


Characterization of Leptoglossus occidentalis Eggs and Egg Glue

 ,
,  , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. On Compounds Released by Insects
1.2. Description and Importance of Leptoglossus occidentalis
1.3. Aim of the Study
2. Material and Methods
2.1. Egg and Egg Glue Samples
2.2. Characterization
3. Results and Discussion
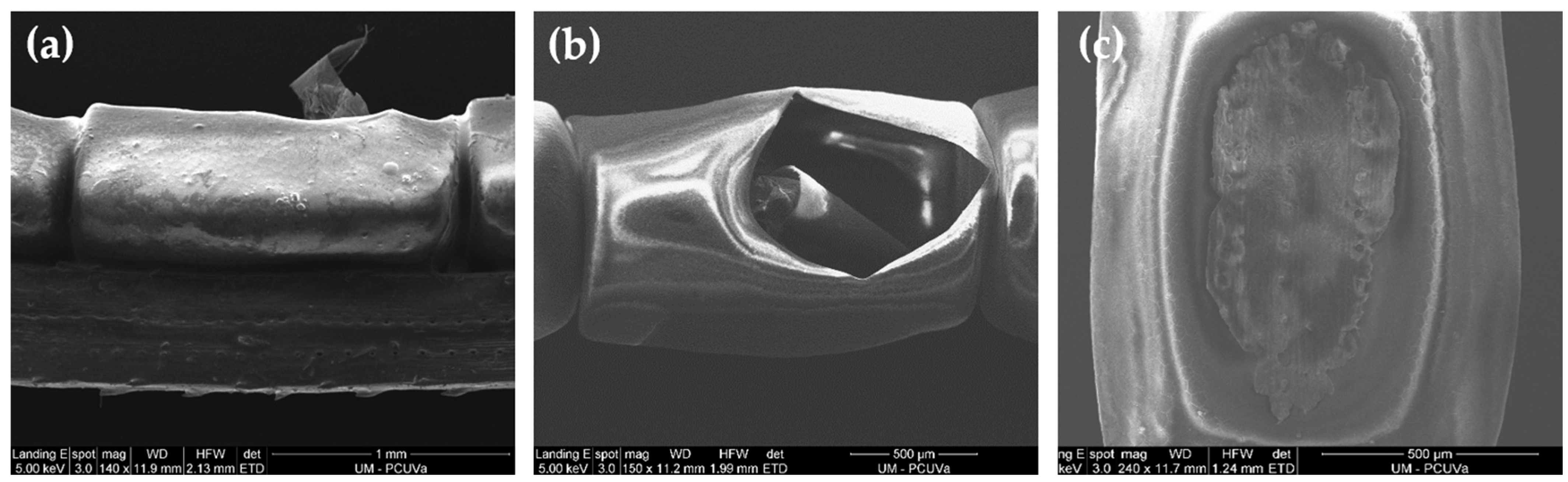
3.1. SEM−EDS Characterization
3.2. Vibrational Characterization of the Eggs and Their Glue

{kind=link}
{kind=link}
| Eggs | Egg Glue | Gum Moth Eggs Glue [4] | Sorghum Bugs Gelatin [29] | Shellac Wax [27] | Saffron [28] | Assignment |
|---|---|---|---|---|---|---|
| 3422 3386 | 3365 | O-H stretching in alcohols | ||||
| 3270 | 3269 | 3274 | 3269 | N-H stretching/typical of geraniol | ||
| 3068 | 3106 | 3081 | hydrocarbon chains in polymeric films/ =C-H stretching in aromatics | |||
| 2965 | 2960 | 2953 | 2955 | C-H methoxyl, cuticles (chitins) | ||
| 2922 | 2918 | 2927 | 2916 | 2924 | CH2– asymmetrical stretching, in butyl itaconate/ COCH3, in chitins/policosanol | |
| 2858 | 2849 | 2850 | 2854 | CH2– symmetrical stretching, in policosanol and cuticles (chitins) | ||
| 1729 | 1736 | 1738 | 1741 | C=O stretching, in policosanol, fatty acids, and lipids/C=C, phorone | ||
| 1634 | 1626 | 1653 | 1633 | 1653 | amide I (C=O of N-acetyl group), in gelatin and chitins | |
| 1547 | 1544 | 1542 | 1543 | C–N stretching/amide II (N-H bend), in chitins and chitosan | ||
| 1515 | 1472 | aromatic squeletal vibrations | ||||
| 1455 | 1463 1445 | 1462 | 1462 | C–H bending (scissor)/aromatic –C=C stretching, in PAla and policosanol | ||
| 1385 | 1377 | 1378 | 1399 | 1374 | 1400 1376 | –OH bending/-C-O-H in-plane bending/–CH3 out-of-plane bending/CH2– wagging and twisting/in chitins |
| 1308 | 1317 | CH3 wagging, in amide III, proteins | ||||
| 1226 | 1245 | 1265 | 1246 | 1227 | C-O-C chitosan/C-O polyols | |
| 1173 | 1173 | C(O)O policosanol | ||||
| 1156 | 1150 | 1148 | 1157 | C-O stretch in C-O-C linkages)/-C-H in chitosan and PAla | ||
| 1115 | 1115 | 1119 | asymmetric in-phase ring stretching | |||
| 1073 | 1065 | 1067 | 1081 | 1075 | C-H bending/C–O–C asym. stretching, in saccharide rings | |
| 1015 | 1028 | 1042 | 1029 | 1020 | C-OH stretching/aromatics, phenols | |
| 953 | 945 | 943 | 945 | CH3 wagging along the chain | ||
