity, and defense [13,14,15]. Furthermore, Wolbachia and Rickettsia are the master manipulators of arthropod reproduction that can induce a number of reproductive abnormalities, such as male killing, feminization, parthenogenesis, and cytoplasmic incompatibility (CI), thus increasing the spread of Wolbachia in host populations [16,17]. CI has attracted much attention due to its application in the biological control of insect pests [16,17,18,19,20]. The utilization of Wolbachia or Rickettsia as a pest control agent requires generating novel Wolbachia or Rickettsia–host interactions by transinfection or microinjection of Wolbachia or Rickettsia strains into different insect pests [21]. Although there are a large number of studies on the effects of Wolbachia or Rickettsia on host fitness and behavior and their potential application as biological control agents [22], there is a lack of detailed information on the behavior of bacteria within different tissues during the host’s life cycle.
In this study, the spread of Wolbachia and Rickettsia spread within different life stages of B. tabaci was detected under laboratory conditions. We compared the spatial and temporal distribution of Wolbachia and Rickettsia in eggs (3–120 h after egg laying), nymphs, and adults of B. tabaci using qPCR quantification and fluorescent in situ hybridization (FISH). These results will further enhance our understanding of the vertical transmission of symbiotic bacteria in insects. This study will also provide basic dynamic change pattern on the quantity and localization of Wolbachia and Rickettsia within different life stages of B. tabaci and will serve as a reference for the selection of the optimal time for the horizontal transfer of symbiotic bacteria within the eggs of different whiteflies by microinjection techniques, to study the interactions between the same symbiotic bacteria and different whitefly cryptic species or transfection of symbiotic bacteria inducing cytoplasmic incompatibility for pest control by microinjection technique.
2. Materials and Methods
2.1. Host Plant and Whitefly Cultures
The cotton plant Gossypium hirsutum Linnaeus 1763. (var. Lumianyan no. 32) was used to rear B. tabaci Asia II 1. Cultivate cotton seedlings according to Li et al. [23]. Healthy plants at the six to eight expanded-leaf stage were used in experiments. B. tabaci Asia II 1 individuals were collected from the soybean plants, Glycine max (Linn.) Merr in Guangzhou in 2017 and reared on the cotton plants at 26.0 ± 0.5 °C and 70–80% relative humidity under a 14:10 h (L:D) photoperiod in the laboratory at South China Agricultural University. The species and infection rate of symbiotic bacteria in the original population of B. tabaci were determined by PCR detection, and the experimental population line was screened and established based on the original laboratory line using the “single-pair” and PCR detection method (newly emerged single pair of whiteflies (one female and one male) were released into a leaf cage, which was attached onto a clean cotton leaf to allow egg laying to establish the population), and a laboratory line that was coinfected with Rickettsia and Wolbachia (IWR).
2.2. PCR Detection of Rickettsia and Wolbachia in Asia II1 Whitefly
On the basis of clarifying the type and infection rate of secondary endosymbionts, newly emerged adults were randomly selected in the original population was measured by PCR with special primers of 16S rRNA, 23S rRNA, and wsp genes of endosymbionts (Supplementary Table S1). Results revealed that the original laboratory line of Asia II1 whitefly was infected with Rickettsia and Wolbachia, but the infection percentages of Rickettsia and Wolbachia in whitefly adults were 100% and 83.3% (Supplementary File 1). So, the experimental line with co-infection of Rickettsia and Wolbachia was established using the “single-pair screen” and PCR detection method described by Liu et al. [24]. Adult individuals were randomly selected for the PCR detection of secondary endosymbionts Rickettsia and Wolbachia. Specific primers and cycling conditions are shown in Table S1. Total genomic DNA of individual whitefly was extracted according to Ahmed et al. [25]. PCRs were performed in a 25 μL volume containing 1 μL of the template DNA lysate, 2.5 mM MgCl2, 200 mM for each dNTP, 1 μM of each primer, and 1 unit of DNA Taq polymerase (Invitrogen, Guangzhou, China). A 3 μL volume of the PCR product was visualized on 1% agarose gel containing GoldView colorant (Invitrogen, Guangzhou, China) and then photographed. When bands with the expected size were visible on the gels, the remaining 17 μL of PCR product was sent for sequencing in Sangon Biotech (Shanghai) Co., Ltd. Portiera aleyrodidarum DNA was used as a positive control and ddH2O was used as a negative control to eliminate potential confounding variables and judge the quality of extracted DNA.
2.3. Infection Monitoring and Maternal Transmission Efficiency of Rickettsia and Wolbachia in Laboratory Lines
The co-infection status of Rickettsia and Wolbachia in the IWR line, which included both females and males, was monitored prior to and during the experiments using polymerase chain reaction (PCR) using Rickettsia-specific 16S rRNA and Wolbachia-specific wsp primers as described previously [26,27]. A single experiment to detect the endosymbiont of whitefly was considered as one repeat, and the experiment was repeated 60 times. The infection status of Rickettsia and Wolbachia was monitored for three generations. The maternal transmission efficiency of Rickettsia and Wolbachia was studied according to the methods of Nguyen [28]. Ten mothers were arbitrarily selected and tested to determine if they undergo arrhenotokous or amphigenetic reproduction. Then, the maternal transmission efficiency of Rickettsia and Wolbachia was recorded in the arrhenotokous parthenogenetic progeny of IWR virgin females and the amphigenetic progeny of IWR virgin females and IWR virgin males. The infection status of Rickettsia and Wolbachia was tested with PCR using Rickettsia-specific 16S rRNA and Wolbachia-specific wsp primers (Supplementary Table S1) to measure the maternal transmission efficiency of Rickettsia and Wolbachia, and 10 male or female progenies of each mother were examined for a total of 100 progeny individuals per setup.
2.4. Quantitative Detection of Rickettsia and Wolbachia
To collect eggs, nymphs, and newly emerged males and females of Asia II1 for quantitative detection of endosymbionts, 500 females and males of IWR lines were isolated with leaf cages on cotton leaves. The tested insects were allowed to oviposit for 3 h, followed by their removal from the cages. Eggs were collected at 3, 6, 9, 12, 18, 24, 36, 48, 72, 96, and 120 h post adult removal. Newly emerged nymphs of each instar (from first to fourth instar) samples were collected. Newly emerged adults were collected and separated with dactylethrae, and their sex was examined under a microscope. DNA samples were extracted using a TIANamp Genomic DNA kit (Tiangen, Beijing, China). Quantitative real-time PCR (qPCR) was used to detect the titer of Rickettsia and Wolbachia in the line of B. tabaci Asia II1 following the protocol of Ghanim and Kontsedalov [29]. The qPCR was performed in a 10 μL reaction system, including 5 μL SYBR Premix (ThunderbirdTM, Osaka, Japan), 10 μmol of each primer, 2 μL extracted DNA, and 1 μL ddH2O (Total 10 μL). The procedure was 5 min activation at 95 °C, then 45 cycles of 20 s at 95 °C, 30 s at 60 °C, and 45 s at 72 °C in a quantitative real-time PCR machine (CFX-96, Bio-Rad Co. Ltd.). Melting ramp included 30 s at 50 °C to 98 °C, rising 1 °C at each step, and waiting 5 s after each step. The primers of gltA and coxA genes were used as described previously [30,31]. A β-actin whitefly gene was selected as an internal control for data normalization and quantification [29]. Thirty sets of eggs, thirty nymphs of each instar, and thirty pairs of newly emerged males and females of the IWR line quantified the titer of Rickettsia and Wolbachia as one repeat, and the experiment was repeated three times. Three technical replicates were performed for each biological replicate.
2.5. Fluorescence In Situ Hybridization Microscopy of Rickettsia and Wolbachia in Asia II1 Whiteflies
Eggs, nymphs, and adults of Asia II1 were collected for fluorescence in situ hybridization visualization of endosymbionts. Localizations of Wolbachia and Rickettsia in the eggs, nymphs, and adults of whiteflies was studied by FISH using the previously described protocol [31]. Whitefly samples were fixed overnight in Carnoy’s solution (chloroform–ethanol–acetic acid 6:3:1) at 4 °C. The samples were washed three times (5 min per wash) in 50% ethanol, followed by decolorization with 6% H2O2 in ethanol for 24 h in the dark and incubation in a hybridization buffer (20 mmol/L Tris-HCl pH 8.0, 0.9 mol/L NaCl, 0.01% sodium dodecyl sulfate, 30% formamide) containing 10 pmol/mL of the 6-FAM-labeled Wolbachia 16S rRNA probe and the Cyanine3-labeled Rickettsia 16S rRNA probe as described previously [32]. After overnight incubation, the samples were thoroughly washed in the washing buffer (0.3 mol/L NaCl, 0.03 mol/L sodium citrate, 0.01% sodium dodecyl sulfate) and then observed under a Nikon eclipse Ti-U FluoView inverted microscope (Nikon Instruments Inc., Tokyo, Japan).
2.6. Data Analysis
The Bio-Rad real-time PCR system (USA) and accompanying software (Bio-Rad CFX Manager) were used for qPCR data normalization, and the relative quantities of endosymbionts were calculated using the method of 2−ΔΔCt [33]. Statistical analysis was performed using IBM SPSS Statistics version 19.0 (SPSS 19.0) (IBM, Armonk, NY, USA). Tukey’s HSD was used for analysis of variance (ANOVA) to indicate significant differences between different nymphs or adults of whitefly. Figures were generated using SigmaPlot 10.0. Error bars in all graphs represent standard error.
