
Potential of Hedgerows with Aromatic Plants as Reservoirs of Natural Enemies of Pests in Orange Orchards

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
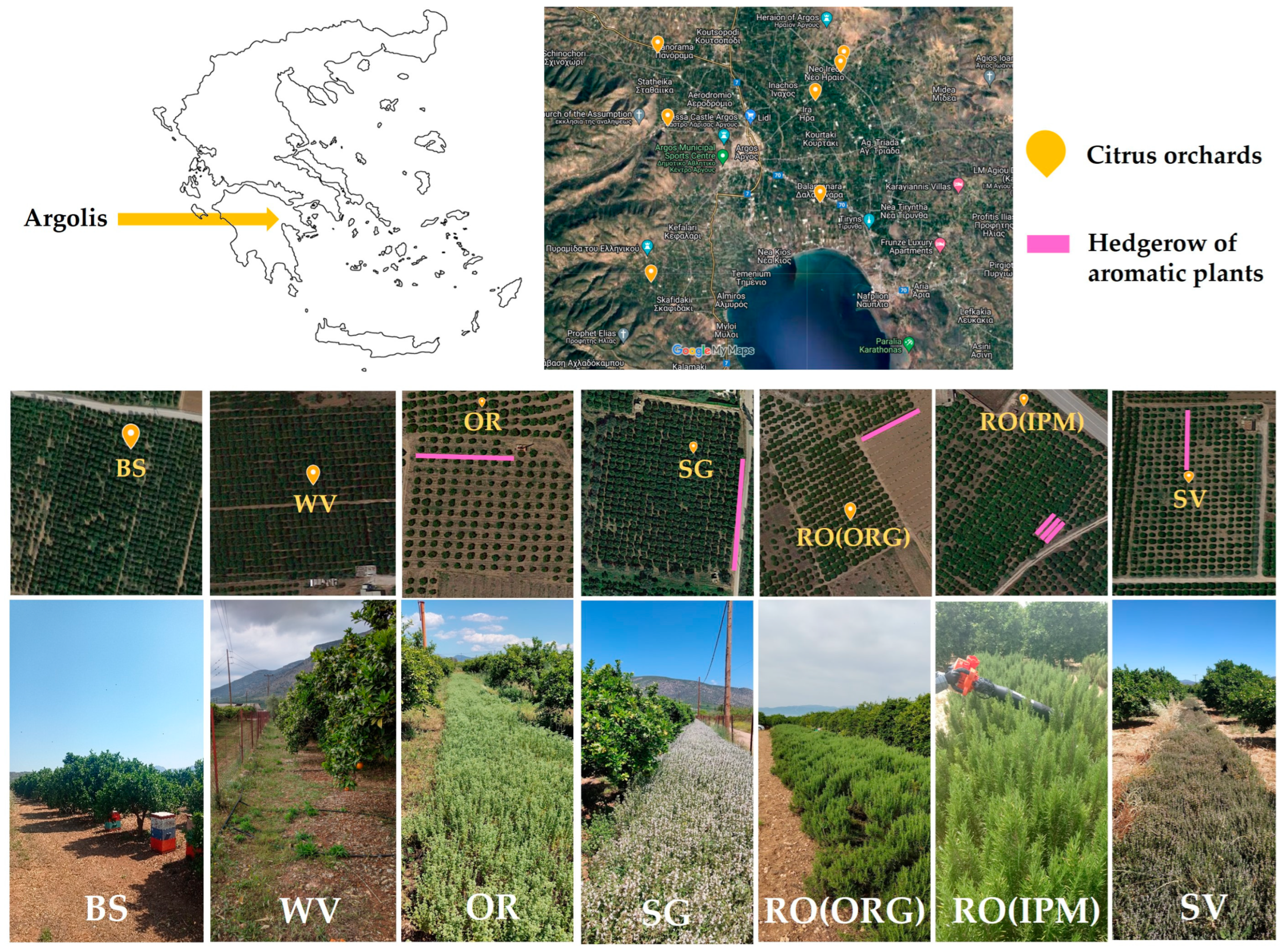
2. Materials and Methods
2.1. Experimental Fields and Establishment of Flowering Hedgerows
2.2. Flower and Plant Cover Measurements in Field Margins
2.3. Arthropod Presence Measurements
2.4. Statistical Analysis
3. Results
3.1. Baseline Year
3.2. After the Establishment of Hedgerows
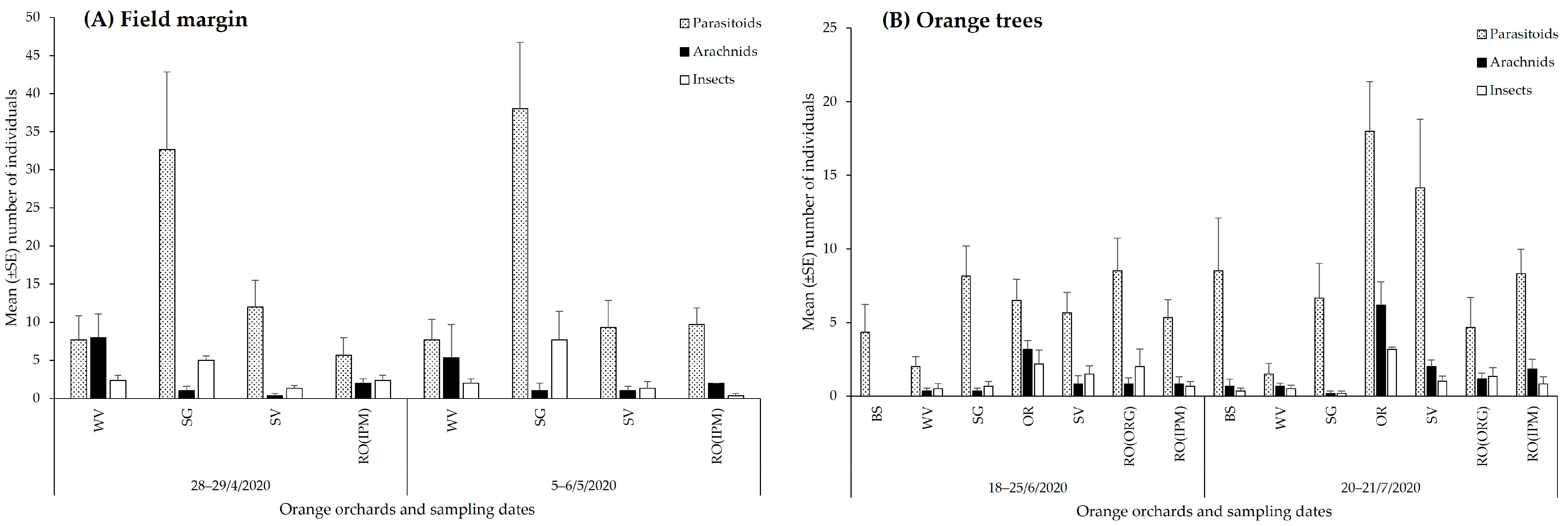
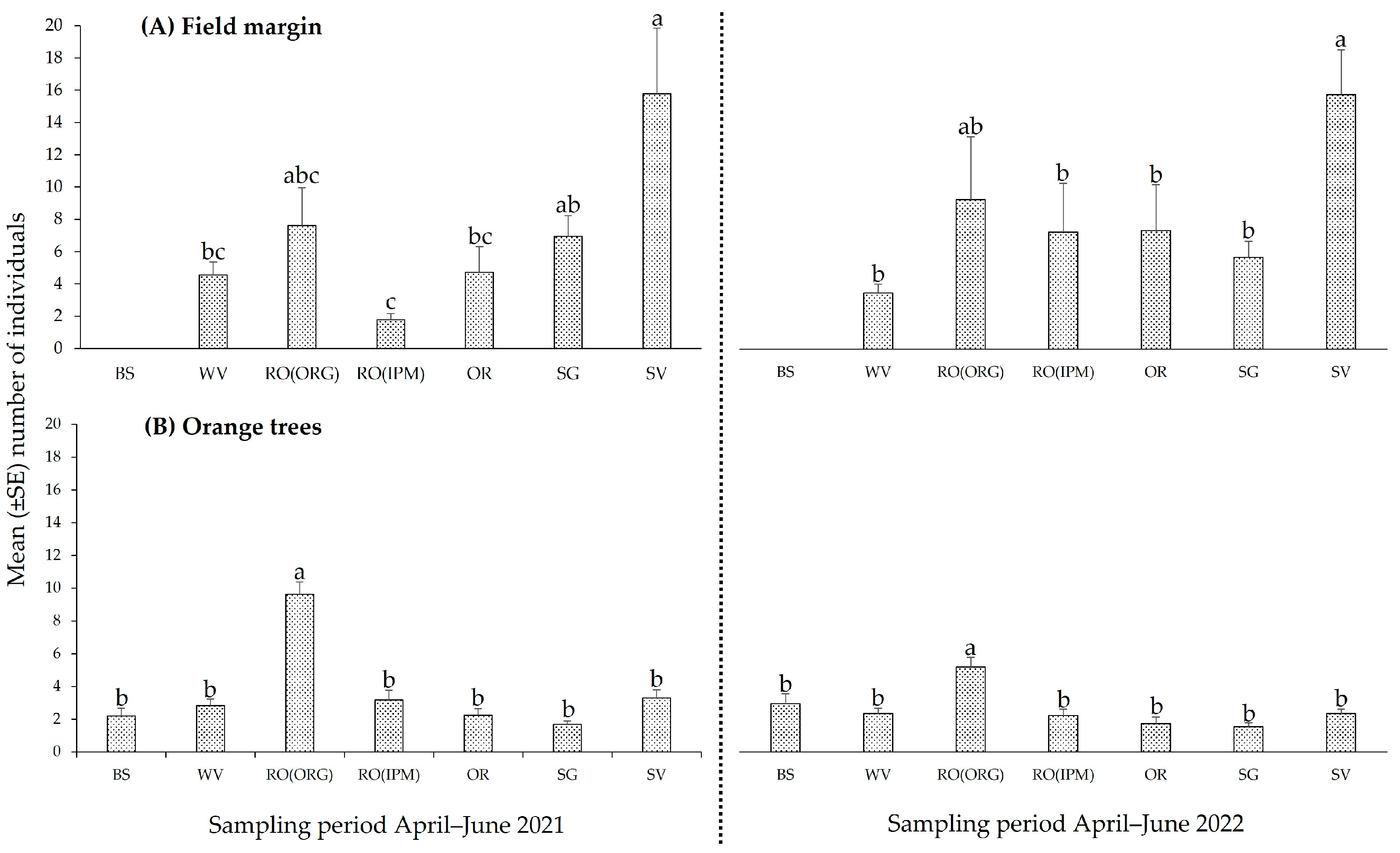
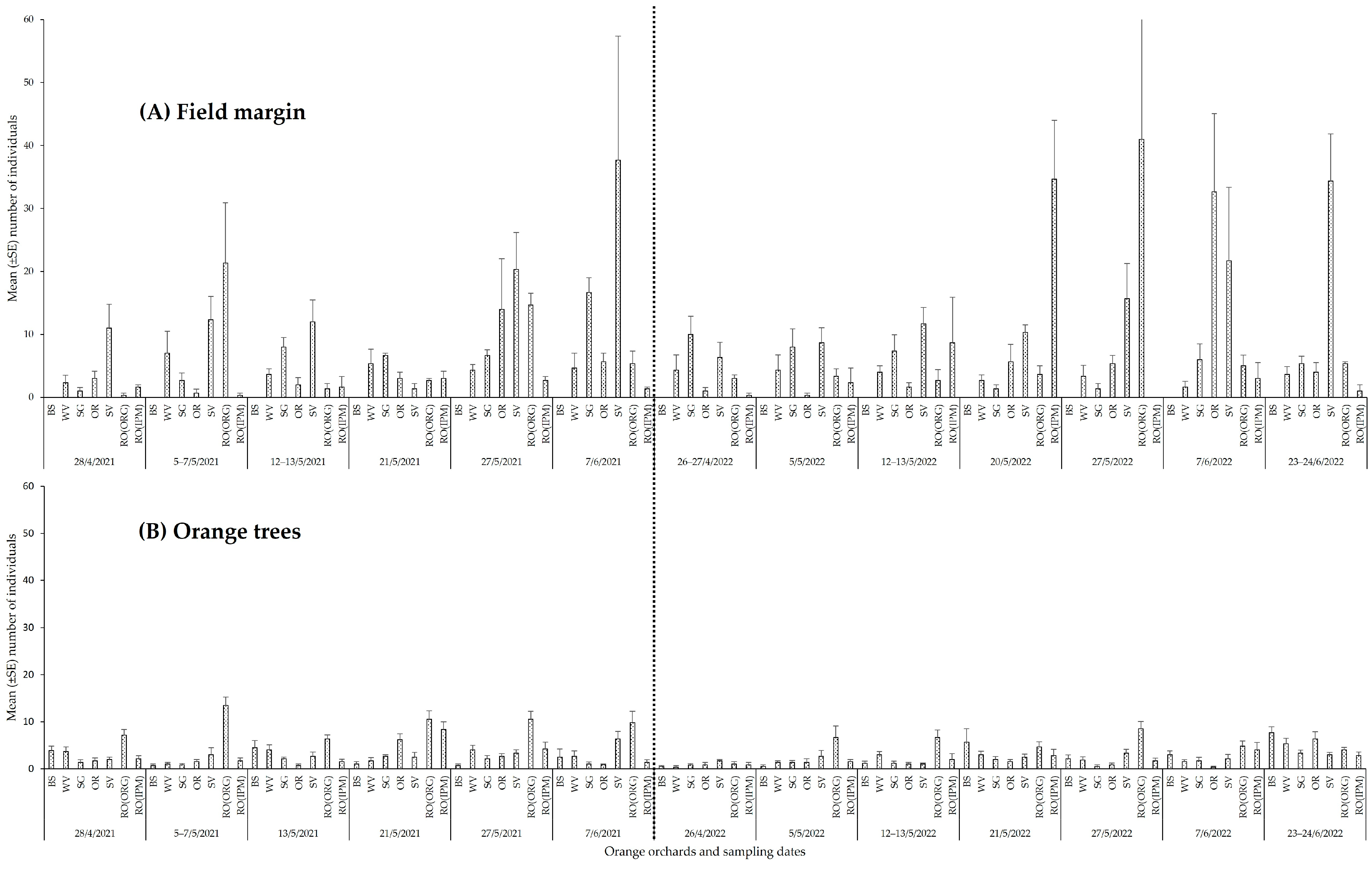
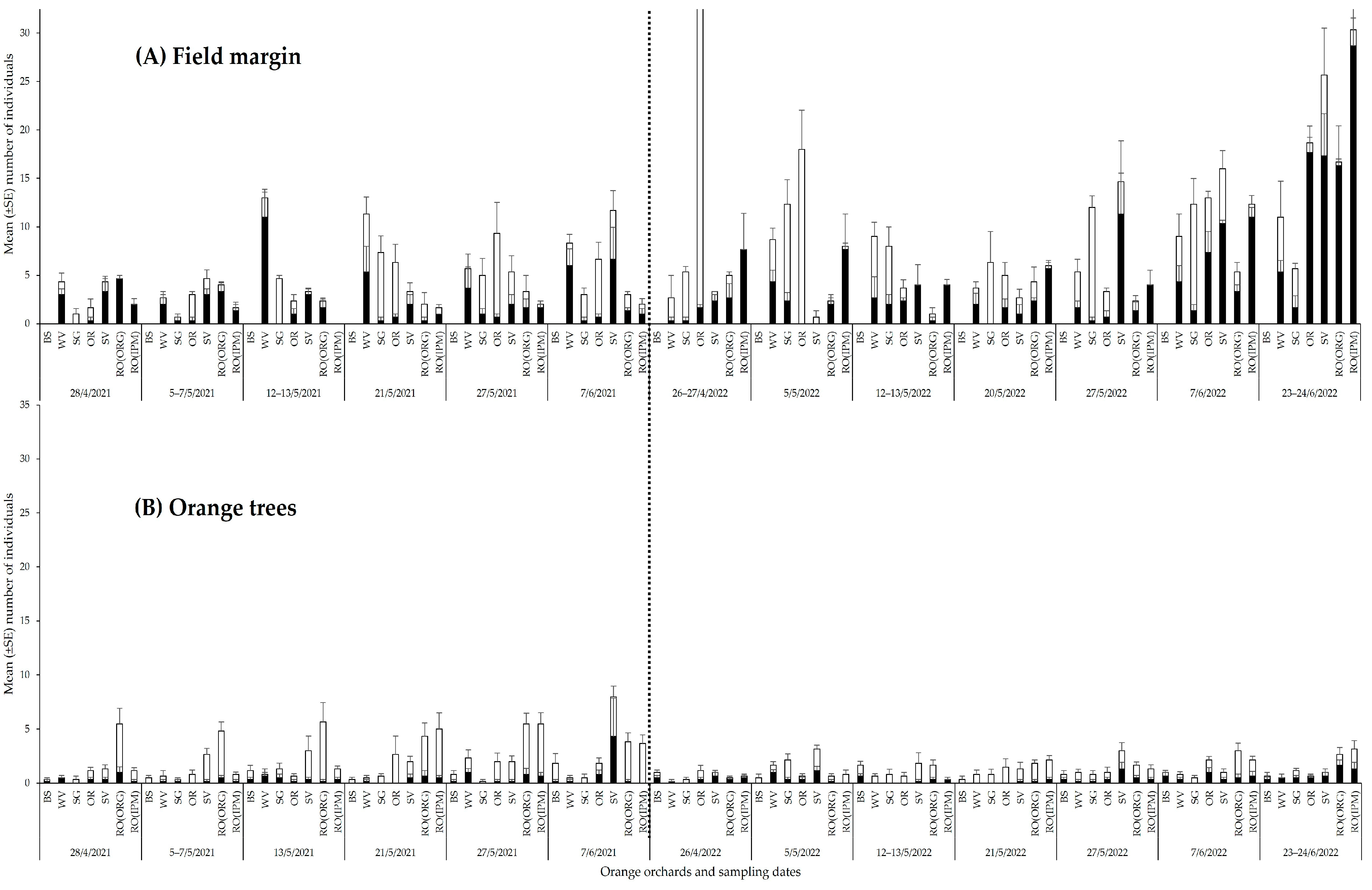
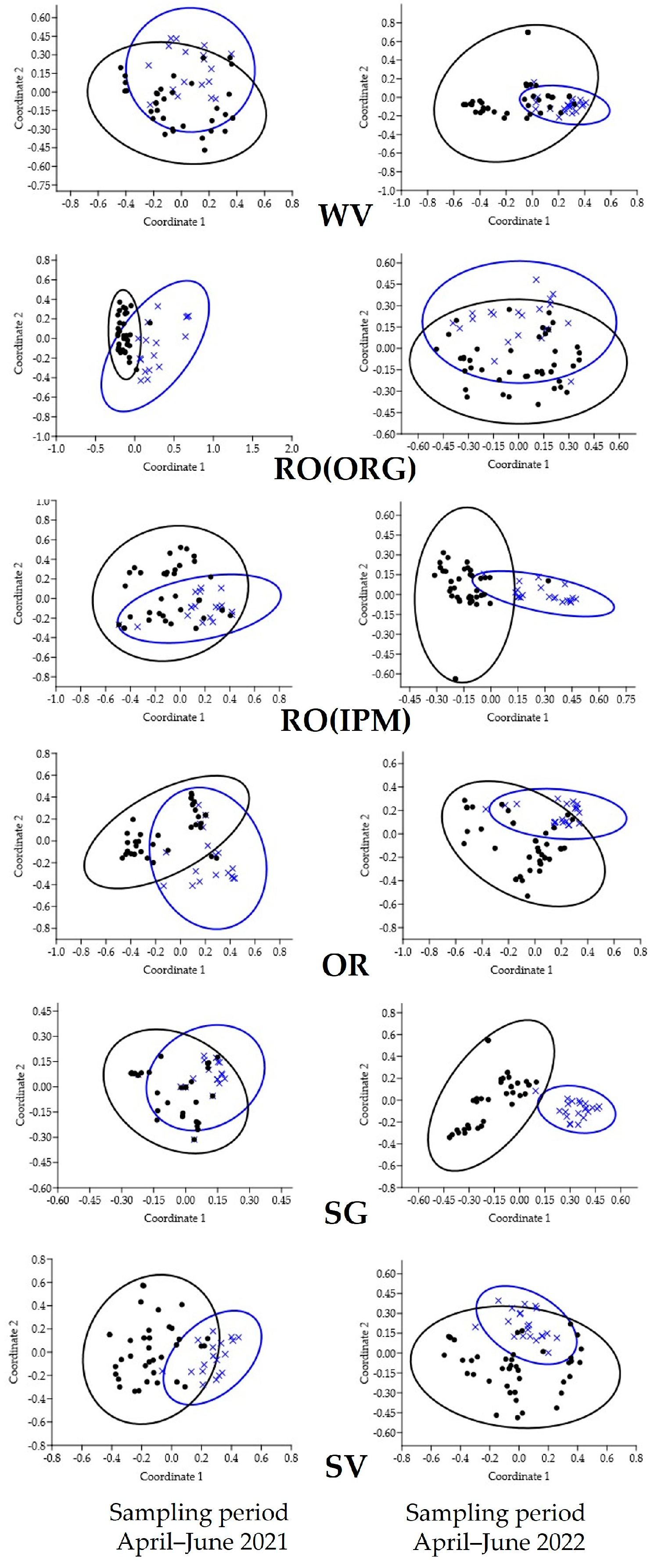
3.2.1. Spatial and Temporal Patterns of Natural Enemies
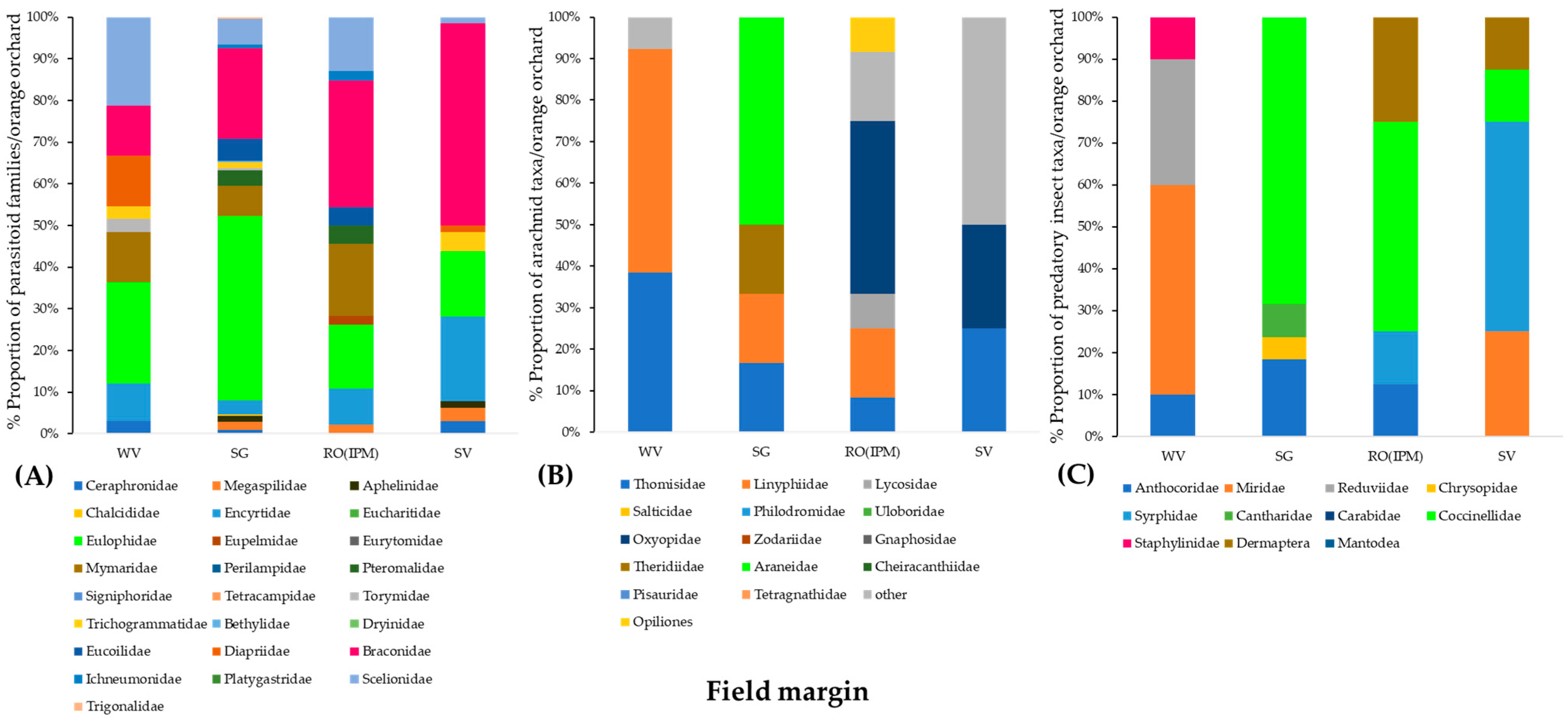
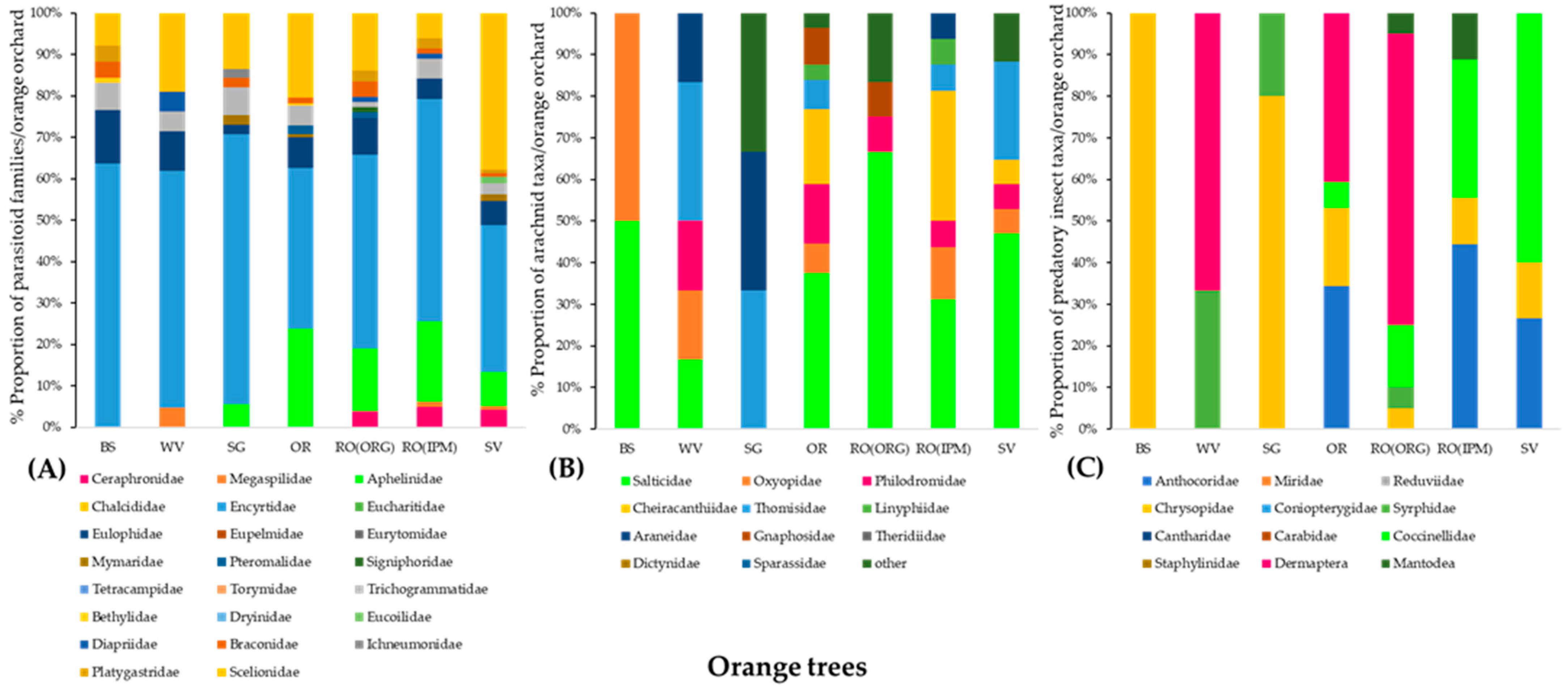
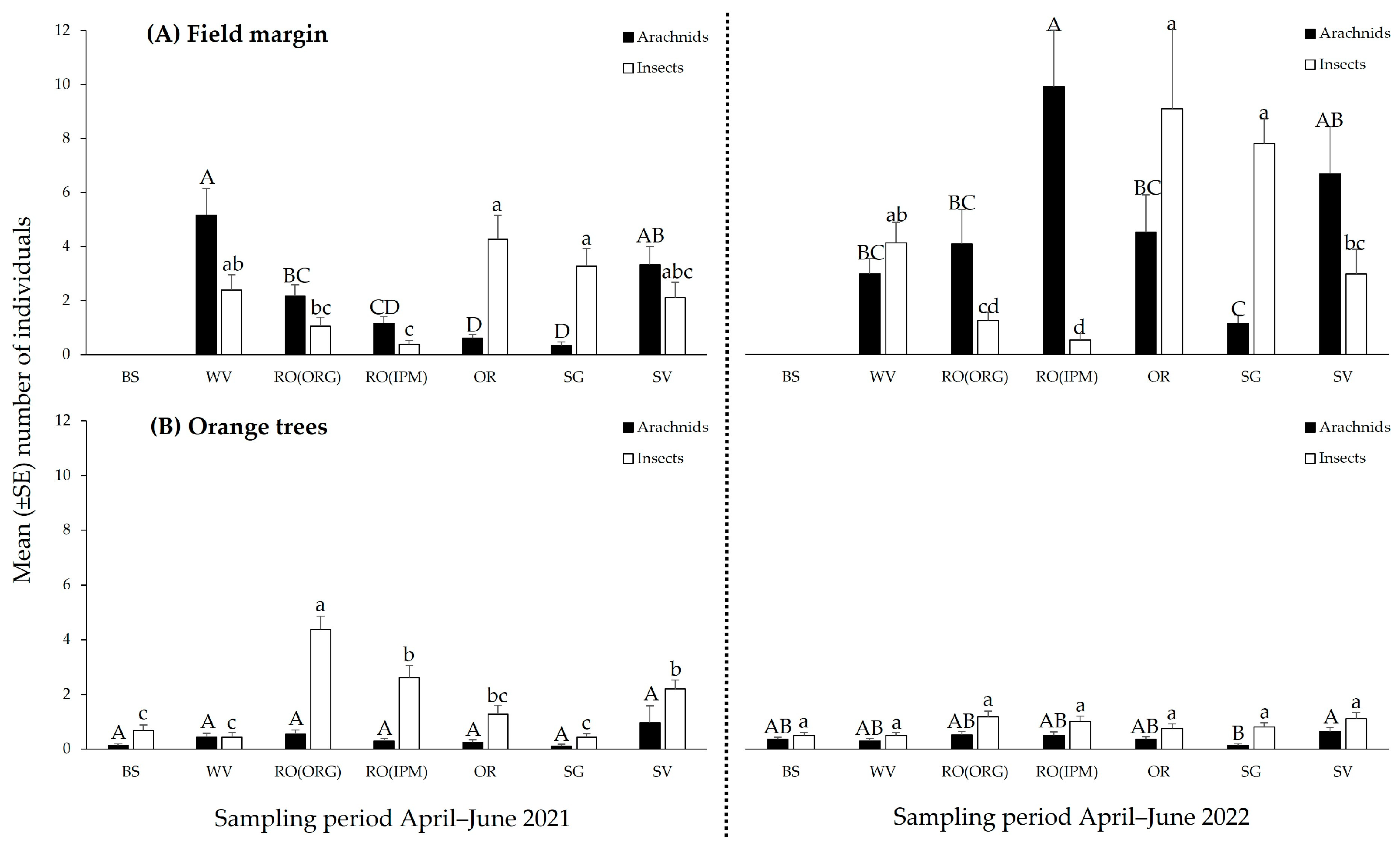
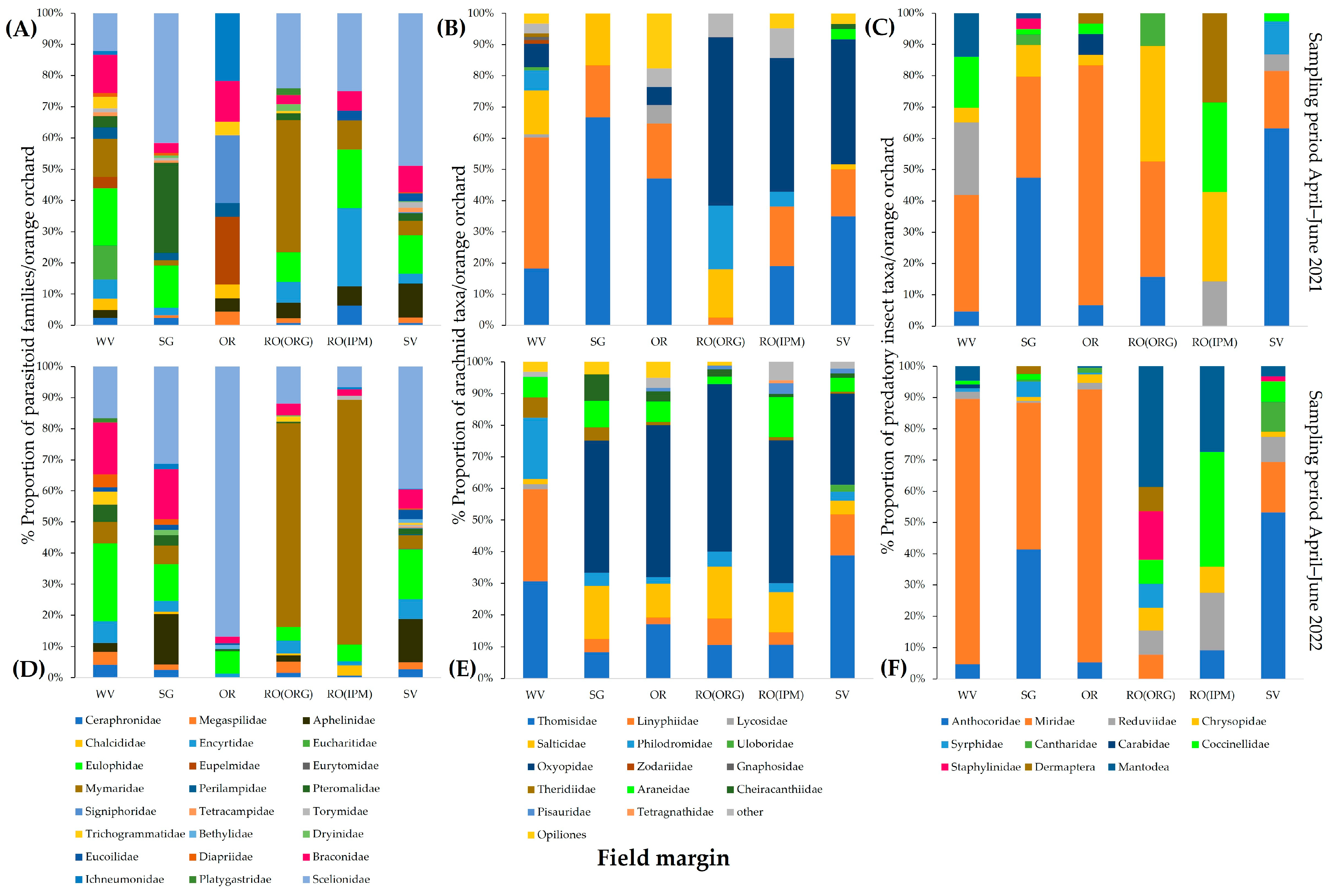
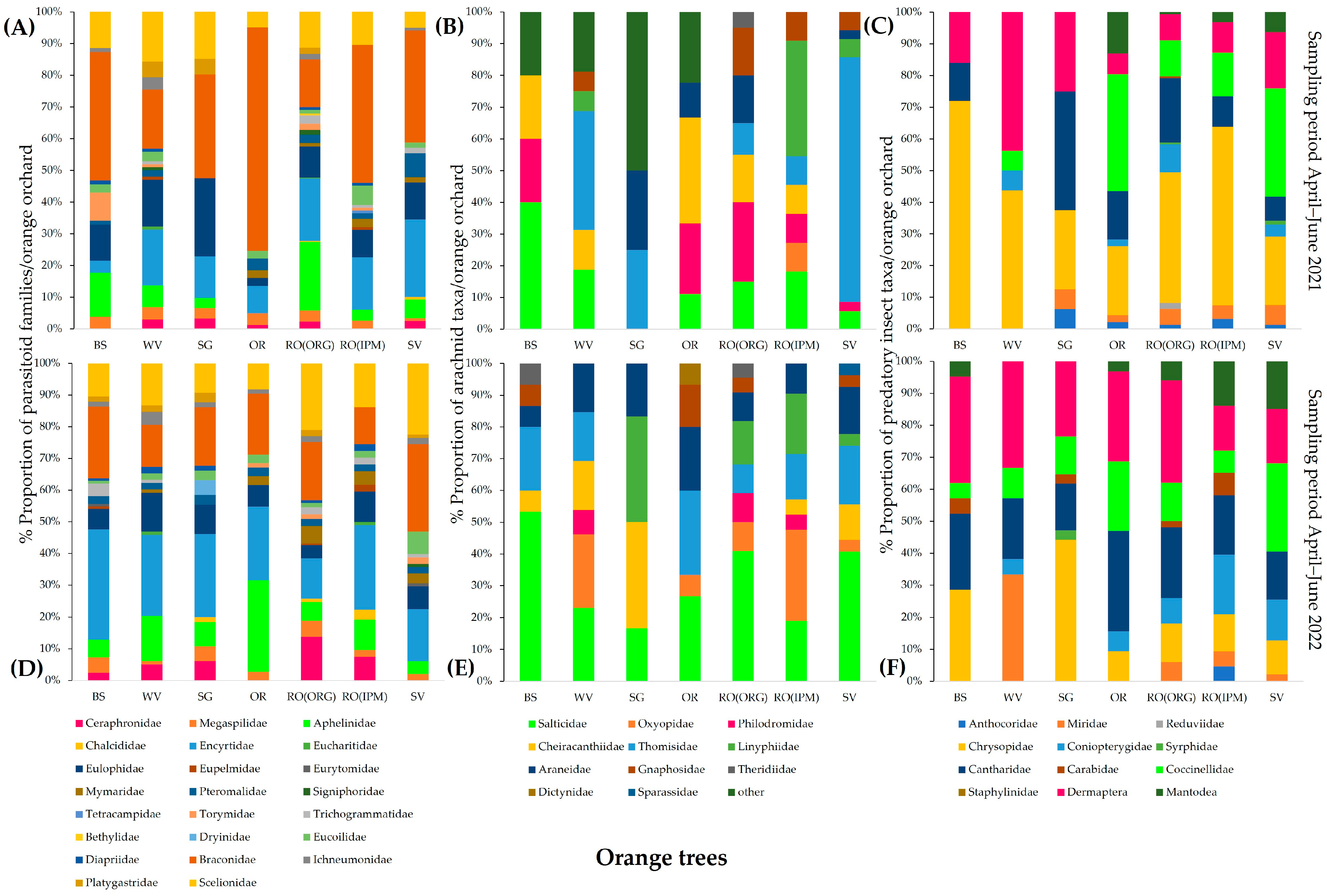
3.2.2. Community Composition of Natural Enemies
3.2.3. Phytophagous Arthropods
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Wezel, A.; Goris, M.; Bruil, J.; Felix, G.F.; Peeters, A.; Barberi, P.; Bellon, S.; Migliorini, P. Challenges and Action Points to Amplify Agroecology in Europe. Sustainability 2018, 10, 1598. [Google Scholar] [CrossRef]
- Geiger, F.; Wäckers, F.L.; Bianchi, F.J.J.A. Hibernation of Predatory Arthropods in Semi-Natural Habitats. BioControl 2009, 54, 529–535. [Google Scholar] [CrossRef]
- Meek, B.; Loxton, D.; Sparks, T.; Pywell, R.; Pickett, H.; Nowakowski, M. The Effect of Arable Field Margin Composition on Invertebrate Biodiversity. Biol. Conserv. 2002, 106, 259–271. [Google Scholar] [CrossRef]