| 837 | 862 | trans =C–H out-of-plane bending | ||||
| 759 729 | 759 | 720 | 740 | cis =C-H out-of-plane bending/C=C in aromatic structures |
3.3. GC–MS Characterization of Chemical Species Present in Eggs and Their Glue
4. Discussion
4.1. On Elemental Analysis Results
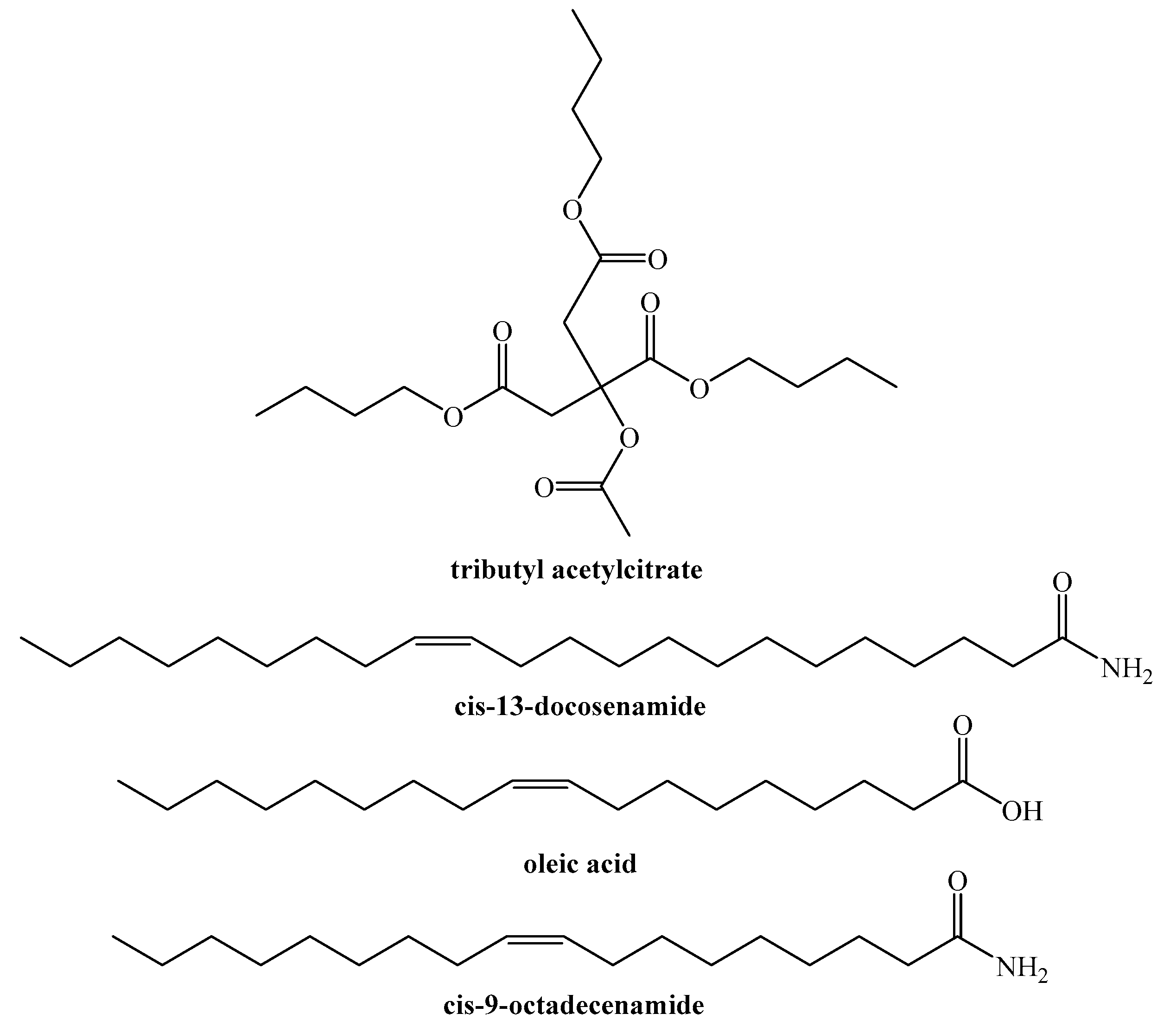
4.2. On Compounds Identified by GC−MS
4.3. Comparison with Pine Constituents
4.4. Usefulness of the Obtained Results
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hilker, M.; Meiners, T. Chemoecology of Insect Eggs and Egg Deposition; Wiley-Blackwell: Berlin, Germany, 2003; p. 416. [Google Scholar] [CrossRef]
- Soldi, R.A.; Rodrigues, M.A.C.M.; Aldrich, J.R.; Zarbin, P.H.G. The male-produced sex pheromone of the true bug, Phthia picta, is an unusual hydrocarbon. J. Chem. Ecol. 2012, 38, 814–824. [Google Scholar] [CrossRef] [PubMed]
- Michereff, M.F.F.; Borges, M.; Aquino, M.F.S.; Laumann, R.A.; Mendes Gomes, A.C.M.; Blassioli-Moraes, M.C. The influence of volatile semiochemicals from stink bug eggs and oviposition-damaged plants on the foraging behaviour of the egg parasitoid Telenomus podisi. Bull. Entomol. Res. 2016, 106, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Huson, M.G.; Graham, L.D. Proteinaceous adhesive secretions from insects, and in particular the egg attachment glue of Opodiphthera sp. moths. Arch. Insect Biochem. Physiol. 2008, 69, 85–105. [Google Scholar] [CrossRef]
- Lei, Y.; Guo, K.; Zhang, Y.; Zhang, X.; Qin, L.; Wang, X.; Zhu, H.; Guo, Y.; Yang, W.; Li, B.; et al. Adhesive property and mechanism of silkworm egg glue protein. Acta Biomater. 2021, 134, 499–512. [Google Scholar] [CrossRef] [PubMed]
- Oliveira Farinha, A.C. Impact and Ecological Adaptation of Leptoglossus occidentalis (Hemiptera, Coreidae) on Pinus pinea; Universidade de Lisboa: Lisbon, Portugal, 2019. [Google Scholar]
- Connelly, A.E.; Schowalter, T.D. Seed losses to feeding by Leptoglossus occidentalis (Heteroptera: Coreidae) during two periods of second-year cone development in western white pine. J. Econ. Entomol. 1991, 84, 215–217. [Google Scholar] [CrossRef]
- Bates, S.L.; Borden, J.H. Life table for Leptoglossus occidentalis Heidemann (Heteroptera: Coreidae) and prediction of damage in lodgepole pine seed orchards. Agric. For. Entomol. 2005, 7, 145–151. [Google Scholar] [CrossRef]
