
Nematode and Strepsipteran Parasitism in Bait-Trapped and Hand-Collected Hornets (Hymenoptera, Vespidae, Vespa)
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Dissection of Wasps
2.2. Identification of Parasites
3. Results
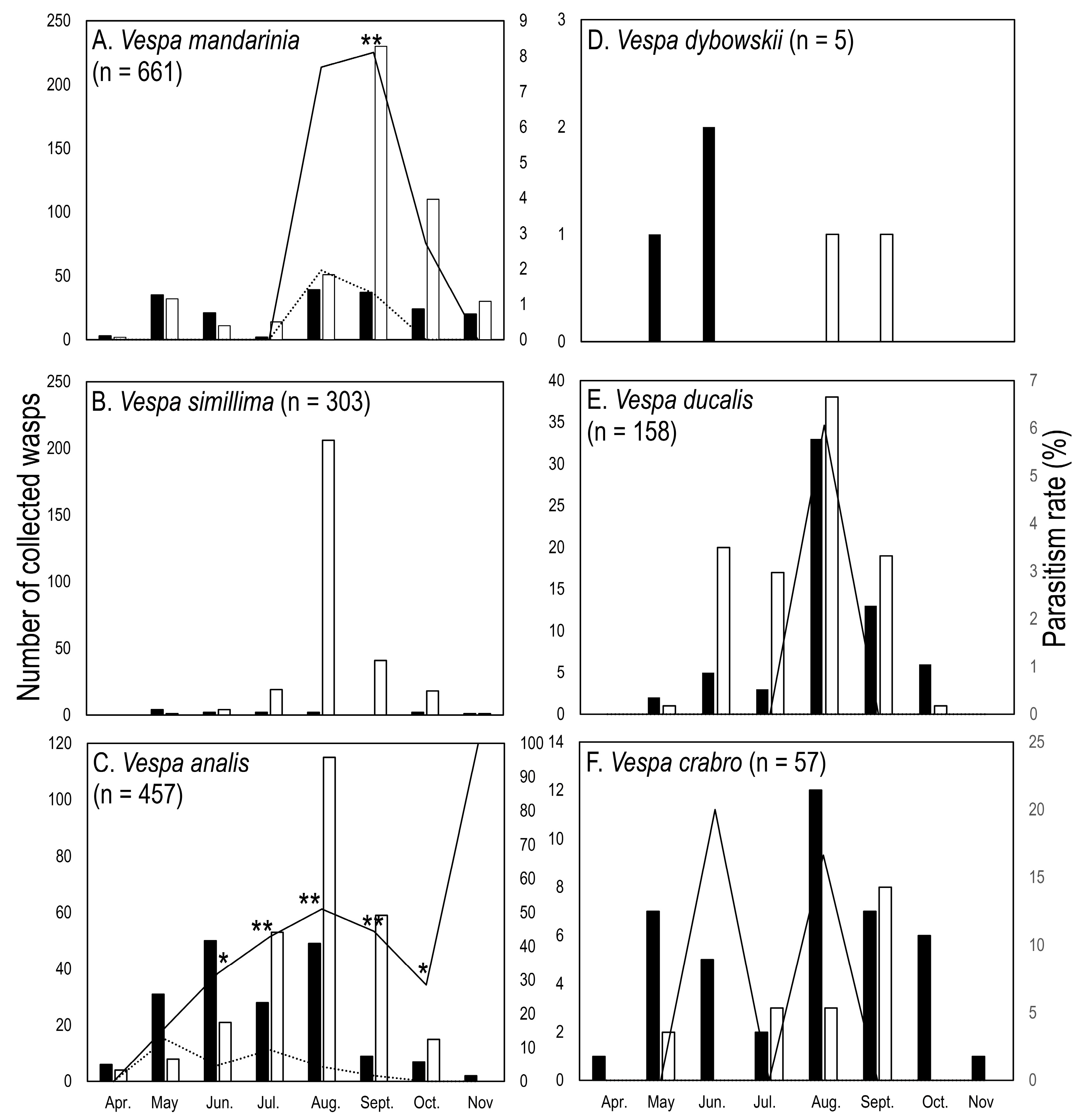
3.1. Hornet Captures
3.2. Nematode Parasitism
3.3. Strepsipteran Parasitism
4. Discussion
4.1. Capture Outcomes of Bait-Trapping Versus Hand-Collecting Hornets
4.2. Comparison of Collection Methods
4.3. Nematode Parasite
4.4. Levels of Xenos Parasitism in Bait-Trapped Hosts
4.5. Number and Sex Ratio of Xenos spp.
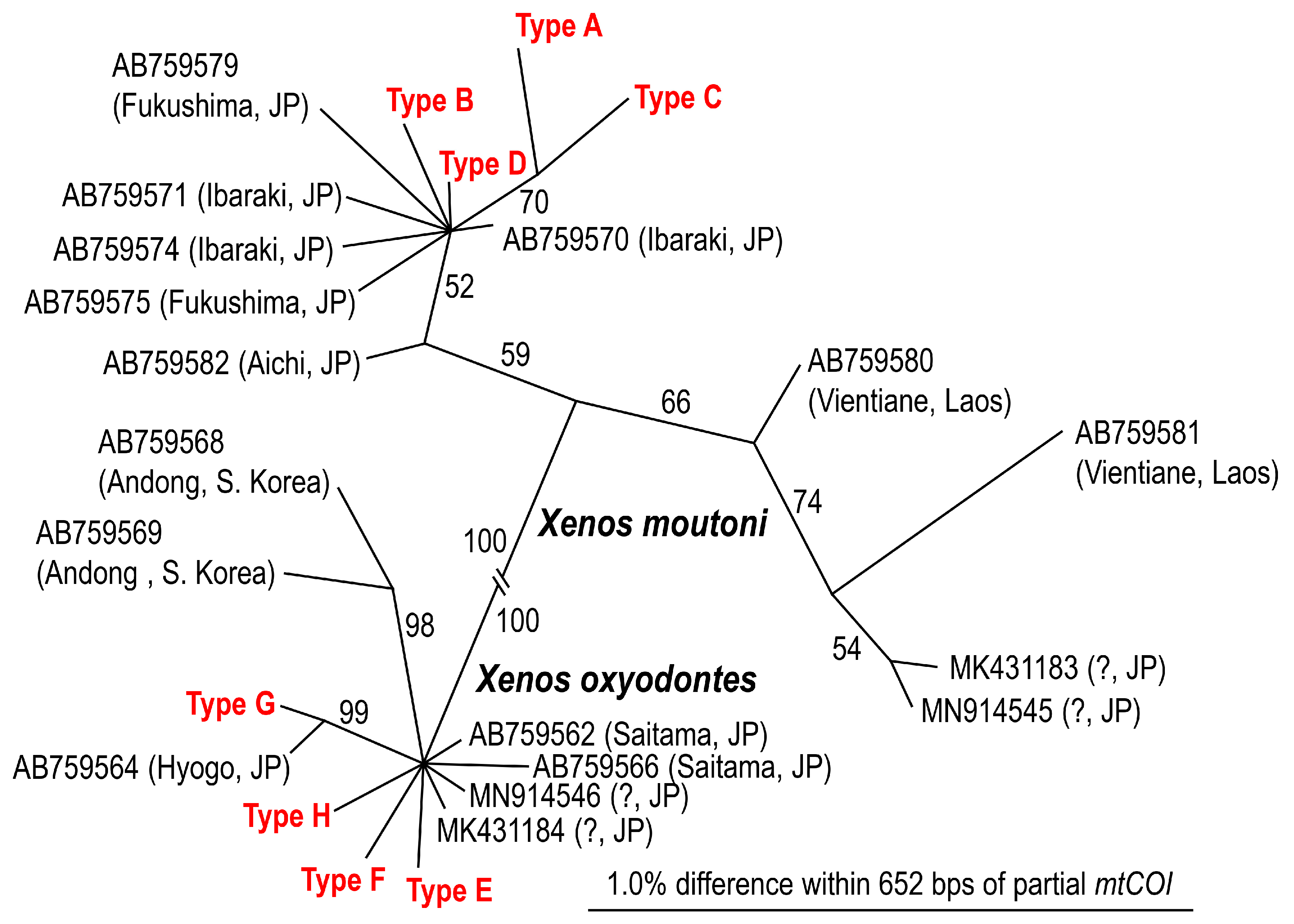
4.6. Molecular Identification and Genotypes of Xenos spp.
4.7. Additional Remarks
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matsuura, M.; Yamane, S.K. Biology of the Vespine Wasps; Springer: Berlin, Germany, 1990; p. 323. [Google Scholar]
- Otaki, N. Allergic symptoms to stings of wasps and bees. In Preventions and Treatments of Stings by Wasps and Bees, 2nd ed.; Matsuura, M., Ed.; Forestry and Timber Manufacturing Safety & Health Association: Tokyo, Japan, 2005; pp. 211–257. (In Japanese) [Google Scholar]
- Budge, G.E.; Hodgetts, J.; Jones, E.P.; Ostojá-Starzewski, J.C.; Hall, J.; Tomkies, V.; Semmence, N.; Brown, M.; Wakefield, M.; Stainton, K. The invasion, provenance and diversity of Vespa velutina Lepeletier (Hymenoptera: Vespidae) in Great Britain. PLoS ONE 2017, 12, e0185172. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Illan, J.G.; Looney, C.; Crowder, D.W. Assessing the ecological niche and invasion potential of the Asian giant hornet. Proc. Natl. Acad. Sci. USA 2020, 117, 24646–24648. [Google Scholar] [CrossRef] [PubMed]
- Poinar, G.O. Entomogenous Nematodes: A Manual and Host List of Insect-Nematode Associations; Brill: Leiden, The Netherlands, 1975; p. 303. [Google Scholar]
- Edwards, R. Social Wasps. Their Biology and Control; Rentokil: East Grinstead, UK, 1980; p. 398. [Google Scholar]
- Ono, M. Science of Japanese Hornets (Vespa spp.); Kaiyusha: Tokyo, Japan, 1997; p. 174. (In Japanese) [Google Scholar]
- Heil, M. Host manipulation by parasites: Cases, patterns, and remaining doubts. Front. Ecol. Evol. 2016, 4, 80. [Google Scholar] [CrossRef]
