
Factors Affecting Population Dynamics of Helicoverpa zea (Lepidoptera: Noctuidae) in a Mixed Landscape with Bt Cotton and Peanut
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Pheromone Trapping and Insect Collection
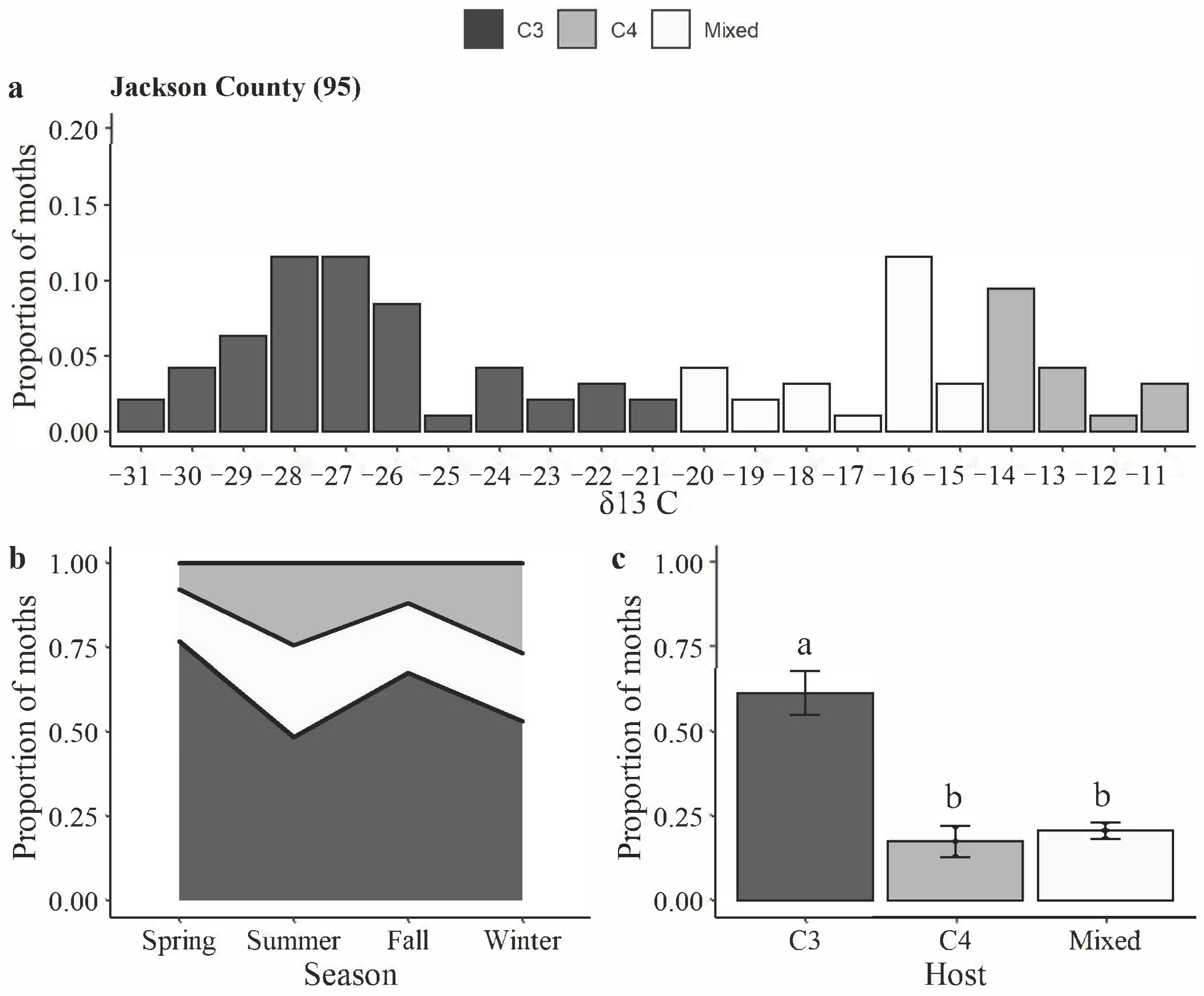
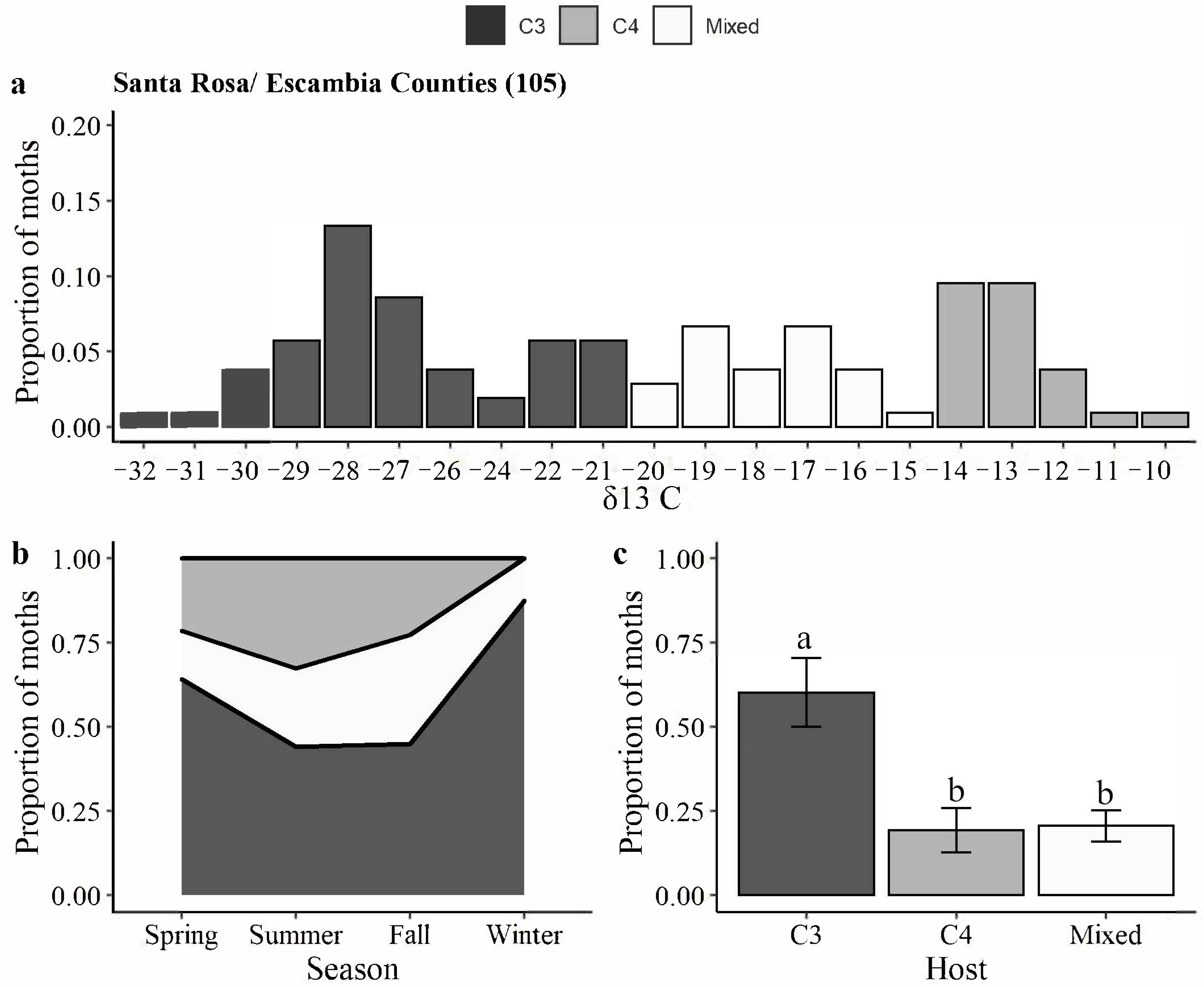
2.2. Carbon Isotopic Signatures of H. zea Moths
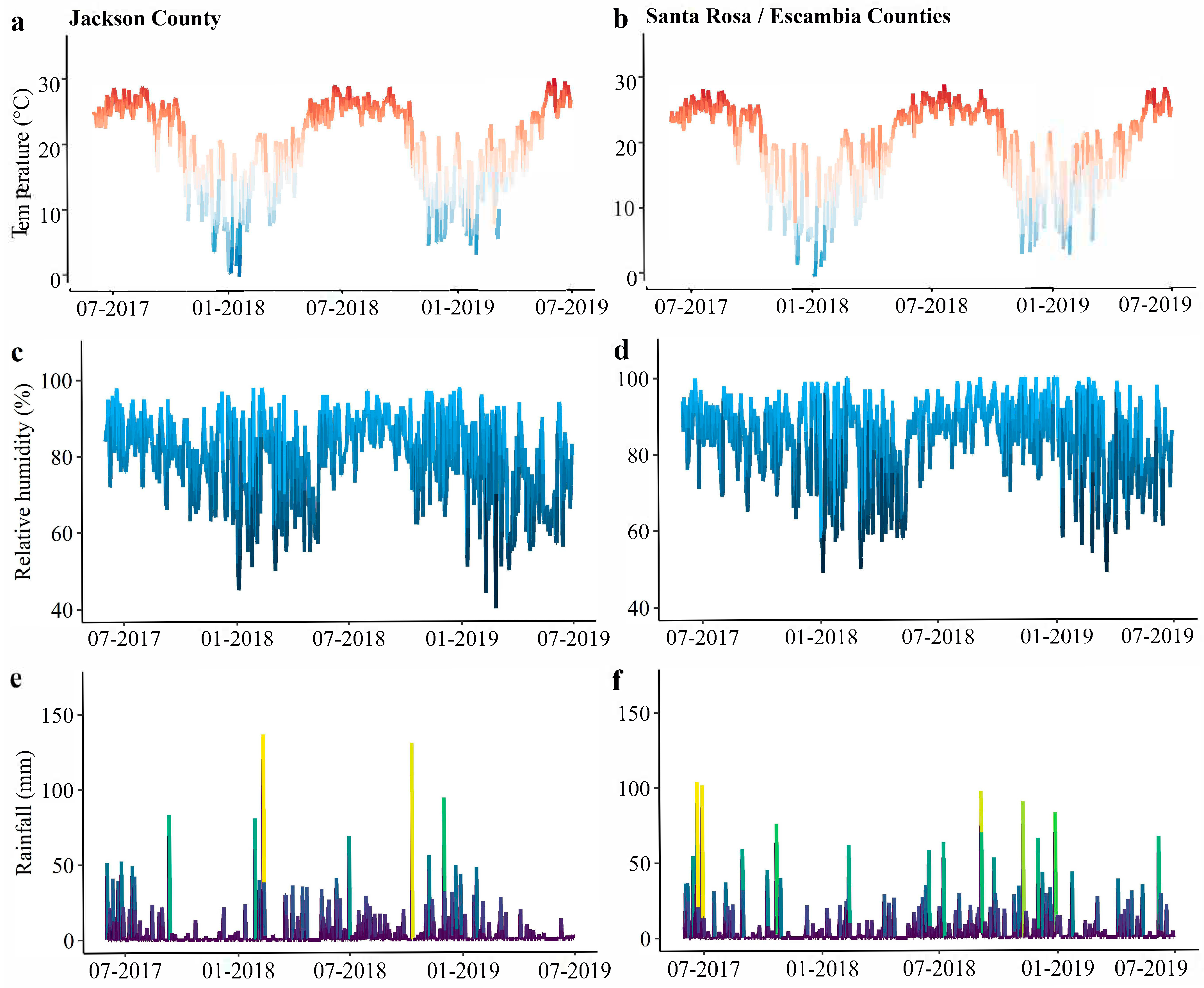
2.3. Weather Data
2.4. Statistical Analysis
3. Results
3.1. Month Abundance
3.2. Carbon Isotopic Signatures of H. zea Moths
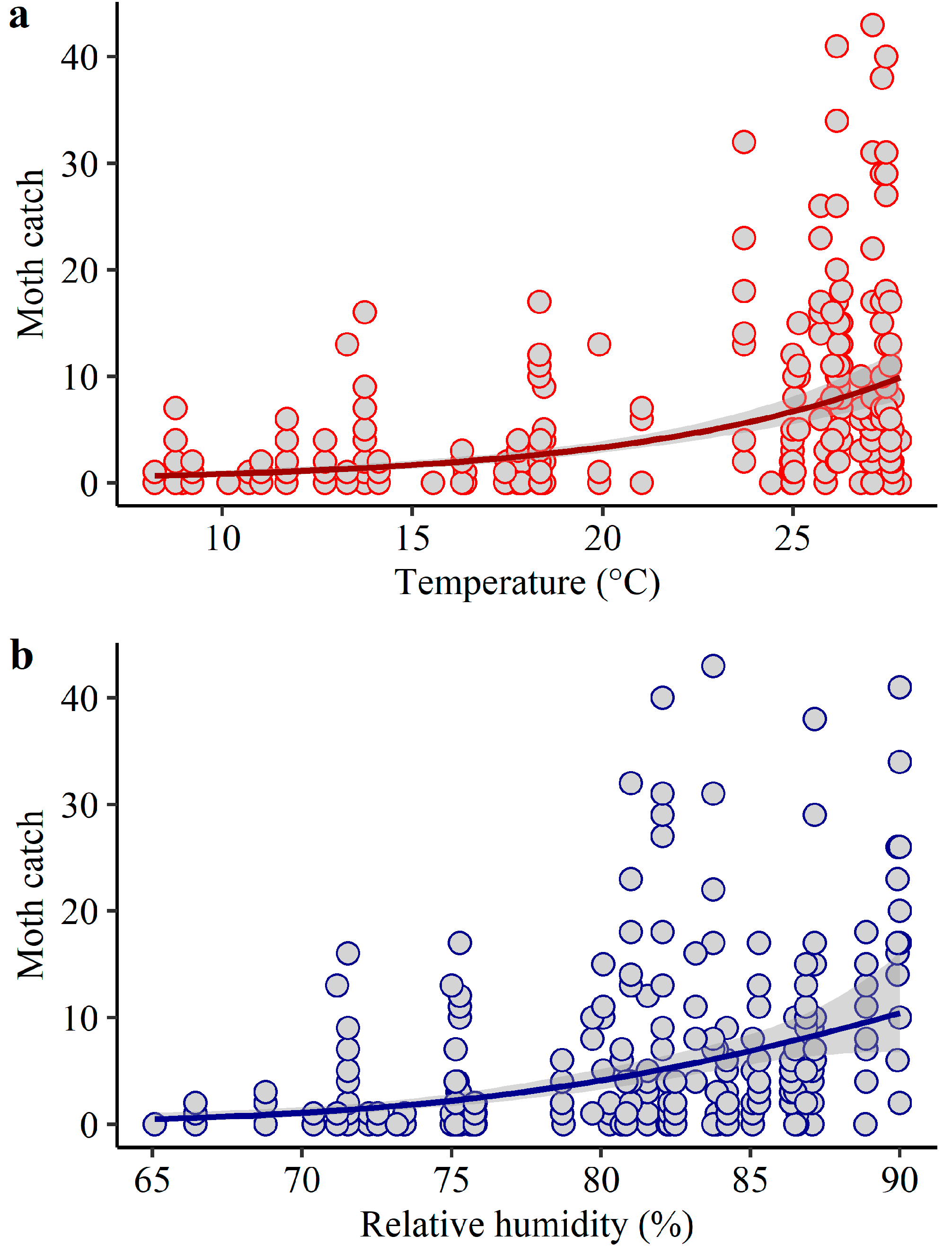
3.3. Association of Moth Catches and Weather in the Florida Panhandle from 2017 to 2019
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neunzig, H.H. Wild Host Plants of the Corn Earworm and the Tobacco Budworm in Eastern North Carolina. J. Econ. Entomol. 1963, 56, 135–139. [Google Scholar] [CrossRef]
- Neunzig, H.H. Biology of the Tobacco Budworm and the Corn Earworm in North Carolina. North Carol. Agric. Exp. Stn. 1969, 196, 63. [Google Scholar]
- Harding, J.A. Heliothis spp.: Seasonal Occurrence, Hosts and Host Importance in the Lower Rio Grande Valley. Environ. Entomol. 1976, 5, 666–668. [Google Scholar] [CrossRef]