3. Results
3.1. Rickettsia and Wolbachia Detection in Different Asia II1 Whiteflies
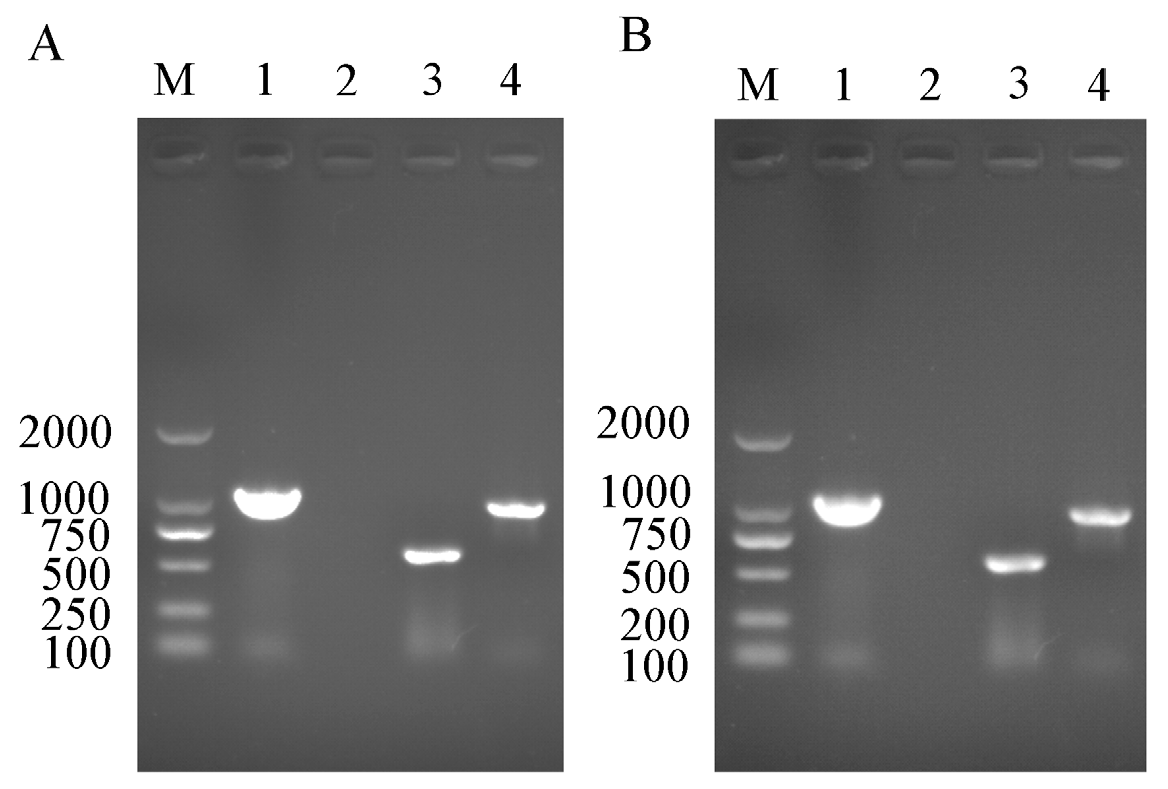
In order to confirm the infection status of Rickettsia and Wolbachia in the IWR line of B. tabaci Asia II1, PCR detection was confirmed by the presence of Rickettsia and Wolbachia in adult males and females of B. tabaci Asia II1 (Figure 1).
3.2. Maternal Transmission Efficiency of Rickettsia and Wolbachia of AsiaII1 IWR Line
The maternal transmission efficiency of Rickettsia and Wolbachia in the IWR line of Asia II1 whiteflies was detected in the arrhenotokous male offspring of virgin females, amphigenetic female offspring of IWR females crossed males, indicating that the revealed complete (100%) maternal transmission of Rickettsia and Wolbachia was observed in the male and female progeny of IWR line (Table 1).
3.3. Quantitative Detection of Rickettsia and Wolbachia in Eggs of Asia II1 Whitefly
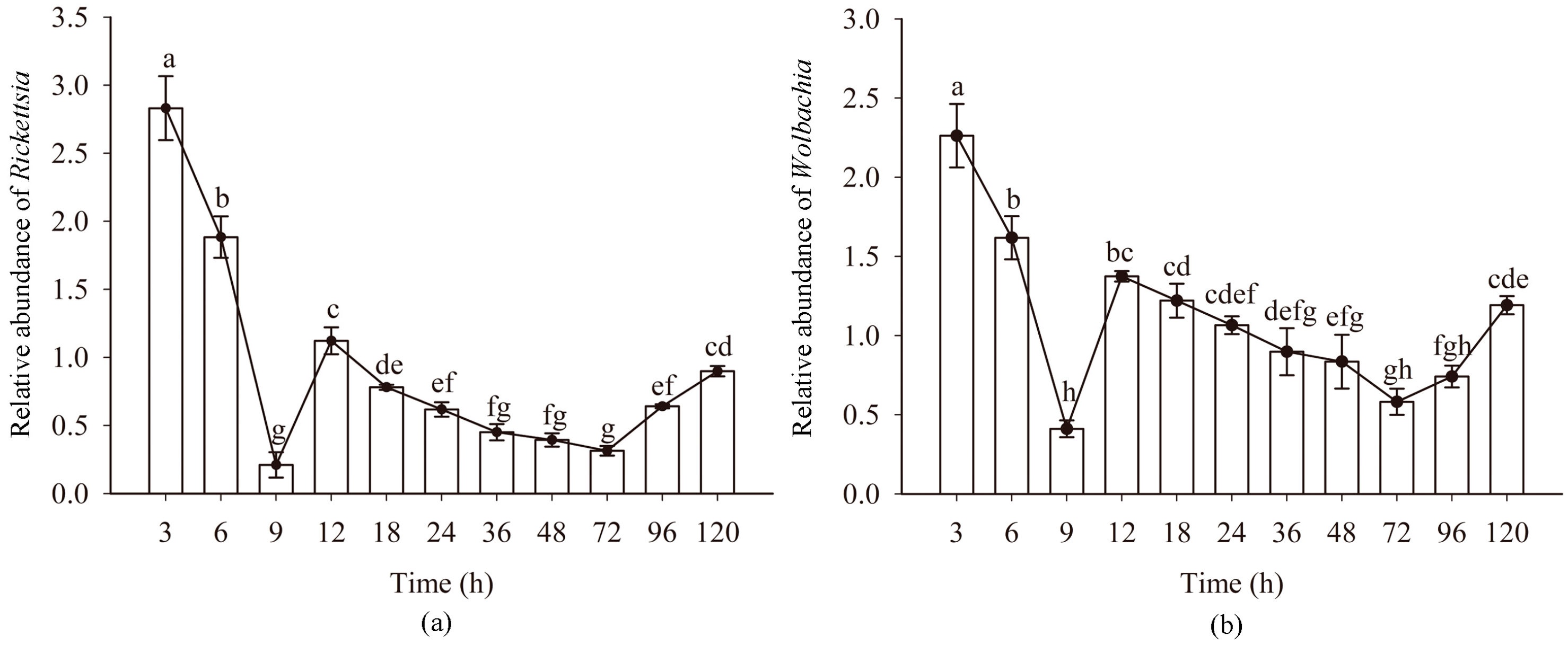
The titers Rickettsia and Wolbachia in eggs of Asia II1 B. tabaci showed a consistent pattern (“W” shape) at different time intervals. The titers of endosymbionts Rickettsia and Wolbachia showed a downward trend at 3–9 h and 12–72 h, especially at 3–9 h. The titers of endosymbionts Rickettsia and Wolbachia showed an upward trend at 9–12 h and 72–120 h. Compared with other time intervals, the increase at 9–12 h was significantly higher (Figure 2). During the dynamic change of the titer of symbiotic bacteria, the titer of Rickettsia and Wolbachia was the highest in 3 h eggs and the lowest in 9 h eggs. The variation of Protiera titer showed the same as that of Rickettsia and Wolbachia (Figure S1).