- Kanzaki, N.; Kosaka, H.; Sayama, K.; Takahashi, J.-I.; Makino, S. Sphaerularia vespae sp. nov. (Nematoda, Tylenchomorpha, Sphaerularioidea), an endoparasite of the common hornet, Vespa simillima Smith (Insecta, Hymenoptera, Vespidae). Zool. Sci. 2007, 24, 1134–1142. [Google Scholar] [CrossRef]
- Sayama, K.; Kosaka, H.; Makino, S. Release of juvenile nematodes at hibernation sites by overwintered queens of the hornet Vespa Simillima. Insectes Soc. 2013, 60, 383–388. [Google Scholar] [CrossRef]
- Poinar, G.O.; Van der Laan, P.A. Morphology and life history of Sphaerularia bombi. Nematologica 1972, 18, 239–252. [Google Scholar] [CrossRef]
- Bedding, R.A. Nematode parasites of Hymenoptera. In Plant and Insect Nematodes; Nickle, W.R., Ed.; Marcel Dekker: New York, NY, USA, 1984; pp. 755–795. [Google Scholar]
- Morooka, F. The prevalence of the parasitic nematode Sphaerularia sp. in the overwintering gynes of Parapolybia spp. (Hymenoptera, Polistinae). J. Hymenopt. Res. 2014, 38, 37–43. [Google Scholar] [CrossRef]
- Kathirithamby, J. Biodiversity of Strepsiptera. In Insect Biodiversity: Science and Society, Vol II. ed.; Foottit, R.G., Adler, P.H., Eds.; John Wiley & Sons: New York, NY, USA, 2018; pp. 673–703. [Google Scholar] [CrossRef]
- Matsuura, M. Social Wasps of Japan in Color; Hokkaido University Press: Sapporo, Japan, 1995; p. 353, (In Japanese with English Summary). [Google Scholar]
- Makino, S.; Yamashita, Y. Levels of parasitism by Xenos moutoni du Buysson (Strepsiptera, Stylopidae) and their seasonal changes in hornets (Hymenoptera: Vespidae, Vespa) caught with bait traps. Entomol. Sci. 1998, 1, 537–543. [Google Scholar]
- Sayama, K. Host utilization and seasonal occurrence of the hornet parasite Xenos moutoni in Hokkaido, northern Japan. Nat. Insects 2012, 47, 21–24. (In Japanese) [Google Scholar]
- Hughes, D.P.; Kathirithamby, J.; Turillazzi, S.; Beani, L. Social wasps desert the colony and aggregate outside if parasitized: Parasite manipulation? Behav. Ecol. 2004, 15, 1037–1043. [Google Scholar] [CrossRef]
- Beani, L.; Cappa, F.; Manfredini, F.; Zaccaroni, M. Preference of Polistes dominula wasps for trumpet creepers when infected by Xenos vesparum: A novel example of co-evolved traits between host and parasite. PLoS ONE 2018, 13, e0205201. [Google Scholar] [CrossRef] [PubMed]