- Stadelbacher, E.A. Role of Early-Season Wild and Naturalized Host Plants in the Buildup of the F1 Generation of Heliothis zea and H. virescens in the Delta of Mississippi. Environ. Entomol. 1981, 10, 766–770. [Google Scholar] [CrossRef]
- Capinera, J. Corn Earworm, Helicoverpa (=Heliothis) zea (Boddie) (Lepidoptera: Noctuidae). EDIS 2002, 2002. [Google Scholar] [CrossRef]
- Hardwick, D.F. The Corn Earworm Complex. Mem. Entomol. Soc. Can. 1965, 97, 5–247. [Google Scholar] [CrossRef]
- Snow, J.W.; Copeland, W.W. Distribution and Abundance of the Corn Earworm in the United States. Dep. Agric. Plant Pest Control Div. Coop. Econ. Insect Rep. 1971, 21, 71–76. [Google Scholar]
- Dorman, S.J.; Hopperstad, K.A.; Reich, B.J.; Kennedy, G.; Huseth, A.S. Soybeans as a Non-Bt Refuge for Helicoverpa zea in Maize-Cotton Agroecosystems. Agric. Ecosyst. Environ. 2021, 322, 107642. [Google Scholar] [CrossRef]
- Lawton, D.; Huseth, A.S.; Kennedy, G.G.; Morey, A.C.; Hutchison, W.D.; Reisig, D.D.; Dorman, S.J.; Dillard, D.; Venette, R.C.; Groves, R.L.; et al. Pest Population Dynamics Are Related to a Continental Overwintering Gradient. Proc. Natl. Acad. Sci. USA 2022, 119, e2203230119. [Google Scholar] [CrossRef]