- Lavandero, B.; Wratten, S.D.; Didham, R.K.; Gurr, G. Increasing Floral Diversity for Selective Enhancement of Biological Control Agents: A Double-Edged Sward? Basic Appl. Ecol. 2006, 7, 236–243. [Google Scholar] [CrossRef]
- Isaacs, R.; Tuell, J.; Fiedler, A.; Gardiner, M.; Landis, D. Maximizing Arthropod-Mediated Ecosystem Services in Agricultural Landscapes: The Role of Native Plants. Front. Ecol. Environ. 2009, 7, 196–203. [Google Scholar] [CrossRef]
- Altieri, M.A.; Letourneau, D.K. Vegetation Management and Biological Control in Agroecosystems. Crop Prot. 1982, 1, 405–430. [Google Scholar] [CrossRef]
- Wyss, E. The Effects of Weed Strips on Aphids and Aphidophagous Predators in an Apple Orchard. Entomol. Exp. Appl. 1995, 75, 43–49. [Google Scholar] [CrossRef]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat Management to Conserve Natural Enemies of Arthropod Pests in Agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Boller, E.F.; Häni, F.; Poehling, H.-M. Ecological Infrastructures: Ideabook on Functional Biodiversity at the Farm Level. In Ecological Infrastructures: Ideabook on Functional Biodiversity at the Farm Level; Landwirtschaftliche Beratungszentrale Lindau: Lindau, Switzerland, 2004. [Google Scholar]
- Gurr, G.M.; Wratten, S.D.; Altieri, M.A. Ecological Engineering for Pest Management: Advances in Habitat Manipulation for Arthropods; CSIRO Publishing: Collingwood, VI, Australia, 2004. [Google Scholar]