3.4. FISH Visualization of Rickettsia and Wolbachia in Eggs at Different Times
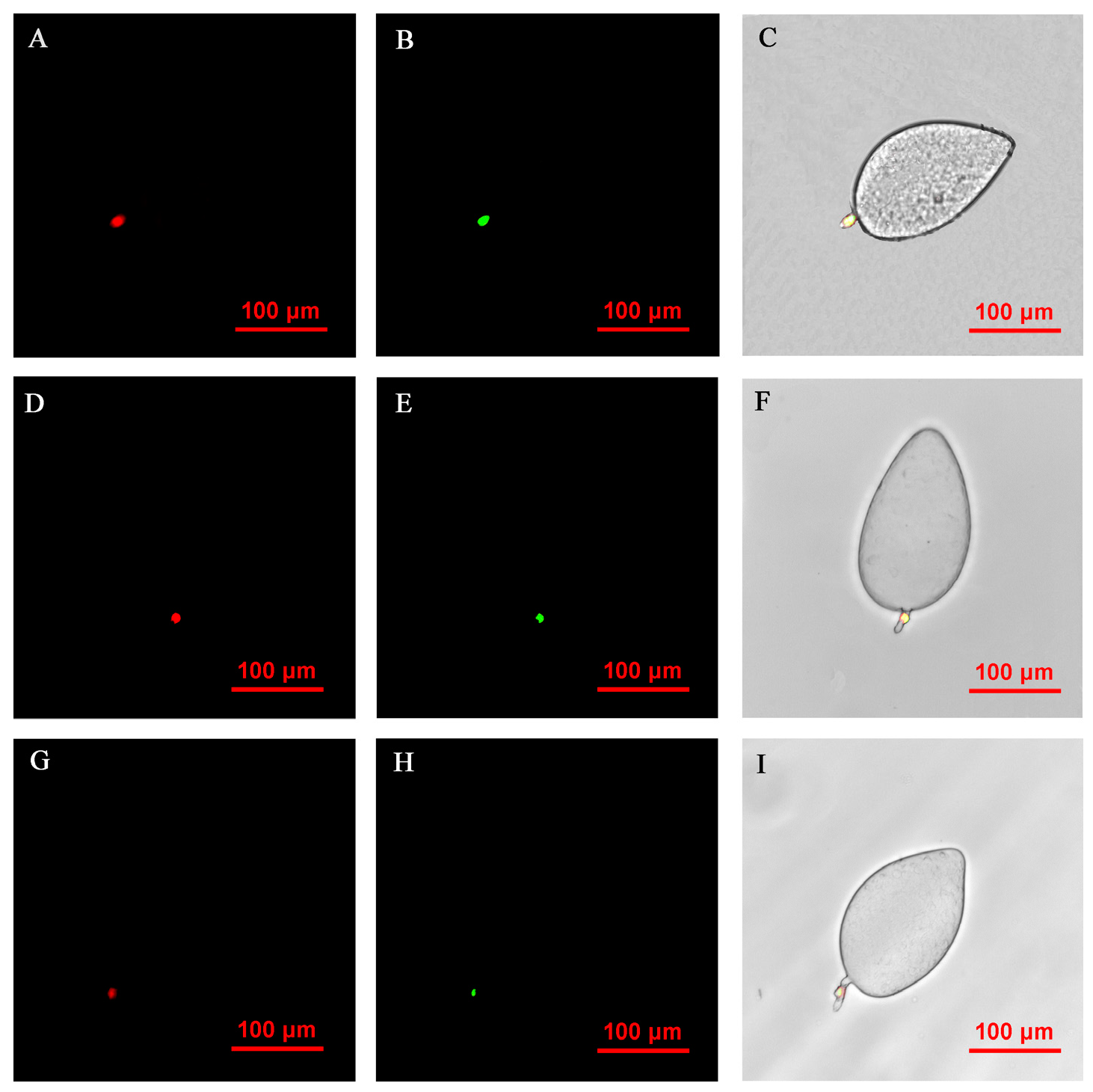
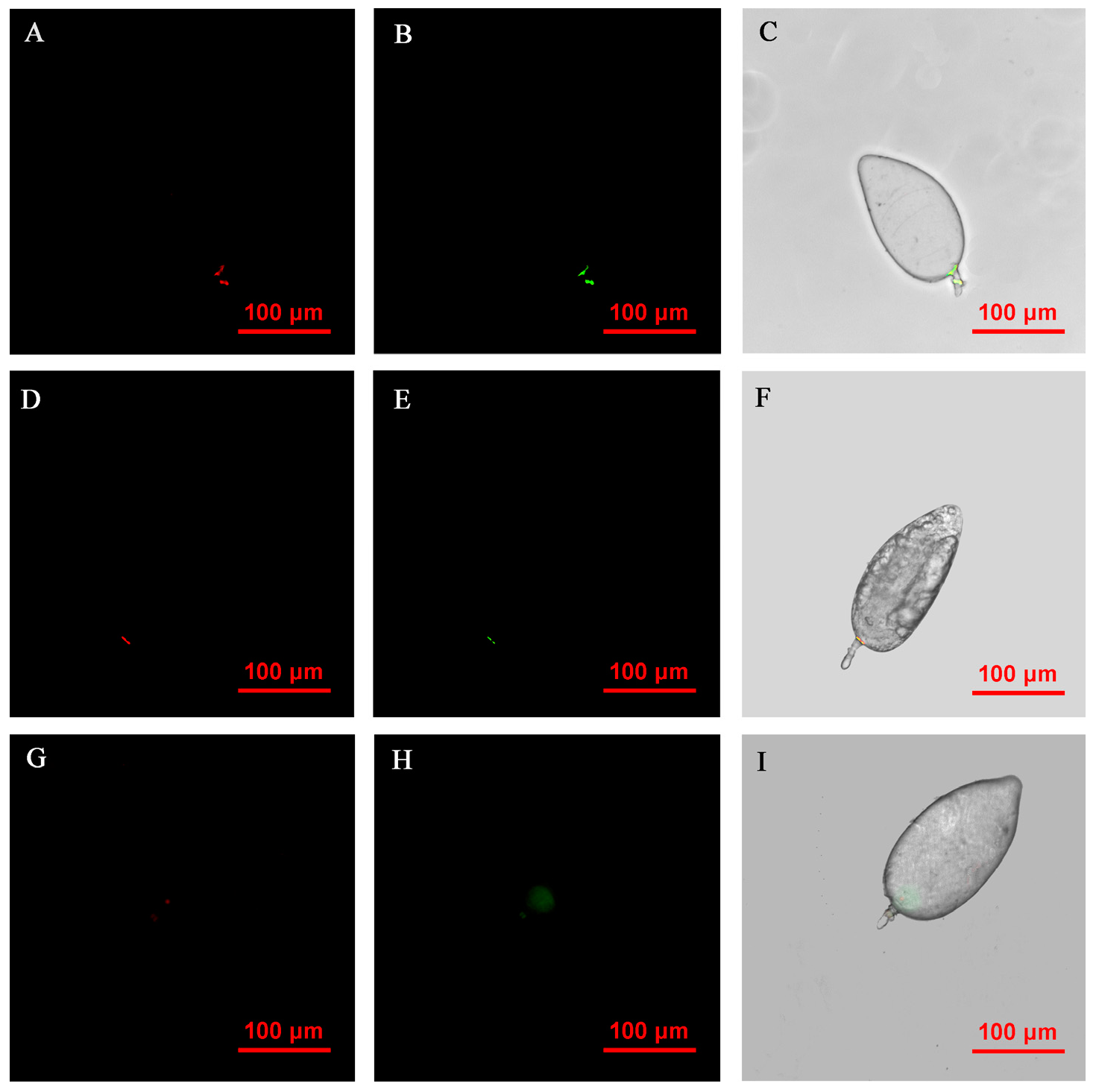
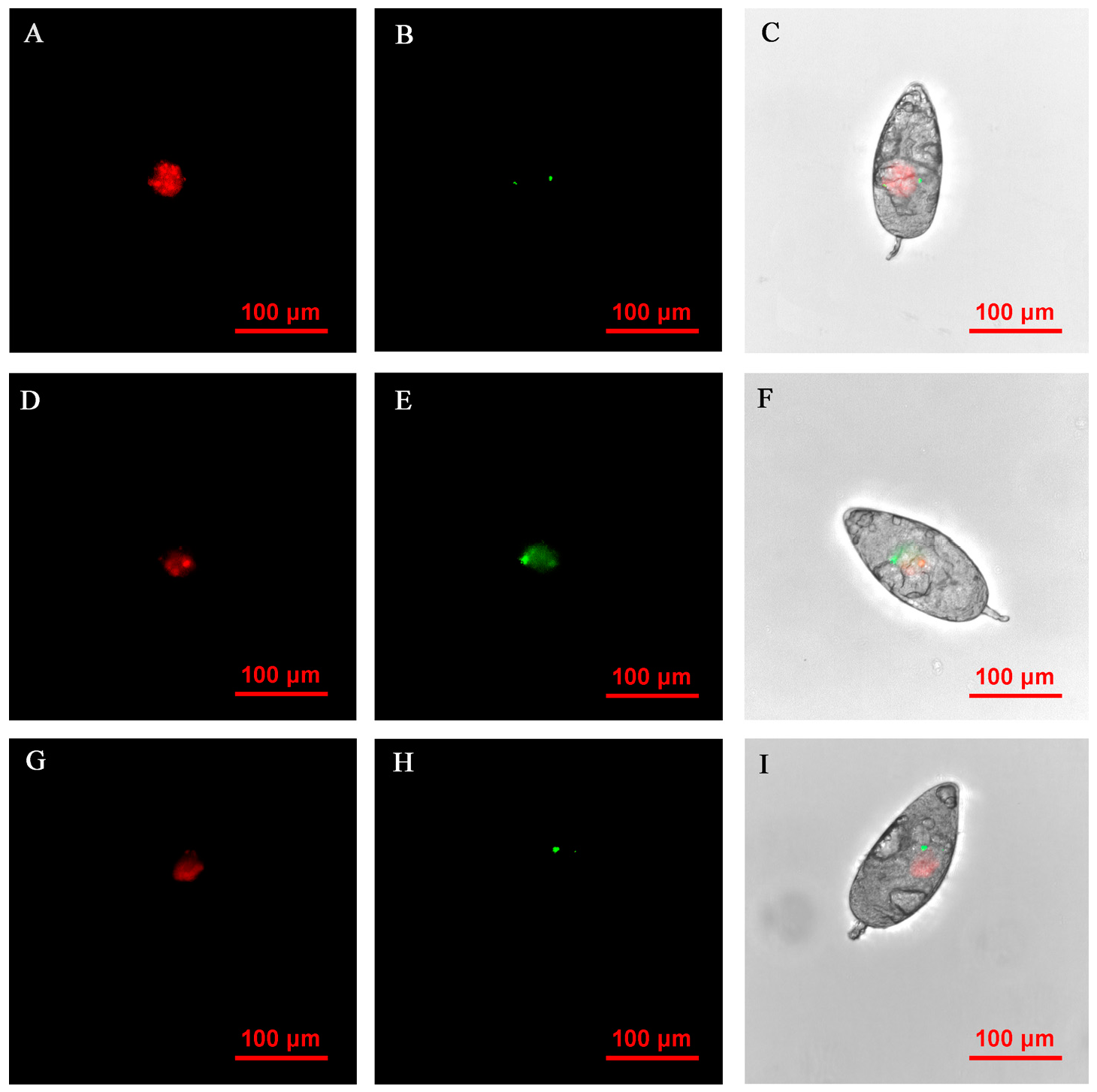
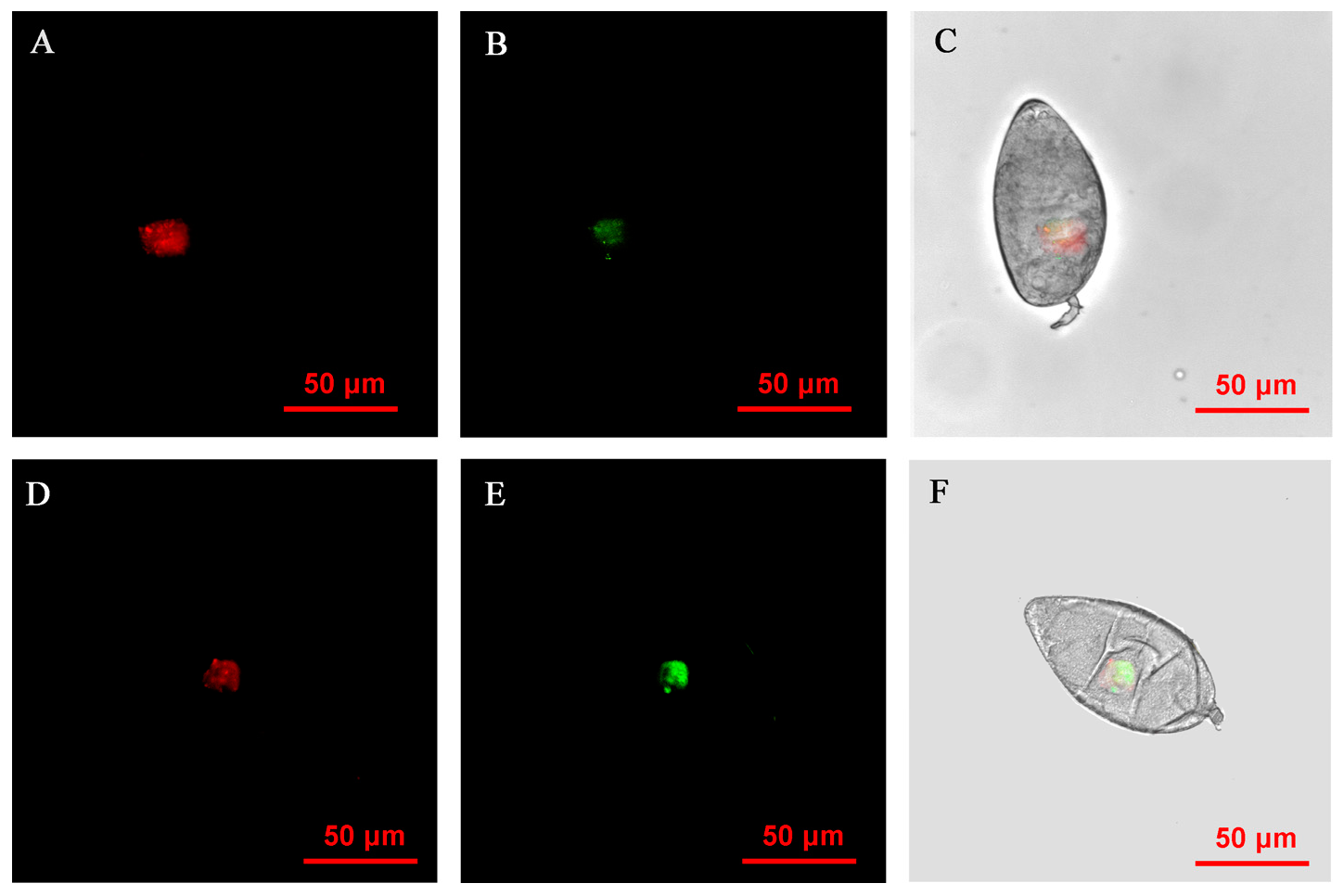
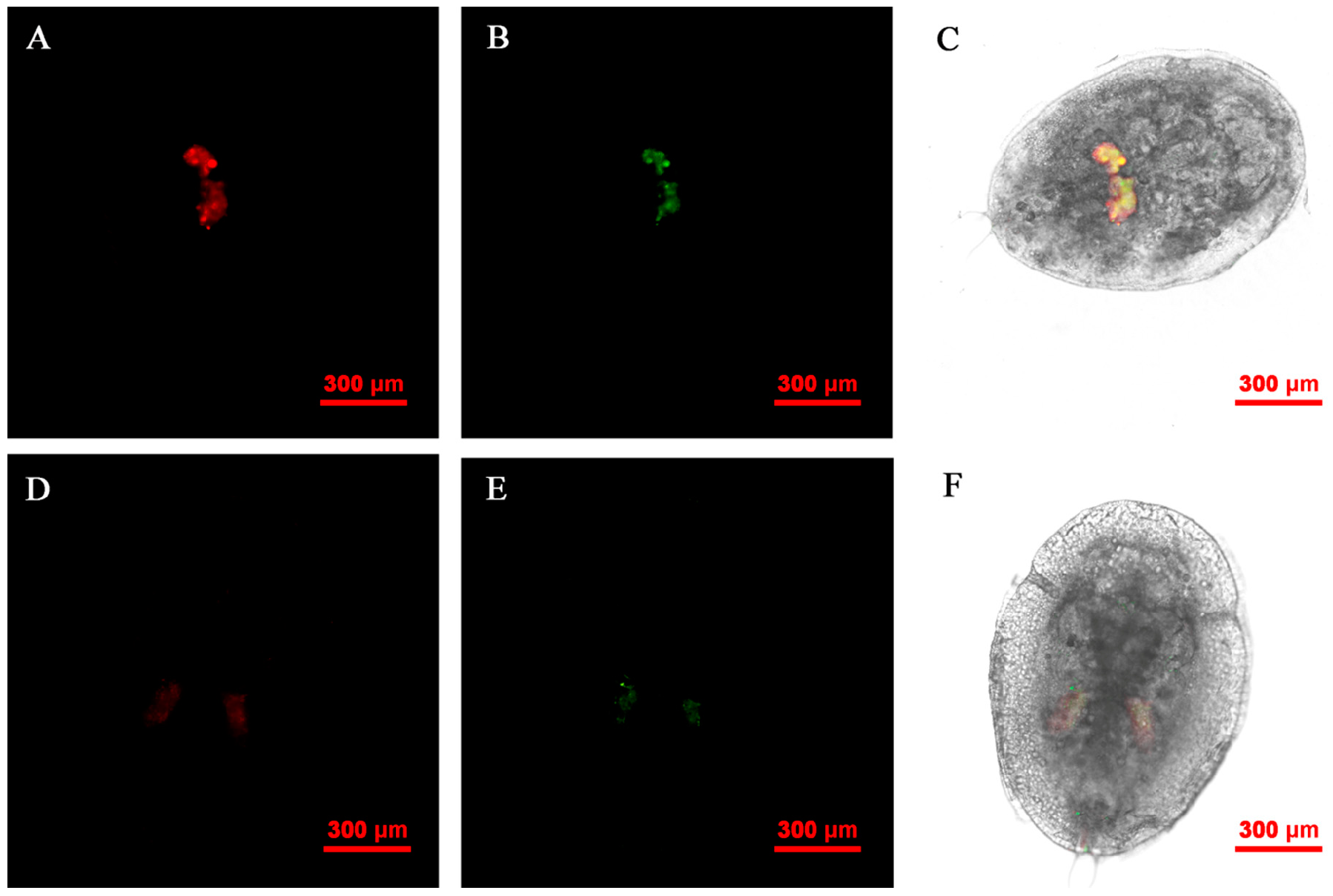
FISH visualization showed that Rickettsia and Wolbachia in IWR eggs of the Asia II1 B. tabaci were mainly distributed in the egg stalk position within 3–9 h, and the fluorescence signal intensity of Rickettsia and Wolbachia becomes weaker and weaker as time goes by (Figure 3). The locations of Rickettsia and Wolbachia were transferred from the egg stalk to the base of the egg, and the fluorescent signals of Rickettsia and Wolbachia in the egg stalk gradually disappeared within 12–24 h (Figure 4). Rickettsia and Wolbachia moved from egg base to the apical portion of the egg within 24–48 h (Figure 5). Finally, Rickettsia and Wolbachia moved back from the apical egg to the basal egg and were fixed in the middle of the egg to complete the transfer process (Figure 6).