- Quaintance, A.L.; Brues, C.T. The Cotton Bollworm. USDA Burn. Bull. 1905, 50, 98–99. [Google Scholar]
- Kennedy, G.G.; Storer, N.P. Life Systems of Polyphagous Arthropod Pests in Temporally Unstable Cropping Systems. Annu. Rev. Entomol. 2000, 45, 467–493. [Google Scholar] [CrossRef] [PubMed]
- Blanco, C.A.; Terán-Vargas, A.P.; López, J.D.; Kauffman, J.V.; Wei, X. Densities of Heliothis virescens and Helicoverpa zea (Lepidoptera: Noctuidae) in Three Plant Hosts. Fla. Entomol. 2007, 90, 742–750. [Google Scholar] [CrossRef]
- Gould, F.; Blair, N.; Reid, M.; Rennie, T.L.; Lopez, J.; Micinski, S. Bacillus thuringiensis-Toxin Resistance Management: Stable Isotope Assessment of Alternate Host Use by Helicoverpa zea. Proc. Natl. Acad. Sci. USA 2002, 99, 16581–16586. [Google Scholar] [CrossRef] [PubMed]
- Head, G.; Jackson, R.E.; Adamczyk, J.; Bradley, J.R.; Van Duyn, J.; Gore, J.; Hardee, D.D.; Leonard, B.R.; Luttrell, R.; Ruberson, J.; et al. Spatial and Temporal Variability in Host Use by Helicoverpa zea as Measured by Analyses of Stable Carbon Isotope Ratios and Gossypol Residues: Host Use by Cotton Bollworms. J. Appl. Ecol. 2010, 47, 583–592. [Google Scholar] [CrossRef]