- Ricketts, T.H.; Regetz, J.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Bogdanski, A.; Gemmill-Herren, B.; Greenleaf, S.S.; Klein, A.M.; Mayfield, M.M.; et al. Landscape Effects on Crop Pollination Services: Are There General Patterns? Ecol. Lett. 2008, 11, 499–515. [Google Scholar] [CrossRef]
- Tscharntke, T.; Bommarco, R.; Clough, Y.; Crist, T.O.; Kleijn, D.; Rand, T.A.; Tylianakis, J.M.; Nouhuys, S.V.; Vidal, S. Conservation biological control and enemy diversity on a landscape scale. Biol. Control 2007, 43, 294–309. [Google Scholar] [CrossRef]
- Holland, J.M.; Bianchi, F.J.; Entling, M.H.; Moonen, A.-C.; Smith, B.M.; Jeanneret, P. Structure, Function and Management of Semi-Natural Habitats for Conservation Biological Control: A Review of European Studies. Pest Manag. Sci. 2016, 72, 1638–1651. [Google Scholar] [CrossRef]
- Holland, J.M.; Douma, J.C.; Crowley, L.; James, L.; Kor, L.; Stevenson, D.R.W.; Smith, B.M. Semi-Natural Habitats Support Biological Control, Pollination and Soil Conservation in Europe. A Review. Agron. Sustain. Dev. 2017, 37, 31. [Google Scholar] [CrossRef]
- Ehler, L. Chapter 1—Conservation Biological Control: Past, Present, and Future. In Conservation Biological Control; Barbosa, P., Ed.; Academic Press: San Diego, CA, USA, 1998; pp. 1–8. [Google Scholar]
- Pell, J.K.; Hannam, J.J.; Steinkraus, D.C. Conservation Biological Control Using Fungal Entomopathogens. BioControl 2010, 55, 187–198. [Google Scholar] [CrossRef]
- Seastedt, T.R. Biological Control: Perspectives for Maintaining Provisioning Services in the Anthropocene. In Integrated Pest Management: Pesticide Problems, Vol. 3; Pimentel, D., Peshin, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 269–280. [Google Scholar]