3.5. Quantitative Detection of Rickettsia and Wolbachia in Asia II1 Whiteflies
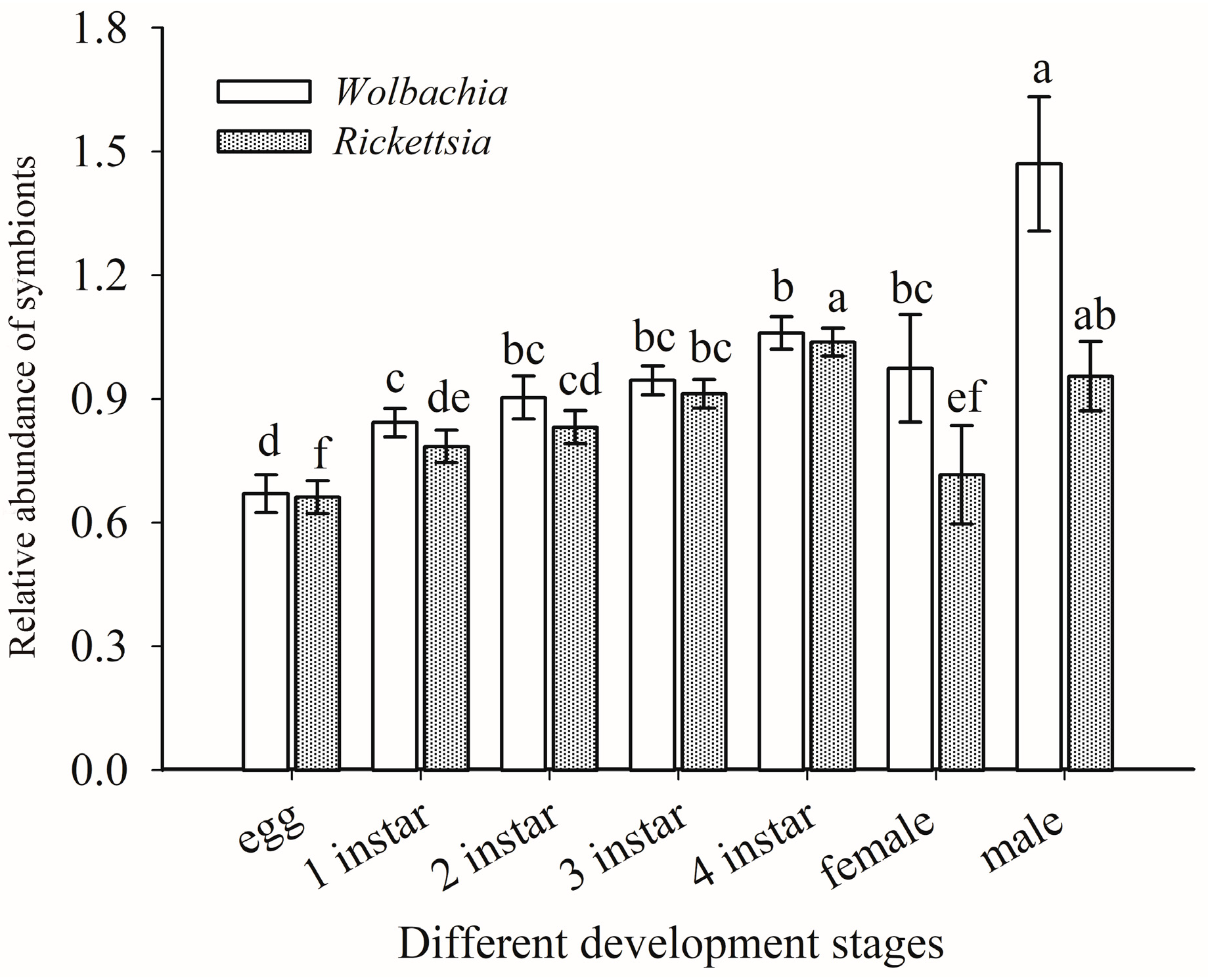
The titers of Rickettsia and Wolbachia nymphal and adult life stages of Asia II1 B. tabaci generally increased with the development of whiteflies. The titers of endosymbionts Rickettsia and Wolbachia in males of Asia II1 B. tabaci was significantly higher than those in females. The highest titer of Wolbachia was observed in male adults, followed by fourth instar larvae (Figure 7). The titers of Rickettsia in the male adults were significantly higher than Wolbachia (t = 7.816, df = 4, p = 0.001), whereas the titer of Rickettsia observed in nymphs were significantly higher than the titer of adult females (Figure 7).
3.6. FISH Visualization of Rickettsia and Wolbachia in Nymphs and Adults
The Rickettsia and Wolbachia of Asia II1 B. tabaci nymph were located by FISH. The results revealed that the presence of Rickettsia and Wolbachia was limited to the bacteriocytes in the abdomen (Figure 8). Rickettsia and Wolbachia can be seen in the center of the abdomen in the adults of Asia II1 B. tabaci. This distribution of endosymbionts is called “the scattered pattern” (Figure 9).
4. Discussion
B. tabaci is a complex composed of at least 40 morphologically indistinguishable species that is known to harbor at least 7 endosymbionts, with infection frequencies varying between cryptic species and conspecific populations [34,35]. In this study, diagnostic PCR screening revealed that the experimental population of B. tabaci Asia II1 included Rickettsia, Wolbachia, Hemipteriphilus, and Fritschea (Figure S2). The species of endosymbionts infecting B. tabaci were affected by many factors, including climate, temperature, and geographical populations. For example, the populations of B. tabaci MEAM1 from Zhejiang and Beijing (China) were infected with two secondary endosymbionts (Rickettsia and Hamiltonella), whereas the populations of B. tabaci MEAM1 from Montenegro also contained a third endosymbiont (Wolbachia). The Guangzhou population of B. tabaci MEAM1 contains Rickettsia, Hamiltonella, and Hemipteriphilus [36]. The above findings suggest that host–endosymbiont interactions may be associated with environmental factors, including climate, geographical factors, and host plant [37].
The vertical transmission of endosymbionts in eggs has been recognized as the main strategy for its persistence in insect hosts, and the vertical transmission mechanism of symbiotic bacteria has been clearly studied [9]; however, the changes in the titers and location of symbiotic bacteria are still unclear. This study indicates that with the increase in egg age, Rickettsia and Wolbachia moved from the egg stalk to the egg, and the content of symbiotic bacteria (Rickettsia and Wolbachia) in eggs increased obviously for the first time at 12 h. The fluorescence intensity and the infection area of symbiotic bacteria in the FISH diagram did not reflect the change in Rickettsia and Wolbachia content at 12–24 h. The infected cell area of symbiotic bacteria increased after the symbiotic bacteria are transferred from the stalk position to the egg base, resulting in a decrease in the content of symbiotic bacteria per unit area, which is best proved by the FISH picture of the original symbiotic bacteria Portiera in the 1-day-old whitefly eggs and the localization of Wolbachia in the 1-day-old eggs of citrus psylla [38,39]. Although the ct value of the reference gene fluctuates in eggs at different times, the non-normalized data and ct-values of the reference gene were displayed in Figure S3, the ct values coxA and gltA gene were presented in Figures S4 and S5, in order to better understand the changing law of symbiotic bacteria. At the same time, the titer change of Rickettsia and Wolbachia can be reasonably explained from the biological point of view (Supplementary File 2). In this study, symbiotic bacteria Wolbachia and Rickettsia are distributed near the base of the egg stalk, which is similar to the localization of Portiera in the cryptophyte of B. tabaci MEAM1 shown by Shan et al. [38]. In addition, in the eggs of Asia II1 B. tabaci, only one cell composition was observed at different times from Wolbachia and Rickettsia, which is similar to the composition of symbiotic bacteria Hamiltonella, Cardinium, and Rickettsia observed in the eggs of different geographical populations of B. tabaci [26,40]. However, these results are different from the two cell structure compositions of Hamiltonella and Arsenophonus in greenhouse whitefly eggs observed by Marisa et al. [40]. These changes in cellular composition can be related to the interaction between different hosts and symbiotic bacteria.
The titers of endosymbionts Rickettsia and Wolbachia in the males of Asia II1 B. tabaci observed during this study were significantly higher than those in the endosymbionts of different nymphal instars and adult females. Furthermore, the titers of endosymbionts Rickettsia and Wolbachia in adult females were significantly lower than fourth instar nymphs. These results are consistent with those of Lv et al. [31], who also observed that Wolbachia titers in Asia II 7 B. tabaci males were higher than in females. The presence of higher Wolbachia titers in males than females can be largely related to their functions [31]. Nguyen et al. [28] revealed that the Wolbachia titers (capable of inducing CI) in thrips were higher in male hosts than in female hosts. The localization of Rickettsia and Wolbachia examined in nymphal instars and adults of Asia II1 B. tabaci by FISH revealed the presence of Rickettsia and Wolbachia in a confined pattern limited to the bacteriocytes localized in the abdomen. These results are consistent with Gottlieb et al. [32], who detected Wolbachia around and inside the bacteriocytes of nymphs and adults; however, in some individuals, the bacterium was also found in the abdomen.