- Mutke, S. Leptoglossus, la chinche del piñón en el mundo y cómo afecta al piñón mediterráneo. Forestalis 2019, 32, 26–29. [Google Scholar]
- Ponce Herrero, L.; Domínguez Alonso, J.C. El chinche de las piñas Leptoglossus occidentalis Heidemann. Rev. For. 2020, 79, 90–99. [Google Scholar]
- Barta, M. Biology and temperature requirements of the invasive seed bug Leptoglossus occidentalis (Heteroptera: Coreidae) in Europe. J. Pest Sci. 2015, 89, 31–44. [Google Scholar] [CrossRef]
- Mas, H.; Naya, M.; Pérez-Laorga, E.; Aguado, A.; Marco, M.; Aragoneses, J.; Rodrigo, E. Estudio del ciclo biológico de Leptoglossus occidentalis Heidemann, 1910 (Hemiptera, Coreidae) en la Comunitat Valenciana. In Proceedings of the 6th Spanish Forest Congress, Vitoria, Spain, 10–14 June 2013; pp. 10–14. [Google Scholar]
- Bernardinelli, I.; Zandigiacomo, P. Leptoglossus occidentalis Heidemann (Heteroptera, Coreidae): A conifer seed bug recently found in northern Italy. J. For. Sci. 2001, 47, 56–58. [Google Scholar]
- Naves, P.; Nóbrega, F.; de Sousa, E. An adult Leptoglossus occidentalis (Heteroptera: Coreidae) parasitized in Portugal by the native Tachinid fly Elomya lateralis (Diptera: Tachinidae). Ann. Soc. Entomol. Fr. (N.S.) 2022, 58, 287–291. [Google Scholar] [CrossRef]
- Strong, W. BC Cone and Seed Pest Research Program; Research and Knowledge Management Branch Seminar Series; Ministry of Forests and Range: Vernon, BC, Canada, 2010. [Google Scholar]
- Blatt, S. An unusually large aggregation of the western conifer seed bug, Leptoglossus occidentalis (Hemiptera: Coreidae), in a man-made structure. J. Entomol. Soc. B. C. 1994, 91, 71–72. [Google Scholar]
- Blatt, S.E.; Borden, J.H. Evidence for a male-produced aggregation pheromone in the western conifer seed bug, Leptoglossus occidentalis Heidemann (Hemiptera: Coreidae). Can. Entomol. 1996, 128, 777–778. [Google Scholar] [CrossRef]
- Blatt, S.E.; Borden, J.H.; Pierce, J.H.D.; Gries, R.; Gries, G. Alarm pheromone system of the western conifer seed bug, Leptoglossus occidentalis. J. Chem. Ecol. 1998, 24, 1013–1031. [Google Scholar] [CrossRef]
- Millar, J.G.; Zou, Y.; Hall, D.R.; Halloran, S.; Pajares, J.A.; Ponce-Herrero, L.; Shates, T.; Wilson, H.; Daane, K.M. Identification and synthesis of Leptotriene, a unique sesquiterpene hydrocarbon from males of the leaf-footed bugs Leptoglossus zonatus and L. occidentalis. J. Nat. Prod. 2022, 85, 2062–2070. [Google Scholar] [CrossRef]