- Begg, G.S.; Cook, S.M.; Dye, R.; Ferrante, M.; Franck, P.; Lavigne, C.; Lövei, G.L.; Mansion-Vaquie, A.; Pell, J.K.; Petit, S.; et al. A Functional Overview of Conservation Biological Control. Crop Prot. 2017, 97, 145–158. [Google Scholar] [CrossRef]
- Barbosa, P.A. Conservation Biological Control; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Garrido, A.; Ventura, J.J. Plagas de los Cítricos. Bases Para el Manejo Integrado; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 1993. [Google Scholar]
- Lacirignola, C.; D’Onghia, A.M. The Mediterranean citriculture: Productions and perspectives. In Citrus Tristeza Virus and Toxoptera Citricidus: A Serious Threat to the Mediterranean Citrus Industry; D’Onghia, A.M., Djelouah, K., Roistacher, C.N., Eds.; CIHEAM: Bari, Italy, 2009; pp. 13–17. [Google Scholar]
- Triantafyllidis, V.; Zotos, A.; Kosma, C.; Kokkotos, E. Environmental Implications from Long-Term Citrus Cultivation and Wide Use of Cu Fungicides in Mediterranean Soils. Water Air Soil Pollut. 2020, 231, 218. [Google Scholar] [CrossRef]
- Karamaouna, F.; Mylonas, P.; Papachristos, D.; Kontodimas, D.; Michaelakis, A.; Kapaxidi, E. Main Arthropod Pests of Citrus Culture and Pest Management in Greece. In Integrated Management of Arthropod Pests and Insect Borne Diseases; Ciancio, A., Mukerji, K.G., Eds.; Integrated Management of Plant Pests and Diseases; Springer: Dordrecht, The Netherlands, 2010; pp. 29–59. [Google Scholar]