- Seymour, M.; Perera, O.P.; Fescemyer, H.W.; Jackson, R.E.; Fleischer, S.J.; Abel, C.A. Peripheral Genetic Structure of Helicoverpa zea Indicates Asymmetrical Panmixia. Ecol. Evol. 2016, 6, 3198–3207. [Google Scholar] [CrossRef] [PubMed]
- USEPA White Paper on Resistance in Lepidopteran Pests of Bacillus thuringiensis (Bt) Plant Incorporated Protectants in the United States. 2018. Available online: https://www.epa.gov/sites/production/files/2018-07/documents/position_paper_07132018.pdf (accessed on 4 January 2023).
- Reay-Jones, F.P.F. Pest Status and Management of Corn Earworm (Lepidoptera: Noctuidae) in Field Corn in the United States. J. Integr. Pest Manag. 2019, 10, 19. [Google Scholar] [CrossRef]
- Yamamoto, T. Engineering of Bacillus thuringiensis Insecticidal Proteins. J. Pestic. Sci. 2022, 47, 47–58. [Google Scholar] [CrossRef]
- Carrière, Y.; Crickmore, N.; Tabashnik, B.E. Optimizing Pyramided Transgenic Bt Crops for Sustainable Pest Management. Nat. Biotechnol. 2015, 33, 161–168. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Reisig, D.D. Management of Insect Pests with Bt Crops in the United States. Annu. Rev. Entomol. 2023, 68, 31–49. [Google Scholar] [CrossRef]