5. Conclusions
In conclusion, these results will improve our understanding of the vertical transmission of symbiotic bacteria in insects and also provide basic information on the quantity and localization of Wolbachia and Rickettsia within different life stages of B. tabaci. These findings will serve as a reference for the interaction between symbiotic bacteria and different tissues of host insects and the horizontal transfer of symbiotics between different hosts for their management through cytoplasmic incompatibility strategy by microinjection technique.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/insects14040401/s1, Figure S1: The endosymbionts information of Asia II1 whitefly B. tabaci; Figure S2: The relative quantity of Portiera in different times eggs of B. tabaci; Figure S3: Ct value of beta-actin gene in whitefly eggs at different time Error bars represent the standard error of the mean; n= 3 biological replicates; Figure S4: Ct value of coxA gene in whitefly eggs at different time Error bars represent the standard error of the mean; n= 3 biological replicates; Figure S5: Ct value of gltA gene in whitefly eggs at different time Error bars represent the standard error of the mean; n= 3 biological replicates; Table S1: The primers and reaction program for PCR detection of endosymbionts in Asia II1 B. tabaci; Table S2: The information of oligonucletide primers used in quantitative PCR in Asia II1 B. tabaci; Supplementary file 1: PCR detection of secondary endosymbionts in Asia II1 whitefly; Supplementary file 2: Biological explanation of dynamic change of symbiotic bacteria titer [26,27,41,42,43,44,45,46].
Author Contributions
N.L., J.P. and Z.-Q.H. carried out the experiments, participated in data analysis; N.L., Q.W. and Z.-Q.S. carried out the statistical analyses; B.-L.Q., C.-Z.L. and N.L. designed the study; N.L., S.A. and B.-L.Q. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Foundation of China to NL, grant number 32060250.
Data Availability Statement
Upon request, the authors can provide the original data used in this paper.
Acknowledgments
The authors thank Yu-Ping Gou for his critical review of this manuscript. We would like to thank all of the participants who volunteered their time in the study. We appreciate very much the valuable comments and suggestion by the reviewers and editors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Weinert, L.A.; Araujo-Jnr, E.V.; Ahmed, M.Z.; Welch, J.J. The incidence of bacterial endosymbionts in terrestrial arthropods. Proc. Biol. Sci. 2015, 282, 20150249. [Google Scholar] [CrossRef]
- Wilson, A.C.C.; Duncan, R.P. Signatures of host/symbiont genome coevolution in insect nutritional endosymbioses. Proc. Natl. Acad. Sci. USA 2015, 112, 10255–10261. [Google Scholar] [CrossRef]
- Morrow, J.L.; Hall, A.A.G.; Riegler, M. Symbionts in waiting: The dynamics of incipient endosymbiont complementation and replacement in minimal bacterial communities of psyllids. Microbiome 2017, 5, 58. [Google Scholar] [CrossRef]
- Lv, N.; Lei, W.; Wen, S.; Liu, C.Z.; Qiu, B.L. Effects of endosymbiont disruption on the nutritional dynamics of the pea aphid Acyrthosiphon pisum. Insects 2018, 9, 161. [Google Scholar] [CrossRef]
- Pan, X.L.; Zhou, G.L.; Wu, J.H.; Bian, G.W.; Lu, P.; Raikhel, A.S. Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2012, 109, 13–14. [Google Scholar] [CrossRef] [PubMed]
- Maire, J.; Vincent-Monégat, C.; Masson, F.; Zaidman-Rémy, A.; Heddi, A. An IMD-like pathway mediates both endosymbiont control and host immunity in the cereal weevil Sitophilus spp. Microbiome 2018, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Ju, J.F.; Bing, X.L.; Zhao, D.S.; Guo, Y.; Xi, Z.Y.; Hoffmann, A.A.; Zhang, K.J.; Huang, H.J.; Gong, J.T.; Zhang, X.; et al. Wolbachia supplement biotin and riboflavin to enhance reproduction in planthoppers. ISME J. 2020, 14, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Luan, J.; Sun, X.; Fei, Z.; Douglas, A.E. Maternal inheritance of a single somatic animal cell displayed by the bacteriocyte in the whitefly Bemisia tabaci. Curr. Biol. 2018, 28, 459–465. [Google Scholar] [CrossRef]
- Shan, H.W.; Liu, Y.Q.; Luan, J.B.; Liu, S.S. New insight into the transovarial transmission of the symbiont Rickettsia in whitefly. Sci. China Life Sci. 2021, 64, 1174–1186. [Google Scholar] [CrossRef]
- De Barro, P.J.; Liu, S.S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A statement of species status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef]
- Mugerwa, H.; Colvin, J.; Alicai, T.; Omongo, A.C.; Kabaalu, P.; Visendi, P. Genetic diversity of whitefly (Bemisia spp.) on crop and uncultivated plants in Uganda: Implications for the control of this devastating pest species complex in Africa. J. Pest. Sci. 2021, 94, 1307–1330. [Google Scholar] [CrossRef]
- Luan, J.B.; Shan, H.W.; Isermann, P.; Huang, J.H.; Lammerding, J.; Liu, S.S.; Douglas, A.E. Cellular and molecular remodelling of a host cell for vertical transmission of bacterial symbionts. Proc. Biol. Sci. 2016, 283, 20160580. [Google Scholar] [CrossRef] [PubMed]
- Perotti, M.A.; Clarke, H.K.; Turner, B.D.; Braig, H.R. Rickettsia as obligate and mycetomic bacteria. FASEB J. 2006, 20, 2372–2374. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.C.; Sakurai, M.; Bonhomme, J.; Tsuchida, T.; Koga, R.; Fukatsu, T. Elimination of a specialized facultative symbiont does not affect the reproductive mode of its aphid host. Ecol. Entomol. 2007, 32, 296–301. [Google Scholar] [CrossRef]
- Kontsedalov, S.E.; Zchori-Fein, E.; Chiel, Y.; Gottlieb, M.; Inbar, M.; Ghanim, M. The presence of Rickettsia is associated with increased susceptibility of Bemisia tabaci (Homoptera: Aleyrodidae) to insecticides. Pest Manag. Sci. 2008, 64, 789–792. [Google Scholar] [CrossRef]
- Zhou, X.F.; Li, Z.X. Establishment of the cytoplasmic incompatibility-inducing Wolbachia strain wMel in an important agricultural pest insect. Sci. Rep. 2016, 6, 39200. [Google Scholar] [CrossRef]
- Giorgini, M.; Bernardo, U.; Monti, M.M.; Nappo, A.G.; Gebiola, M. Rickettsia symbionts cause parthenogenetic reproduction in the parasitoid wasp pnigalio soemius (Hymenoptera: Eulophidae). Appl. Environ. Microbiol. 2010, 76, 2589–2599. [Google Scholar] [CrossRef] [PubMed]
- Bourtzis, K. Wolbachia-based technologies for insect pest population control. Adv. Exp. Med. Biol. 2008, 627, 104–113. [Google Scholar]
- Christodoulou, M. Biological vector control of mosquito-borne diseases. Lancet Infect Dis. 2011, 11, 84–85. [Google Scholar] [CrossRef] [PubMed]
- Hancock, P.A.; Sinkins, S.P.; Godfray, H.C. Strategies for introducing Wolbachia to reduce transmission of mosquito-borne diseases. PLoS Negl. Trop Dis. 2011, 5, e1024. [Google Scholar] [CrossRef]
- Hughes, G.L.; Rasgon, J.L. Transinfection: A method to investigate Wolbachia-host interactions and control arthropd-borne diseases. Insect Mol. Biol. 2014, 23, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Strunov, A.; Kiseleva, E.; Gottlieb, Y. Spatial and temporal distribution of pathogenic Wolbachia strain wMelPop in Drosophila melanogaster central nervous system under different temperature conditions. J. Invertebr. Pathol. 2013, 114, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Li, S.J.; Ahmed, M.; Lv, N.; Shi, P.Q.; Wang, X.M.; Huang, J.L. Plant-mediated horizontal transmission of Wolbachia between whiteflies. ISME J. 2017, 11, 1019–1028. [Google Scholar] [CrossRef]
- Liu, Y.; Fan, Z.Y.; An, X.; Shi, P.Q.; Ahmed, M.Z.; Qiu, B.L. A single-pair method to screen Rickettsia-infected and uninfected whitefly Bemisia tabaci populations. J. Microbiol. Methods 2020, 168, 105795. [Google Scholar] [CrossRef]
- Ahmed, M.Z.; Ren, S.X.; Xue, X.; Li, X.X.; Jin, G.H.; Qiu, B.L. Prevalence of endosymbionts in Bemisia tabaci populations and their in vivo sensitivity to antibiotics. Curr. Microbiol. 2010, 61, 322–328. [Google Scholar] [CrossRef]
- Gottlieb, Y.; Ghanim, M.; Chiel, E.; Gerling, D.; Portnoy, V. Identification and localization of a Rickettsia sp. in Bemisia tabaci (Homoptera: Aleyrodidae). Appl. Environ. Microbiol. 2006, 72, 3646–3652. [Google Scholar] [CrossRef]