- Kapantaidaki, D.E.; Antonatos, S.; Kontodimas, D.; Milonas, P.; Papachristos, D.P. Presence of the Invasive Whitefly Aleurocanthus spiniferus (Hemiptera: Aleyrodidae) in Greece. EPPO Bull. 2019, 49, 127–131. [Google Scholar] [CrossRef]
- Jacas, J.A.; Urbaneja, A. Biological Control in Citrus in Spain: From Classical to Conservation Biological Control. In Integrated Management of Arthropod Pests and Insect Borne Diseases; Ciancio, A., Mukerji, K.G., Eds.; Integrated Management of Plant Pests and Diseases; Springer: Dordrecht, The Netherlands, 2010; pp. 61–72. [Google Scholar]
- Aucejo, S.; Foo, M.; Gimeno, E.; Gomez-Cadenas, A.; Monfort, R.; Obiol, F.; Prades, E.; Ramis, M.; Ripolles, J.L.; Tirado, V.; et al. Management of Tetranychus urticae in citrus in Spain: Acarofauna associated to weeds. IOBC-WPRS Bull. 2003, 26, 213–220. [Google Scholar]
- Aguilar-Fenollosa, E.; Ibáñez-Gual, M.V.; Pascual-Ruiz, S.; Hurtado, M.; Jacas, J.A. Effect of Ground-Cover Management on Spider Mites and Their Phytoseiid Natural Enemies in Clementine Mandarin Orchards (II): Top-down Regulation Mechanisms. Biol. Control 2011, 59, 171–179. [Google Scholar] [CrossRef]
- Aguilar-Fenollosa, E.; Ibáñez-Gual, M.V.; Pascual-Ruiz, S.; Hurtado, M.; Jacas, J.A. Effect of Ground-Cover Management on Spider Mites and Their Phytoseiid Natural Enemies in Clementine Mandarin Orchards (I): Bottom-up Regulation Mechanisms. Biol. Control 2011, 59, 158–170. [Google Scholar] [CrossRef]
- Aguilar-Fenollosa, E.; Pascual-Ruiz, S.; Hurtado, M.A.; Jacas, J.A. Efficacy and Economics of Ground Cover Management as a Conservation Biological Control Strategy against Tetranychus urticae in Clementine Mandarin Orchards. Crop Prot. 2011, 30, 1328–1333. [Google Scholar] [CrossRef]