- Roach, S.H.; Adkisson, P.L. Termination of Pupal Diapause in the Bollworm. J. Econ. Entomol. 1971, 64, 1057–1060. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Wolf, W.W.; Lingren, P.D.; Raulston, J.R.; Lopez, J.D., Jr.; Matis, J.H.; Eyster, R.S.; Esquivel, J.F.; Schleider, P.G. Early-Season Migratory Flights of Corn Earworm (Lepidoptera: Noctuidae). Environ. Entomol. 1997, 26, 12–20. [Google Scholar] [CrossRef]
- Wolf, W.W.; Westbrook, J.K.; Raulston, J.; Pair, S.D.; Hobbs, S.E. Recent Airborne Radar Observations of Migrant Pests in the United States. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1997, 328, 619–630. [Google Scholar] [CrossRef]
- Westbrook, J.K.; López, J.D. Long-Distance Migration in Helicoverpa zea: What We Know and Need to Know. Southwest. Entomol. 2010, 35, 355–360. [Google Scholar] [CrossRef]
- Morey, A.C.; Hutchison, W.D.; Venette, R.C.; Burkness, E.C. Cold Hardiness of Helicoverpa zea (Lepidoptera: Noctuidae) Pupae. Environ. Entomol. 2012, 41, 172–179. [Google Scholar] [CrossRef]
- FDACS Florida Department of Agriculture and Consumer Services. National Agricultural Statistics Service-USDA. 2018. Available online: https://quickstats.nass.usda.gov/ (accessed on 4 January 2023).