- Zhou, W.; Rousset, F.; O’Neill, S. Phylogeny and PCR-based classification of Wolbachia strains using WSP gene sequences. P. Roy. Soc. B-Biol. Sci. 1998, 265, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.T.; Morrow, J.L.; Spooner-Hart, R.N.; Riegler, M. Independent cytoplasmic incompatibility induced by Cardinium and Wolbachia maintains endosymbiont coinfections in haplodiploid thrips populations. Evolution 2017, 71, 995–1008. [Google Scholar] [CrossRef]
- Ghanim, M.; Kontsedalov, S. Susceptibility to insecticides in the Q biotype of Bemisia tabaci is correlated with bacterial symbiont densities. Pest Manag. Sci. 2009, 65, 939–942. [Google Scholar] [CrossRef]
- Pan, H.P.; Chu, D.; Liu, B.M.; Xie, W.; Wang, S.L.; Wu, Q.J.; Xu, Y.X.; Zhang, Y.J. Relative amount of symbionts in insect hosts changes with host-plant adaptation and insecticide resistance. Environ. Entomol. 2013, 42, 74–78. [Google Scholar] [CrossRef]
- Lv, N.; Peng, J.; Chen, X.Y.; Guo, C.F.; Sang, W.; Wnag, X.M. Antagnostic interaction between male-killing and cytoplasmic incompatibility induced by Cardinium and Wolbachia in the whitefly, Bemisia tabaci. Insect Sci. 2021, 28, 330–346. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, Y.; Ghanim, M.; Gueguen, G.; Kontsedalov, S.; Zchori-Fein, E. Inherited intracellular ecosystem: Symbiotic bacteria share bacteriocytes in whiteflies. FASEB J. 2008, 22, 2591–2599. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Liu, S.S.; Colvin, J.; De Barro, P.J. Species concepts as applied to the whitefly Bemisia tabaci systematics: How many species are there? J. Integr. Agr. 2012, 11, 176–186. [Google Scholar] [CrossRef]
- Tang, X.T.; Cai, L.; Shen, Y.; Du, Y.Z. Diversity and evolution of the endosymbionts of Bemisia tabaci in China. Peer J. 2018, 6, e5516. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.Q.; He, Z.; Li, S.j.; An, X.; Lv, N.; Ghanim, M.; Qiu, B.L. Wolbachia has two different localization patterns in whitefly Bemisia tabaci Asia II 7 species. PLoS ONE 2016, 11, e0162558. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.X.; Song, Y.L.; Zhang, Y.K.; Hoffmann, A.A.; Zhou, J.C.; Sun, J.T. Incidence of facultative bacterial endosymbionts in spider mites associated with local environment and host plant. Appl. Environ. Microb. 2018, 84, e02546-17. [Google Scholar] [CrossRef]
- Shan, H.W.; Lu, Y.H.; Bing, X.L.; Liu, S.S.; Liu, Y.Q. Differential responses of the whitefly Bemisia tabaci symbionts to unfavorable low and high temperatures. Microb. Ecol. 2014, 68, 472–482. [Google Scholar] [CrossRef]
- Ren, S.L.; Li, Y.H.; Ou, D.; Guo, Y.J.; Qiu, B.L. Localization and dynamics of Wolbachia infection in Asian citrus psyllid Diaphorina citri, the insect vector of the causal pathogens of Huanglongbing. Microbiol. Open 2018, 7, e00561. [Google Scholar] [CrossRef] [PubMed]
- Marisa, S.; Katja, Z.; Smiljana Goreta, B.; Svetlana, K.; Murad, G. Co-infection and localization of secondary symbionts in two whitefly species. BMC Microbiol. 2010, 10, 142. [Google Scholar]
- Zchori-Fein, E.; Brown, J.K. Diversity of prokaryotes associated with Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). Ann. Entomol. Soc. Am. 2002, 6, 711–718. [Google Scholar] [CrossRef]
- Chiel, E.; Gottlieb, Y.; Zchori-Fein, E.; Mozes-Daube, N.; Katzir, N.; Inbar, M.; Ghanim, M. Biotype-dependent secondary symbiont communities in sympatric populations of Bemisia tabaci. B Entomol. Res. 2007, 97, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Weeks, A.R.; Velten, R.; Stouthamer, R. Incidence of a new sex-ratio-distorting endosymbiotic bacterium among arthropods. Proc. R. Soc. Lond. B. 2003, 270, 1857–1865. [Google Scholar] [CrossRef] [PubMed]
- Thao, M.L.; Baumann, P. Evolutionary relationships of primary prokaryotic endosymbionts of whiteflies and their hosts. Appl. Environ. Microb. 2004, 70, 3401–3406. [Google Scholar] [CrossRef] [PubMed]
- Everett, K.D.E.; Thao, M.L.; Horn, M.; Dyszynski, G.E.; Baumann, P. Novel chlamydiae in whiteflies and scale insects: Endosymbionts ‘Candidatus Fritschea bemisiae’ strain Falk and ‘Candidatus Fritschea eriococci’ strain Elm. Int. J. Syst. Evol. Micr. 2005, 55, 1581–1587. [Google Scholar] [CrossRef] [PubMed]
- Bing, X.L.; Yang, J.; Zchori-Fein, E.; Wang, X.W.; Liu, S.S. Characterization of a newly discovered symbiont of the whitefly bemisia tabaci (Hemiptera: Aleyrodidae). Appl. Environ. Microb. 2012, 79, 569–575. [Google Scholar] [CrossRef]
Figure 1.
PCR detection of Rickettsia and Wolbachia in different Asia II 1 whitefly lines. (A,B): respectively, the PCR detection endosymbiont female and male in the IWR whitefly population; M: DNA marker; Lanes 1: positive control (Portiera); Lanes 2: negative control (ddH2O); Lanes 3: sequence fragments of Wolbachia wsp gene in the IWR whitefly populations; Lanes 4, sequence fragments of Rickettsia 16S rDNA gene in IWR whitefly population.
Figure 1.
PCR detection of Rickettsia and Wolbachia in different Asia II 1 whitefly lines. (A,B): respectively, the PCR detection endosymbiont female and male in the IWR whitefly population; M: DNA marker; Lanes 1: positive control (Portiera); Lanes 2: negative control (ddH2O); Lanes 3: sequence fragments of Wolbachia wsp gene in the IWR whitefly populations; Lanes 4, sequence fragments of Rickettsia 16S rDNA gene in IWR whitefly population.

Figure 2.
The relative abundance of Rickettsia and Wolbachia in B. tabaci eggs at different time intervals: (a) relative abundance of Rickettsia; (b) relative abundance of Wolbachia. n = 3 biological replicates. Error bars represent the standard error of the mean; Means marked with different letters are significantly different from each other (p ≤ 0.05).
Figure 2.
The relative abundance of Rickettsia and Wolbachia in B. tabaci eggs at different time intervals: (a) relative abundance of Rickettsia; (b) relative abundance of Wolbachia. n = 3 biological replicates. Error bars represent the standard error of the mean; Means marked with different letters are significantly different from each other (p ≤ 0.05).

Figure 3.
FISH visualization of Wolbachia and Rickettsia in egg stages (3–9 h) of B. tabaci. Localization of symbiotic bacteria Rickettsia (red) and Wolbachia (green) in the B. tabaci eggs at different times ((A–C): 3 h, (D–F): 6 h, (G–I): 9 h). Rickettsia fluorescence (A,D,G) and Wolbachia fluorescence (B,E,H) are displayed in dark field; Wolbachia and Rickettsia fluorescence (C,F,I) is displayed in bright field.
Figure 3.
FISH visualization of Wolbachia and Rickettsia in egg stages (3–9 h) of B. tabaci. Localization of symbiotic bacteria Rickettsia (red) and Wolbachia (green) in the B. tabaci eggs at different times ((A–C): 3 h, (D–F): 6 h, (G–I): 9 h). Rickettsia fluorescence (A,D,G) and Wolbachia fluorescence (B,E,H) are displayed in dark field; Wolbachia and Rickettsia fluorescence (C,F,I) is displayed in bright field.

Figure 4.
FISH visualization of Wolbachia and Rickettsia in egg stages (12–24 h) of B. tabaci. Localization of symbiotic bacteria Rickettsia (red) and Wolbachia (green) in the B. tabaci eggs at different times ((A–C): 12 h, (D–F): 18 h, (G–I): 24 h). Rickettsia fluorescence (A,D,G) and Wolbachia fluorescence (B,E,H) are displayed in dark field; Wolbachia and Rickettsia fluorescence (C,F,I) is displayed in bright field.
Figure 4.
FISH visualization of Wolbachia and Rickettsia in egg stages (12–24 h) of B. tabaci. Localization of symbiotic bacteria Rickettsia (red) and Wolbachia (green) in the B. tabaci eggs at different times ((A–C): 12 h, (D–F): 18 h, (G–I): 24 h). Rickettsia fluorescence (A,D,G) and Wolbachia fluorescence (B,E,H) are displayed in dark field; Wolbachia and Rickettsia fluorescence (C,F,I) is displayed in bright field.

Figure 5.
FISH visualization of Wolbachia and Rickettsia in egg stages (36–72 h) of B. tabaci. Localization of symbiotic bacteria Rickettsia (red) and Wolbachia (green) in the B. tabaci eggs at different times ((A–C): 36 h, (D–F): 48 h, (G–I): 72 h). Rickettsia fluorescence (A,D,G) and Wolbachia fluorescence (B,E,H) are displayed in dark field; Wolbachia and Rickettsia fluorescence (C,F,I) is displayed in bright field.