- Aguilar-Fenollosa, E.; Pina, T.; Gómez-Martínez, M.A.; Hurtado, M.A.; Jacas, J.A. Does Host Adaptation of Tetranychus urticae Populations in Clementine Orchards with a Festuca arundinacea Cover Contribute to a Better Natural Regulation of This Pest Mite? Entomol. Exp. Appl. 2012, 144, 181–190. [Google Scholar] [CrossRef]
- Cruz-Miralles, J.; Guzzo, M.; Ibáñez-Gual, M.V.; Dembilio, Ó.; Jaques, J.A. Ground-Covers Affect the Activity Density of Ground-Dwelling Predators and Their Impact on the Mediterranean Fruit Fly, Ceratitis Capitata. BioControl 2022, 67, 583–592. [Google Scholar] [CrossRef]
- Parolin, P.; Bresch, C.; Desneux, N.; Brun, R.; Bout, A.; Boll, R.; Poncet, C. Secondary Plants Used in Biological Control: A Review. Int. J. Pest Manag. 2012, 58, 91–100. [Google Scholar] [CrossRef]
- Baggen, L.R.; Gurr, G.M. The Influence of Food On Copidosoma koehleri (Hymenoptera: Encyrtidae), and the Use of Flowering Plants as a Habitat Management Tool to Enhance Biological Control of Potato Moth, Phthorimaea operculella (Lepidoptera: Gelechiidae). Biol. Control 1998, 11, 9–17. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Moonen, A.C. Field Margins in Northern Europe: Their Functions and Interactions with Agriculture. Agric. Ecosyst. Environ. 2002, 89, 5–21. [Google Scholar] [CrossRef]
- Ambrosino, M.D.; Luna, J.M.; Jepson, P.C.; Wratten, S.D. Relative Frequencies of Visits to Selected Insectary Plants by Predatory Hoverflies (Diptera: Syrphidae), Other Beneficial Insects, and Herbivores. Environ. Entomol. 2006, 35, 394–400. [Google Scholar] [CrossRef]