- Beani, L.; Dallai, R.; Cappa, F.; Manfredini, F.; Zaccaroni, M.; Lorenzi, M.C.; Mercati, D. A Strepsipteran parasite extends the lifespan of workers in a social wasp. Sci. Rep. 2021, 11, 7235. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Aikawa, T.; Oeda, Y.; Karim, N.; Kanzaki, N. A rapid and precise diagnostic method for detecting the pinewood nematode Bursaphelenchus xylophilus by loop-mediated isothermal amplification (LAMP). Phytopathology 2009, 99, 1365–1369. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, R.; Kikuchi, T.; Aikawa, T.; Kanzaki, N. Simple and quick methods for nematode DNA preparation. Appl. Entomol. Zool. 2012, 47, 291–294. [Google Scholar] [CrossRef]
- Kanzaki, N.; Masuya, H.; Ichihara, Y.; Maehara, N.; Aikawa, T.; Ekino, T.; Ide, T. Bursaphelenchus carpini n. sp., B. laciniatae n. sp. and B. cryphali okhotskensis n. subsp. (Nematoda: Aphelenchoididae) isolated from Japan. Nematology 2019, 21, 361–388. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 1754–1755. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Larget, B.; Simon, D.L. Markov chain Monte Carlo algorithms for the Bayesian analysis of phylogenetic trees. Mol. Biol. Evol. 1999, 16, 750–759. [Google Scholar] [CrossRef]
- Kelber, A.; Jonsson, F.; Wallén, R.; Warrant, E.; Kornfeldt, T.; Baird, E. Hornets can fly at night without obvious adaptations of eyes and ocelli. PLoS ONE 2011, 6, e21892. [Google Scholar] [CrossRef] [PubMed]
- Makino, S. Seasonal changes in levels of parasitism and sex ratio of Xenos moutoni du Buysson (Strepsiptera, Stylopidae) in the Japanese hornet, Vespa analis insularis Dalla Torre (Hymenoptera, Vespidae), collected with attractant traps. Tijdschr. Voor Entomol. 2001, 144, 217–222. [Google Scholar] [CrossRef]
- Makino, S.; Sayama, K. Species compositions of vespine wasps collected with bait traps in recreation forests in northern and central Japan (Insecta, Hymenoptera, Vespidae). Bull. Forestry For. Prod. Res. Inst. 2005, 4, 283–289. [Google Scholar]
- Oyaizu, W.; Kudô, K. Seasonal changes in the number of vespine wasps and levels of parasitism by Xenos moutoni (Strepsiptera, Stylopidae) collected with attractant traps in Matsunoyama forest, Tokamachi city, Japan. Bull. Facul. Educat. Nat. Sci. Niigata Univ. 2013, 6, 49–57. [Google Scholar]