- Takács, S.; Bottomley, H.; Andreller, I.; Zaradnik, T.; Schwarz, J.; Bennett, R.; Strong, W.; Gries, G. Infrared radiation from hot cones on cool conifers attracts seed-feeding insects. Proc. R. Soc. B Biol. Sci. 2008, 276, 649–655. [Google Scholar] [CrossRef]
- Franco-Archundia, S.; Gonzaga-Segura, A.; Jiménez-Pérez, A.; Castrejón-Gómez, V. Behavioral response of Leptoglossus zonatus (Heteroptera: Coreidae) to stimuli based on colors and its aggregation pheromone. Insects 2018, 9, 91. [Google Scholar] [CrossRef]
- Wilson, H.; Maccaro, J.; Daane, K. Optimizing trap characteristics to monitor the leaf-footed bug Leptoglossus zonatus (Heteroptera: Coreidae) in orchards. Insects 2020, 11, 358. [Google Scholar] [CrossRef]
- Inoue, K.; Vidal, D.; Saad, E.; Martins, C.; Zarbin, P. Identification of the alarm and sex pheromones of the leaf-footed bug, Leptoglossus zonatus (Heteroptera: Coreidae). J. Braz. Chem. Soc. 2018, 30, 939–947. [Google Scholar] [CrossRef]
- Mitchell, P.L. Leaf-footed bugs (Coreidae). In Heteroptera of Economic Importance; Schaefer, C.W., Panizzi, A.R., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 337–403. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, D.E.; Park, S.; Clark, J.M.; Lee, S.H. Characterization of nit sheath protein functions and transglutaminase-mediated cross-linking in the human head louse, Pediculus humanus capitis. Parasites Vectors 2021, 14, 425. [Google Scholar] [CrossRef]
- Ma, J.; Ma, L.; Zhang, H.; Zhang, Z.; Wang, Y.; Li, K.; Chen, X. Policosanol fabrication from insect wax and optimization by response surface methodology. PLoS ONE 2018, 13, e0197343. [Google Scholar] [CrossRef]
- Yan, G.; Cao, Z.; Devine, D.; Penning, M.; Gately, N.M. Physical properties of shellac material used for hot melt extrusion with potential application in the pharmaceutical industry. Polymers 2021, 13, 3723. [Google Scholar] [CrossRef]
- Andronie, L.; Coroian, A.; Miresan, V.; Pop, I.; Raducu, C.; Rotaru, A.; Olar, L. Results obtained by investigating saffron ussing FT-IR spectroscopy. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca Anim. Sci. Biotechnol. 2016, 73, 238. [Google Scholar] [CrossRef] [PubMed]