- USEPA The Environmental Protection Agency. Meeting Minutes of the FIFRA Scientific Advisory Panel Meeting Held 13–15 June 2006 on the Analysis of a Natural Refuge of Non-Cotton Hosts for Monsanto’s Bollgard II Cotton. 2006. Available online: https://www.regulations.gov/document/EPA-HQ-OPP-2006-0217-0042 (accessed on 4 January 2022).
- Kaur, G.; Guo, J.; Brown, S.; Head, G.P.; Price, P.A.; Paula-Moraes, S.; Ni, X.; Dimase, M.; Huang, F. Field-Evolved Resistance of Helicoverpa zea (Boddie) to Transgenic Maize Expressing Pyramided Cry1A.105/Cry2Ab2 Proteins in Northeast Louisiana, the United States. J. Invertebr. Pathol. 2019, 163, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Lin, S.; Dimase, M.; Niu, Y.; Brown, S.; Head, G.P.; Price, P.A.; Reay-Jones, F.P.F.; Cook, D.; Reisig, D.; et al. Extended Investigation of Field-Evolved Resistance of the Corn Earworm Helicoverpa zea (Lepidoptera: Noctuidae) to Bacillus thuringiensis Cry1A.105 and Cry2Ab2 Proteins in the southeastern United States. J. Invertebr. Pathol. 2021, 183, 107560. [Google Scholar] [CrossRef]
- Specht, A.; Sosa-Gómez, D.R.; de Paula-Moraes, S.V.; Yano, S.A.C. Morphological and Molecular Identification of Helicoverpa armigera (Lepidoptera: Noctuidae) and Expansion of Its Occurrence Record in Brazil. Pesqui. Agropecuária Bras. 2013, 48, 689–692. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Version 4.0.4. Available online: www.R-project.org (accessed on 4 January 2023).
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. GlmmTMB Balances Speed and Flexibility Among Packages for Zero-Inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378. [Google Scholar] [CrossRef]
- Lenth, R.; Buerkner, P.; Herve, M.; Love, J.; Riebl, H.; Singmann, H. Emmeans: Estimated Marginal Means, Aka Least-Squares Means, v1. 5.1; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A Review of Methods to Deal with It and a Simulation Study Evaluating Their Performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Lüdecke, D.; Ben-Shachar, M.; Patil, I.; Waggoner, P.; Makowski, D. Performance: An R Package for Assessment, Comparison and Testing of Statistical Models. J. Open Source Softw. 2021, 6, 3139. [Google Scholar] [CrossRef]
- Rabelo, M.M.; Paula-Moraes, S.V.; Pereira, E.J.G.; Siegfried, B.D. Demographic Performance of Helicoverpa zea Populations on Dual and Triple-Gene Bt Cotton. Toxins 2020, 12, 551. [Google Scholar] [CrossRef] [PubMed]
- Burkness, E.C.; Dively, G.; Patton, T.; Morey, A.C.; Hutchison, W.D. Novel Vip3A Bacillus thuringiensis (Bt) Maize Approaches High-Dose Efficacy against Helicoverpa zea (Lepidoptera: Noctuidae) under Field Conditions: Implications for Resistance Management. GM Crops 2010, 1, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Caprio, M.A. Source-Sink Dynamics Between Transgenic and Non-Transgenic Habitats and Their Role in the Evolution of Resistance. J. Econ. Entomol. 2001, 94, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Caprio, M.A.; Faver, M.K.; Hankins, G. Evaluating the Impacts of Refuge Width on Source-Sink Dynamics between Transgenic and Non-Transgenic Cotton. J. Insect Sci. 2004, 4, 3. [Google Scholar] [CrossRef]
- Marra, M.C.; Pardey, P.G.; Alston, J.M. The Payoffs to Agricultural Biotechnology: An Assessment of the Evidence. 2002. Available online: https://ageconsearch.umn.edu/record/16093/ (accessed on 2 January 2023).