- Carvell, C.; Meek, W.R.; Pywell, R.F.; Goulson, D.; Nowakowski, M. Comparing the Efficacy of Agri-Environment Schemes to Enhance Bumble Bee Abundance and Diversity on Arable Field Margins. J. Appl. Ecol. 2007, 44, 29–40. [Google Scholar] [CrossRef]
- Carvalheiro, L.G.; Seymour, C.L.; Nicolson, S.W.; Veldtman, R. Creating Patches of Native Flowers Facilitates Crop Pollination in Large Agricultural Fields: Mango as a Case Study. J. Appl. Ecol. 2012, 49, 1373–1383. [Google Scholar] [CrossRef]
- van Rijn, P.C.J.; Kooijman, J.; Wäckers, F.L. The Contribution of Floral Resources and Honeydew to the Performance of Predatory Hoverflies (Diptera: Syrphidae). Biol. Control 2013, 67, 32–38. [Google Scholar] [CrossRef]
- Blaauw, B.R.; Isaacs, R. Flower Plantings Increase Wild Bee Abundance and the Pollination Services Provided to a Pollination-Dependent Crop. J. Appl. Ecol. 2014, 51, 890–898. [Google Scholar] [CrossRef]
- Campbell, A.J.; Wilby, A.; Sutton, P.; Wäckers, F.L. Do Sown Flower Strips Boost Wild Pollinator Abundance and Pollination Services in a Spring-Flowering Crop? A Case Study from UK Cider Apple Orchards. Agric. Ecosyst. Environ. 2017, 239, 20–29. [Google Scholar] [CrossRef]
- Karamaouna, F.; Kati, V.; Volakakis, N.; Varikou, K.; Garantonakis, N.; Economou, L.; Birouraki, A.; Markellou, E.; Liberopoulou, S.; Edwards, M. Ground Cover Management with Mixtures of Flowering Plants to Enhance Insect Pollinators and Natural Enemies of Pests in Olive Groves. Agric. Ecosyst. Environ. 2019, 274, 76–89. [Google Scholar] [CrossRef]