- Kudô, K.; Oyaizu, W.; Kusama, R.; Yamagishi, K.; Yamaguchi, Y.; Koji, S. Yearly and seasonal changes in species composition of hornets (Hymenoptera: Vespidae) caught with bait traps on the Sea of Japan coast. Far East. Entomol. 2021, 426, 10–18. [Google Scholar] [CrossRef]
- Lioy, S.; Laurino, D.; Capello, M.; Romano, A.; Manino, A.; Porporato, M. Effectiveness and selectiveness of traps and baits for catching the invasive hornet Vespa velutina. Insects 2020, 11, 706. [Google Scholar] [CrossRef]
- Sayama, K.; Kosaka, H.; Makino, S. The first record of infection and sterilization by the nematode Sphaerularia in hornets (Hymenoptera, Vespidae, Vespa). Insectes Soc. 2007, 54, 53–55. [Google Scholar] [CrossRef]
- Kosaka, H.; Sayama, K.; Kanzaki, N.; Takahata, Y.; Makino, S. Host range and geographical distribution of Sphaerularia vespae, the nematode parasite of queen hornets. In Proceedings of the 7th International Congress of Nematology, Antibes Juan-les-Pins, Antibes, France, 1–6 May 2022. [Google Scholar]
- Nakase, Y.; Kato, M. Cryptic diversity and host specificity in giant Xenos strepsipterans parasitic in large Vespa hornets. Zool. Sci. 2013, 30, 331–336. [Google Scholar] [CrossRef]
- Werren, J.H. Sex ratio adaptations to local mate competition in a parasitic wasp. Science 1980, 208, 1157–1159. [Google Scholar] [CrossRef]
- Poulin, R. Population abundance and sex ratio in dioecious helminth parasites. Oecologia 1997, 111, 375–380. [Google Scholar] [CrossRef]
- Carapelli, A.; Vannini, L.; Nardi, F.; Boore, J.L.; Beani, L.; Dallai, R.; Frati, F. The mitochondrial genome of the entomophagous endoparasite Xenos vesparum (Insecta: Strepsiptera). Gene 2006, 376, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Geffre, A.C.; Liu, R.; Manfredini, F.; Beani, L.; Kathirithamby, J.; Grozinger, C.M.; Toth, A.L. Transcriptomics of an extended phenotype: Parasite manipulation of wasp social behaviour shifts expression of caste-related genes. Proc. R. Soc. B 2017, 284, 20170029. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Host Species (Total Number) | Collection Method | Number of Wasps | ||
|---|---|---|---|---|
| Collected | Parasitized by Sphaerularia vespae | Parasitized by Xenos spp. | ||
| Vespa mandarinia (661) | Trap | 181 | 1 (0.6) | 6 (3.3) |
| Net | 480 | 2 (0.4) | 7 (1.5) | |
| Vespa simillima (303) | Trap | 13 | 0 (0.0) | 0 (0.0) |
| Net | 290 | 0 (0.0) | 0 (0.0) | |
| Vespa analis (457) | Trap | 182 | 0 (0.0) | 66 (36.3) |