- Naranjo, S.E. Long-Term Assessment of the Effects of Transgenic Bt Cotton on the Abundance of Nontarget Arthropod Natural Enemies. Environ. Entomol. 2005, 34, 1193–1210. [Google Scholar] [CrossRef]
- Torres, J.B.; Ruberson, J.R. Interactions of Bacillus thuringiensis Cry1Ac Toxin in Genetically Engineered Cotton with Predatory Heteropterans. Transgenic Res. 2008, 17, 345–354. [Google Scholar] [CrossRef]
- Hutchison, W.D.; Burkness, E.C.; Mitchell, P.D.; Moon, R.D.; Leslie, T.W.; Fleischer, S.J.; Abrahamson, M.; Hamilton, K.L.; Steffey, K.L.; Gray, M.E.; et al. Areawide Suppression of European Corn Borer with Bt Maize Reaps Savings to Non-Bt Maize Growers. Science 2010, 330, 222–225. [Google Scholar] [CrossRef]
- Adamczyk, J.J., Jr.; Hubbard, D. Changes in Populations of Heliothis virescens (F.) (Lepidoptera: Noctuidae) and Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae) in the Mississippi Delta from 1986 to 2005 as Indicated by Adult Male Pheromone Traps. J. Cotton Sci. 2006, 10, 842–847. [Google Scholar]
- Arends, B.R.; Reisig, D.D.; Gundry, S.; Greene, J.K.; Kennedy, G.G.; Reay-Jones, F.P.F.; Huseth, A.S. Helicoverpa zea (Lepidoptera: Noctuidae) Feeding Incidence and Survival on Bt Maize in Relation to Maize in the Landscape. Pest Manag. Sci. 2022, 78, 2309–2315. [Google Scholar] [CrossRef]
- Felland, C.M.; Pitre, H.N. Survivorship and Development of Noctuid Larvae (Lepidoptera: Noctuidae) on Irrigated and Dryland Soybean. Environ. Entomol. 1991, 20, 1315–1321. [Google Scholar] [CrossRef]
- Young, J.H.; Price, R.G. Effect of Irrigation and Submersion in Water on Pupal Survival of the Bollworm. J. Econ. Entomol. 1968, 61, 959–961. [Google Scholar] [CrossRef]
- Burkness, E.C.; Cira, T.M.; Moser, S.E.; Hutchison, W.D. Bt Maize Seed Mixtures for Helicoverpa zea (Lepidoptera: Noctuidae): Larval Movement, Development, and Survival on Non-Transgenic Maize. J. Econ. Entomol. 2015, 108, 2761–2769. [Google Scholar] [CrossRef]
- Dimase, M.; Oyediran, I.; Brown, S.; Walker, W.; Guo, J.; Yu, W.; Zhang, Y.; Chen, J.; Wen, Z.; Huang, F. Larval Movement and Survival of Helicoverpa zea (Boddie) in Seed Blends of Non-Bt and Bt Maize Containing Agrisure Viptera® Trait: Implications for Resistance Management. Crop Prot. 2020, 138, 105339. [Google Scholar] [CrossRef]
- Wright, D.; Small, I.; Mackowiak, C.; Grabau, Z.; Devkota, P.; Paula-Moraes, S. Field Corn Production Guide: SS-AGR-85/AG202, Rev. 8/2022. EDIS 2022, 2022, 13. [Google Scholar] [CrossRef]
- Arends, B.; Reisig, D.D.; Gundry, S.; Huseth, A.S.; Reay-Jones, F.P.F.; Greene, J.K.; Kennedy, G.G. Effectiveness of the Natural Resistance Management Refuge for Bt-Cotton Is Dominated by Local Abundance of Soybean and Maize. Sci. Rep. 2021, 11, 17601. [Google Scholar] [CrossRef]
- Reisig, D.D.; Huseth, A.S.; Bacheler, J.S.; Aghaee, M.-A.; Braswell, L.; Burrack, H.J.; Flanders, K.; Greene, J.K.; Herbert, D.A.; Jacobson, A.; et al. Long-Term Empirical and Observational Evidence of Practical Helicoverpa zea Resistance to Cotton With Pyramided Bt Toxins. J. Econ. Entomol. 2018, 111, 1824–1833. [Google Scholar] [CrossRef]
- FDACS Florida Department of Agriculture and Consumer Services. National Agricultural Statistics Service-USDA. 2020. Available online: https://quickstats.nass.usda.gov/ (accessed on 4 January 2023).