Figure 5.
FISH visualization of Wolbachia and Rickettsia in egg stages (36–72 h) of B. tabaci. Localization of symbiotic bacteria Rickettsia (red) and Wolbachia (green) in the B. tabaci eggs at different times ((A–C): 36 h, (D–F): 48 h, (G–I): 72 h). Rickettsia fluorescence (A,D,G) and Wolbachia fluorescence (B,E,H) are displayed in dark field; Wolbachia and Rickettsia fluorescence (C,F,I) is displayed in bright field.

Figure 6.
FISH visualization of Wolbachia and Rickettsia in egg stages (96–120 h) of B. tabaci. Localization of symbiotic bacteria Rickettsia (red) and Wolbachia (green) in the B. tabaci eggs at different times ((A–C): 96 h, (D–F): 120 h). Rickettsia fluorescence (A,D) and Wolbachia fluorescence (B,E) are displayed in dark field; Wolbachia and Rickettsia fluorescence (C,F) is displayed in bright field.
Figure 6.
FISH visualization of Wolbachia and Rickettsia in egg stages (96–120 h) of B. tabaci. Localization of symbiotic bacteria Rickettsia (red) and Wolbachia (green) in the B. tabaci eggs at different times ((A–C): 96 h, (D–F): 120 h). Rickettsia fluorescence (A,D) and Wolbachia fluorescence (B,E) are displayed in dark field; Wolbachia and Rickettsia fluorescence (C,F) is displayed in bright field.

Figure 7.
Relative abundance of Wolbachia and Rickettsia in different life stages of B. tabaci Asia II 1. n = 3 biological replicates. The column and error bars represent the fold change in mean ± SE. Means marked with different letters are significantly different from each other (p ≤ 0.05).
Figure 7.
Relative abundance of Wolbachia and Rickettsia in different life stages of B. tabaci Asia II 1. n = 3 biological replicates. The column and error bars represent the fold change in mean ± SE. Means marked with different letters are significantly different from each other (p ≤ 0.05).

Figure 8.
FISH visualization of Wolbachia and Rickettsia in nymph stages of B. tabaci. Localization of symbiotic bacteria Rickettsia (red) and Wolbachia (green) in nymph stages of B. tabaci. Rickettsia fluorescence (A,D) and Wolbachia fluorescence (B,E) are displayed in dark field; Wolbachia and Rickettsia fluorescence (C,F) is displayed in bright field.
Figure 8.
FISH visualization of Wolbachia and Rickettsia in nymph stages of B. tabaci. Localization of symbiotic bacteria Rickettsia (red) and Wolbachia (green) in nymph stages of B. tabaci. Rickettsia fluorescence (A,D) and Wolbachia fluorescence (B,E) are displayed in dark field; Wolbachia and Rickettsia fluorescence (C,F) is displayed in bright field.

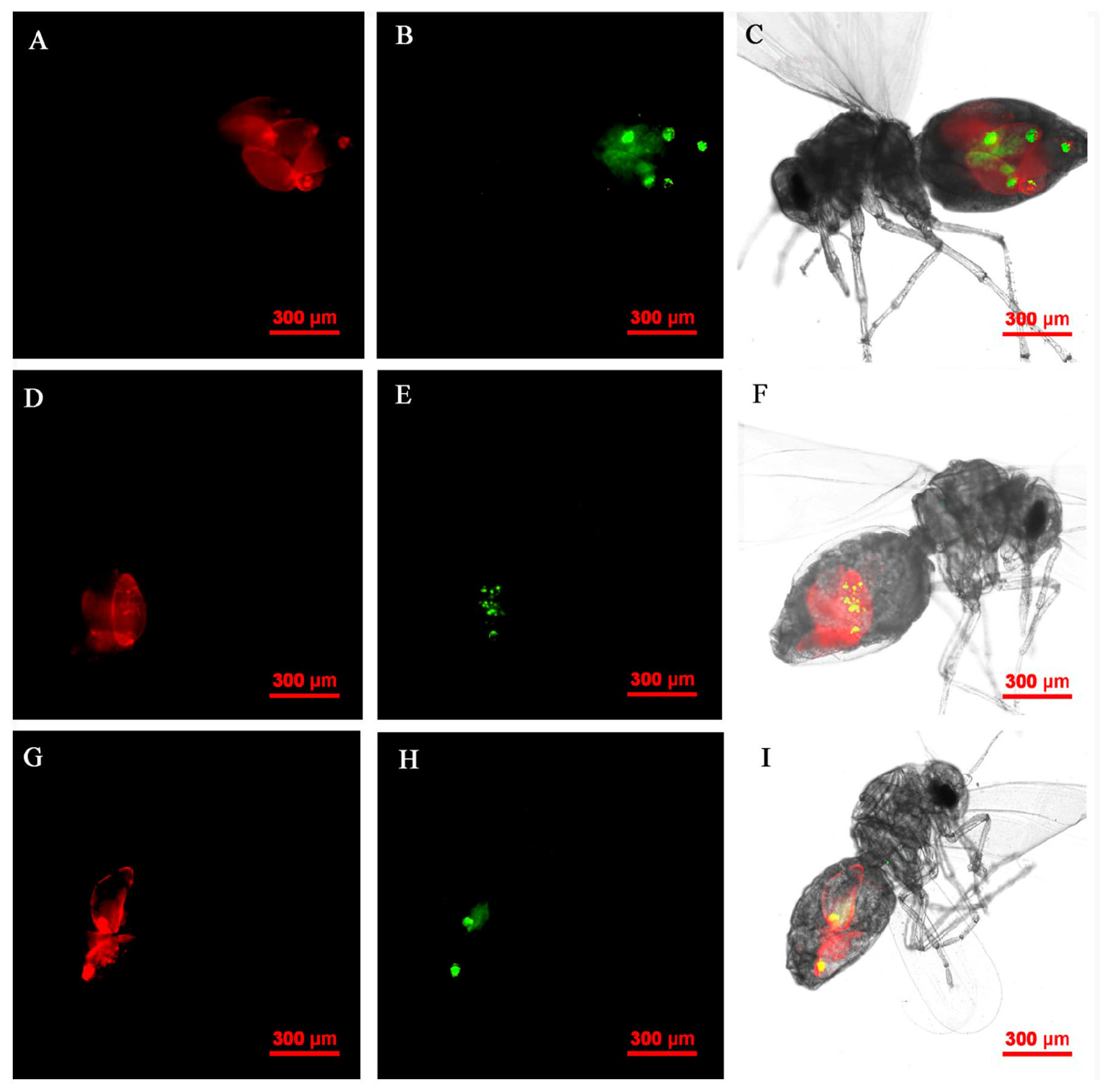
Figure 9.
FISH visualization of Wolbachia and Rickettsia in adult stages of B. tabaci. Localization of symbiotic bacteria Rickettsia (red) and Wolbachia (green) in adult stages of B. tabaci. Rickettsia fluorescence (A,D,G) and Wolbachia fluorescence (B,E,H) are displayed in dark field; Wolbachia and Rickettsia fluorescence (C,F,I) is displayed in bright field.
Figure 9.
FISH visualization of Wolbachia and Rickettsia in adult stages of B. tabaci. Localization of symbiotic bacteria Rickettsia (red) and Wolbachia (green) in adult stages of B. tabaci. Rickettsia fluorescence (A,D,G) and Wolbachia fluorescence (B,E,H) are displayed in dark field; Wolbachia and Rickettsia fluorescence (C,F,I) is displayed in bright field.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Maternal transmission efficiency of Rickettsia and Wolbachia of Asia II1 IWR line.
| Whitefly Samples | n Females | n Offspring Per Female | Total N Tested | Rickettsia | Wolbachia | ||||
|---|---|---|---|---|---|---|---|---|---|
| n+ | n− | % | n+ | n− | % | ||||
| ♀IWR | 10 | 10 | 100 | 100 | 0 | 100 | 100 | 0 | 100 |
| ♀IWR × IWR♂ | 10 | 10 | 100 | 100 | 0 | 100 | 100 | 0 | 100 |
n, numbers of; n+, the numbers of positive individuals; n−, the numbers of negative individuals.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lv, N.; Peng, J.; He, Z.-Q.; Wen, Q.; Su, Z.-Q.; Ali, S.; Liu, C.-Z.; Qiu, B.-L. The Dynamic Distribution of Wolbachia and Rickettsia in AsiaII1 Bemisia tabaci. Insects 2023, 14, 401. https://doi.org/10.3390/insects14040401
AMA Style
Lv N, Peng J, He Z-Q, Wen Q, Su Z-Q, Ali S, Liu C-Z, Qiu B-L. The Dynamic Distribution of Wolbachia and Rickettsia in AsiaII1 Bemisia tabaci. Insects. 2023; 14(4):401. https://doi.org/10.3390/insects14040401
Chicago/Turabian StyleLv, Ning, Jing Peng, Zi-Qi He, Qin Wen, Zheng-Qin Su, Shaukat Ali, Chang-Zhong Liu, and Bao-Li Qiu. 2023. "The Dynamic Distribution of Wolbachia and Rickettsia in AsiaII1 Bemisia tabaci" Insects 14, no. 4: 401. https://doi.org/10.3390/insects14040401
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.