| Net | 275 | 0 (0.0) | 13 (4.7) | |
| Vespa dybowskii (4) | Trap | 3 | 0 (0.0) | 0 (0.0) |
| Net | 2 | 0 (0.0) | 0 (0.0) | |
| Vespa ducalis (158) | Trap | 62 | 1 (1.6) | 2 (3.2) |
| Net | 96 | 0 (0.0) | 0 (0.0) | |
| Vespa crabro (57) | Trap | 41 | 0 (0.0) | 3 (7.3) |
| Net | 16 | 0 (0.0) | 0 (0.0) | |
| Host Species | Host Indiv. ID | Collection Method | Date |
|---|---|---|---|
| Vespa mandarinia | 2021-0021 | Trap | 4 May |
| 2021-0069 | Net | 18 May | |
| 2021-0122 | Net | 27 May | |
| Vespa ducalis | 2021-0137 | Trap | 31 May |
| (A): Xenos moutoni | ||||||
| Code (Number) | GenBank Accession Number | Host Species | Base Count from 5′ End in 652 bps Segment | |||
| 109 | 112 | 436 | 544 | |||
| A (1) | LC764843 | Vespa mandarinia | C | A | A | A |
| B (1) | LC764844 | Vespa mandarinia | T | T | A | G |
| C (1) | LC764845 | Vespa mandarinia | T | A | G | A |
| D (13) | LC764846 | Vespa mandarinia, V. ducalis, V. crabro | T | A | A | G |
| (B): Xenos oxyodontes | ||||||
| Code (Number) | GenBank Accession Number | Host Species | Base Count from 5′ End in 652 bps Segment | |||
| 286 | 340 | 391 | 463 | |||
| E (1) | LC764847 | Vespa analis | A | T | A | C |
| F (2) | LC764848 | Vespa analis | G | C | A | C |
| G (19) | LC764849 | Vespa analis | G | T | T | C |
| H (30) | LC764850 | Vespa analis | G | T | A | A |
| Host Species (Number of Host Individuals) | Xenos Species | Female | Male | Total | Sex Ratio |
|---|---|---|---|---|---|
| Vespa mandarinia (13) | Xenos moutoni | 1 | 0 | 1 | 100 |
| Vespa analis (77) | Xenos oxyodontes | 0.82 ± 0.74 (0–5) | 0.69 ± 0.98 (0–4) | 1.51 ± 1.25 (1–9) | 54.3 |
| Vespa ducalis (2) | Xenos moutoni | 1.5 ± 0.71 (1–2) | 0 | 1.5 ± 0.71 (1–2) | 100 |
| Vespa crabro (3) | Xenos moutoni | 0.33 ± 0.56 (0–1) | 0.66 ± 0.56 (0–1) | 1 | 33 |
| Species | Host | Sex | May | Jun. | Jul. | Aug. | Sept. | Oct. | Nov. | Total |
|---|---|---|---|---|---|---|---|---|---|---|
| Xenos moutoni | Vespa mandarinia, V. ducalis, V. crabro | Female | 0 | 0 | 0 | 8 | 6 | 3 | 0 | 17 |
| Male | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 2 | ||
| Sex ratio | - | 0 | - | 88 | 100 | 100 | - | 89 | ||
| Xenos oxyodontes | Vespa analis | Female | 6 | 17 | 11 | 24 | 4 | 1 | 0 | 63 |
| Male | 0 | 0 | 11 | 27 | 11 | 2 | 2 | 53 | ||
| Sex ratio | 100 | 100 | 50 | 47 | 27 | 33 | 0 | 54 |