- Kanissery, R.; Frey, C.; Dufault, N.S.; Desaeger, J.; Beuzelin, J. Sweet Corn Production: VPH Ch. 17, CV135/HS737, Rev. 5/2022. EDIS 2022. [Google Scholar] [CrossRef]
- Schwan Resende, E.; Beuzelin, J.M.; Dunkley, V.E.; Paula-Moraes, S.V.; Seal, D.R.; Nuessly, G.S. Pyrethroid Susceptibility in Field Populations of Picture-Winged Flies (Diptera: Ulidiidae) Infesting Fresh Market Sweet Corn in Florida. J. Econ. Entomol. 2022, 115, 1685–1692. [Google Scholar] [CrossRef]
- Buntin, G.D.; Lee, R.D.; Wilson, D.M.; McPherson, R.M. Evaluation of Yieldgard Transgenic Resistance for Control of Fall Armyworm and Corn Earworm (Lepidoptera: Noctuidae) on Corn. Fla. Entomol. 2001, 84, 37–42. [Google Scholar] [CrossRef]
- Olmstead, D.L.; Nault, B.A.; Shelton, A.M. Biology, Ecology, and Evolving Management of Helicoverpa zea (Lepidoptera: Noctuidae) in Sweet Corn in the United States. J. Econ. Entomol. 2016, 109, 1667–1676. [Google Scholar] [CrossRef]
- Lingren, P.D.; Westbrook, J.K.; Bryant, V.M.; Raulston, J.R.; Esquivel, J.F.; Jones, G.D. Origin of Corn Earworm (Lepidoptera: Noctuidae) Migrants as Determined by Citrus Pollen Markers and Synoptic Weather Systems. Environ. Entomol. 1994, 23, 562–570. [Google Scholar] [CrossRef]
- Yela, J.L.; Holyoak, M. Effects of Moonlight and Meteorological Factors on Light and Bait Trap Catches of Noctuid Moths (Lepidoptera: Noctuidae). Environ. Entomol. 1997, 26, 1283–1290. [Google Scholar] [CrossRef]
- Slosser, J.E.; Witz, J.A.; Puterka, G.J.; Price, J.R.; Hartstack, A.W. Seasonal Changes in Bollworm (Lepidoptera: Noctuidae) Moth Catches in Pheromone Traps in a Large Area. Environ. Entomol. 1987, 16, 1296–1301. [Google Scholar] [CrossRef]
- Parajulee, M.N.; Rummel, D.R.; Arnold, M.D.; Carroll, S.C. Long-Term Seasonal Abundance Patterns of Helicoverpa zea and Heliothis virescens (Lepidoptera: Noctuidae) in the Texas High Plains. J. Econ. Entomol. 2004, 97, 668–677. [Google Scholar] [CrossRef]
- Royer, L.; McNeil, J.N. Effect of Relative Humidity Conditions on Responsiveness of European Corn Borer (Ostrinia nubilalis) Males to Female Sex Pheromone in a Wind Tunnel. J. Chem. Ecol. 1993, 19, 61–69. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Variable | χ2 | DF | p-Value 1 |
|---|---|---|---|---|
| Jackson | Crop | 0.21 | 1 | 0.65 |
| Irrigation | 0.15 | 1 | 0.90 | |
| Crop × Irrigation | 2.01 | 1 | 0.46 | |
| Santa Rosa/Escambia | Crop | 0.54 | 1 | 0.46 |
| Irrigation | 4.45 | 1 | 0.03 | |
| Crop × Irrigation | 0.13 | 1 | 0.72 |
| Region | Parameter | Estimate | SD | z Value | Pr (>|z|) |
|---|---|---|---|---|---|
| Jackson | Intercept | −4.10 | 1.19 | −3.46 | 0.0005 |
| Temperature | 0.11 | 0.02 | 6.68 | <0.0001 | |
| Relative humidity | 0.04 | 0.02 | 2.19 | 0.03 | |
| Rainfall | −0.0002 | 0.00 | −0.10 | 0.92 | |
| Nagelkerke’s R2 | 0.38 | ||||
| Escambia/Santa Rosa | Intercept | 4.94 | 1.35 | 3.67 | 0.0002 |
| Temperature | 0.23 | 0.02 | 14.16 | <0.0001 | |
| Relative humidity | −0.10 | 0.02 | −5.76 | <0.0001 | |
| Rainfall | 0.005 | 0.001 | 4.06 | <0.0001 | |
| Nagelkerke’s R2 | 0.59 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, I.B.d.; Paula-Moraes, S.V.; Beuzelin, J.M.; Hahn, D.A.; Perera, O.P.; Fraisse, C. Factors Affecting Population Dynamics of Helicoverpa zea (Lepidoptera: Noctuidae) in a Mixed Landscape with Bt Cotton and Peanut. Insects 2023, 14, 395. https://doi.org/10.3390/insects14040395
Santos IBd, Paula-Moraes SV, Beuzelin JM, Hahn DA, Perera OP, Fraisse C. Factors Affecting Population Dynamics of Helicoverpa zea (Lepidoptera: Noctuidae) in a Mixed Landscape with Bt Cotton and Peanut. Insects. 2023; 14(4):395. https://doi.org/10.3390/insects14040395
Chicago/Turabian StyleSantos, Izailda Barbosa dos, Silvana V. Paula-Moraes, Julien M. Beuzelin, Daniel A. Hahn, Omaththage P. Perera, and Clyde Fraisse. 2023. "Factors Affecting Population Dynamics of Helicoverpa zea (Lepidoptera: Noctuidae) in a Mixed Landscape with Bt Cotton and Peanut" Insects 14, no. 4: 395. https://doi.org/10.3390/insects14040395