- Mariod, A.A.; Fadul, H. Extraction and characterization of gelatin from two edible Sudanese insects and its applications in ice cream making. Food Sci. Technol. Int. 2015, 21, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Venditti, A. What is and what should never be: Artifacts, improbable phytochemicals, contaminants and natural products. Nat. Prod. Res. 2018, 34, 1014–1031. [Google Scholar] [CrossRef] [PubMed]
- Chaos, A.; Sangroniz, A.; Gonzalez, A.; Iriarte, M.; Sarasua, J.-R.; del Río, J.; Etxeberria, A. Tributyl citrate as an effective plasticizer for biodegradable polymers: Effect of plasticizer on free volume and transport and mechanical properties. Polym. Int. 2019, 68, 125–133. [Google Scholar] [CrossRef]
- Tsochatzis, E.D.; Alberto Lopes, J.; Hoekstra, E.; Emons, H. Development and validation of a multi-analyte GC-MS method for the determination of 84 substances from plastic food contact materials. Anal. Bioanal. Chem. 2020, 412, 5419–5434. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, S.; Yang, Z.; Wang, F.; Deng, L. A novel environment-friendly synthetic technology of dibutyl itaconate. J. Chem. Technol. Biotechnol. 2020, 95, 2879–2885. [Google Scholar] [CrossRef]
- Renthal, R.; Lohmeyer, K.; Borges, L.M.F.; Pérez de León, A.A. Surface lipidome of the lone star tick, Amblyomma americanum, provides leads on semiochemicals and lipid metabolism. Ticks Tick-Borne Dis. 2019, 10, 138–145. [Google Scholar] [CrossRef]
- Jiang, X.-C.; Wen-Xia, D.; Chen, B.; Xiao, C.; Gui, F.-R.; Nai-Sheng, Y.A.N.; Qian, L.; Li, Z.-Y. Electrophysiological and oviposition responses of Asian corn borer, Ostrinia furnacalis (Lepidoptera: Crambidae), to compounds rinsed from the surfaces of sugarcane and maize leaves. Eur. J. Entomol. 2015, 112, 295–301. [Google Scholar] [CrossRef]
- McAfee, A.; Chapman, A.; Iovinella, I.; Gallagher-Kurtzke, Y.; Collins, T.F.; Higo, H.; Madilao, L.L.; Pelosi, P.; Foster, L.J. A death pheromone, oleic acid, triggers hygienic behavior in honey bees (Apis mellifera L.). Sci. Rep. 2018, 8, 5719. [Google Scholar] [CrossRef]
- Chow, Y.S.; Tsai, R.S. Protective chemicals in caterpillar survival. Experientia 1989, 45, 390–392. [Google Scholar] [CrossRef]
- Honda, K. Osmeterial secretions of papilionid larvae in the genera Luehdorfia, Graphium and Atrophaneura (lepidoptera). Insect Biochem. 1980, 10, 583–588. [Google Scholar] [CrossRef]