| Host | Expected Xenos Species | Locality (Climate) * | Parasitism (%) | Reference |
|---|---|---|---|---|
| Vespa mandarinia | Xenos moutoni | Hokkaido (SA-CT) | 0 | [17] |
| Niigata (CT) | 16.7–23.5 | [32] | ||
| Ibaraki (T) | 0 | [30] | ||
| Kyoto (T-WT) | 3.3 (2.0) | This study | ||
| Miyazaki (WT) | 3.8 | [16] | ||
| Vespa ducalis | Xenos moutoni | Hokkaido (SA-CT) | Not collected | [17] |
| Niigata (CT) | 12.5 | [32] | ||
| Ibaraki (T) | <2.0 | [30] | ||
| Kyoto (T-WT) | 3.2 (1.3) | This study | ||
| Miyazaki (WT) | 3.8 | [16] | ||
| Vespa dybowskii | Xenos moutoni | Hokkaido (SA-CT) | 0 | [17] |
| Niigata (CT) | Not collected | [32] | ||
| Ibaraki (T) | Not collected | [30] | ||
| Kyoto (T-WT) | 0 (0) | This study | ||
| Miyazaki (WT) | Not collected | [16] | ||
| Vespa crabro | Xenos moutoni | Hokkaido (SA-CT) | <0.5 | [17] |
| Niigata (CT) | 10.1 | [32] | ||
| Ibaraki (T) | <2.0 | [30] | ||
| Kyoto (T-WT) | 7.3 (5.3) | This study | ||
| Miyazaki (WT) | Not collected | [16] | ||
| Vespa simillima | Xenos moutoni or X. oxyodontes | Hokkaido (SA-CT) | <0.5 | [17] |
| Niigata (CT) | 0.4–1.2 | [32] | ||
| Ibaraki (T) | <2.0 | [30] | ||
| Kyoto (T-WT) | 0 (0) | This study | ||
| Miyazaki (WT) | 1.4 | [16] | ||
| Vespa analis | Xenos oxyodontes | Hokkaido (SA-CT) | ca 8.0 | [17] |
| Niigata (CT) | 8.8–16.7 | [32] | ||
| Ibaraki (T) | 15–20 | [30] | ||
| Kyoto (T-WT) | 36.3 (16.8) | This study | ||
| Miyazaki (WT) | 10.6 | [16] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanzaki, N.; Makino, S.; Kosaka, H.; Sayama, K.; Hamaguchi, K.; Narayama, S. Nematode and Strepsipteran Parasitism in Bait-Trapped and Hand-Collected Hornets (Hymenoptera, Vespidae, Vespa). Insects 2023, 14, 398. https://doi.org/10.3390/insects14040398
Kanzaki N, Makino S, Kosaka H, Sayama K, Hamaguchi K, Narayama S. Nematode and Strepsipteran Parasitism in Bait-Trapped and Hand-Collected Hornets (Hymenoptera, Vespidae, Vespa). Insects. 2023; 14(4):398. https://doi.org/10.3390/insects14040398
Chicago/Turabian StyleKanzaki, Natsumi, Shun’ichi Makino, Hajime Kosaka, Katsuhiko Sayama, Keiko Hamaguchi, and Shinji Narayama. 2023. "Nematode and Strepsipteran Parasitism in Bait-Trapped and Hand-Collected Hornets (Hymenoptera, Vespidae, Vespa)" Insects 14, no. 4: 398. https://doi.org/10.3390/insects14040398