- Aldrich, J.R.; Schaefer, P.W.; Oliver, J.E.; Puapoomchareon, P.; Lee, C.-J.; Meer, R.K.V. Biochemistry of the exocrine secretion from gypsy moth caterpillars (Lepidoptera: Lymantriidae). Ann. Entomol. Soc. Am. 1997, 90, 75–82. [Google Scholar] [CrossRef]
- Murphy, M.P.; O’Neill, L.A.J. Krebs cycle reimagined: The emerging roles of succinate and itaconate as signal transducers. Cell 2018, 174, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Xu, B. Characterization of essential oil in pine nut shells from commodity waste in China by steam distillation and GC-MS. Food Anal. Methods 2011, 5, 435–440. [Google Scholar] [CrossRef]
- Kadri, N.; Khettal, B.; Aid, Y.; Kherfellah, S.; Sobhi, W.; Barragan-Montero, V. Some physicochemical characteristics of pinus (Pinus halepensis Mill., Pinus pinea L., Pinus pinaster and Pinus canariensis) seeds from North Algeria, their lipid profiles and volatile contents. Food Chem. 2015, 188, 184–192. [Google Scholar] [CrossRef] [PubMed]
- El Omari, N.; Ezzahrae Guaouguaou, F.; El Menyiy, N.; Benali, T.; Aanniz, T.; Chamkhi, I.; Balahbib, A.; Taha, D.; Shariati, M.A.; Zengin, G.; et al. Phytochemical and biological activities of Pinus halepensis mill., and their ethnomedicinal use. J. Ethnopharmacol. 2021, 268, 113661. [Google Scholar] [CrossRef]
- Mohamed, A.A.; Behiry, S.I.; Ali, H.M.; El-Hefny, M.; Salem, M.Z.M.; Ashmawy, N.A. Phytochemical compounds of branches from P. halepensis oily liquid extract and S. terebinthifolius essential oil and their potential antifungal activity. Processes 2020, 8, 330. [Google Scholar] [CrossRef]
- Dhibi, M.; Mechri, B.; Brahmi, F.; Skhiri, F.; Alsaif, M.A.; Hammami, M. Fatty acid profiles, antioxidant compounds and antiradical properties of Pinus halepensis Mill. cones and seeds. J. Sci. Food Agric. 2012, 92, 1702–1708. [Google Scholar] [CrossRef]
- Anttonen, S.; Herranen, J.; Peura, P.; Kärenlampi, L. Fatty acids and ultrastructure of ozone-exposed Aleppo pine (Pinus halepensis Mill.) needles. Environ. Pollut. 1995, 87, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.E.; Pereira, R.M.; Koehler, P.G. Complications with controlling insect eggs. In Insecticides Resistance; Trdan, S., Ed.; InTechOpen: London, UK, 2016; pp. 83–96. [Google Scholar] [CrossRef]
- Bruns, F.H.; Werners, P.H. Dehydrogenases: Glucose-6-phosphate dehydrogenase, 6-phosphogluconate dehydrogenase, glutathione reductase, methemoglobin reductase, polyol dehydrogenases. In Advances in Clinical Chemistry; Elsevier: Amsterdam, The Netherlands, 1963; Volume 5, pp. 237–294. [Google Scholar] [CrossRef]


| C | N | O | S | Si | C/N | C/O | N/O | CxNyOz | |
|---|---|---|---|---|---|---|---|---|---|
| Eggs (inner surface) | 56.8 | 25.1 | 18.0 | - | - | 2.27 | 3.15 | 1.40 | C5N2O |
| Eggs (outer surface) | 72.0 | 9.7 | 18.3 | - | - | 7.42 | 5.57 | 0.53 | C10NO2 |
| Eggs glue | 65.5 | 15.0 | 19.5 | - | - | 4.36 | 3.36 | 0.77 | C10N2O2 or CxNO |
| Gelatin | 50.3 | 17.8 | 25.6 | - | - | 2.82 | 1.97 | 0.69 | C102[H151]N31O39 |
| Lignin | 46.7 | 9.1 | 27.6 | 1.4 | - | 5.13 | 1.69 | 0.33 | C18[H13]N3O8S2 |
| Arginine | 41.4 | 32.2 | 18.4 | - | - | 1.28 | 2.25 | 1.75 | C6[H14]N4O2 |
| Chitin | 47.2 | 6.9 | 39.4 | - | - | 6.84 | 1.20 | 0.18 | C6[H14]NO5 |
| 4-Hydroxyproline | 45.8 | 10.7 | 35.1 | - | - | 4.28 | 1.31 | 0.30 | C5[H9]NO3 |
| Shellac | 61.4 | - | 30.0 | - | - | - | 2.04 | - | C30[H50]O11 |
| Hexadecanoic acid | 75.0 | - | 12.5 | - | - | - | 6.00 | - | C16[H32]O2 |
| Oleic acid | 76.6 | - | 14.8 | - | - | - | 5.17 | - | C18[H34]O2 |
| Pine needle | 67.0 | - | 28.5 | - | 4.5 | - | 2.35 | - | C10[H11]O3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Hernández, E.; Martín-Ramos, P.; Niño-Sánchez, J.; Diez-Hermano, S.; Álvarez-Taboada, F.; Pérez-García, R.; Santiago-Aliste, A.; Martín-Gil, J.; Diez-Casero, J.J. Characterization of Leptoglossus occidentalis Eggs and Egg Glue. Insects 2023, 14, 396. https://doi.org/10.3390/insects14040396
Sánchez-Hernández E, Martín-Ramos P, Niño-Sánchez J, Diez-Hermano S, Álvarez-Taboada F, Pérez-García R, Santiago-Aliste A, Martín-Gil J, Diez-Casero JJ. Characterization of Leptoglossus occidentalis Eggs and Egg Glue. Insects. 2023; 14(4):396. https://doi.org/10.3390/insects14040396
Chicago/Turabian StyleSánchez-Hernández, Eva, Pablo Martín-Ramos, Jonatan Niño-Sánchez, Sergio Diez-Hermano, Flor Álvarez-Taboada, Rodrigo Pérez-García, Alberto Santiago-Aliste, Jesús Martín-Gil, and Julio Javier Diez-Casero. 2023. "Characterization of Leptoglossus occidentalis Eggs and Egg Glue" Insects 14, no. 4: 396. https://doi.org/10.3390/insects14040396