- Kati, V.; Karamaouna, F.; Economou, L.; Mylona, P.V.; Samara, M.; Mitroiu, M.-D.; Barda, M.; Edwards, M.; Liberopoulou, S. Sown Wildflowers Enhance Habitats of Pollinators and Beneficial Arthropods in a Tomato Field Margin. Plants 2021, 10, 1003. [Google Scholar] [CrossRef]
- Karamaouna, F.; Kati, V.; Economou, L.; Troyanos, G.; Samara, M.; Liberopoulou, S.; Barda, M.; Mitroiu, M.-D.; Edwards, M. Selected Flowering Plants as a Habitat for Pollinators and Natural Enemies in Field Margins of a Watermelon Crop—Implications for Crop Yield. Int. J. Pest Manag. 2022, 0, 1–17. [Google Scholar] [CrossRef]
- Barda, M.; Karamaouna, F.; Kati, V.; Perdikis, D. Do Patches of Flowering Plants Enhance Insect Pollinators in Apple Orchards? Insects 2023, 14, 208. [Google Scholar] [CrossRef]
- Karpiński, T.M. Essential Oils of Lamiaceae Family Plants as Antifungals. Biomolecules 2020, 10, 103. [Google Scholar] [CrossRef]
- Xie, Y.; Tian, L.; Han, X.; Yang, Y. Research Advances in Allelopathy of Volatile Organic Compounds (VOCs) of Plants. Horticulturae 2021, 7, 278. [Google Scholar] [CrossRef]
- Hatt, S.; Xu, Q.; Francis, F.; Osawa, N. Aromatic Plants of East Asia to Enhance Natural Enemies towards Biological Control of Insect Pests. A Review. Entomol. Gen. 2019, 38, 275–315. [Google Scholar] [CrossRef]
- Schippmann, U.; Cunningham, A.B.; Leaman, D.J. Impact of Cultivation and Gathering of Medicinal Plants on Biodiversity: Global Trends and Issues. In Biodiversity and the Ecosystem Approach in Agriculture, Forestry and Fisheries; FAO: Rome, Italy, 2003; pp. 140–167. [Google Scholar]
- Hogg, B.N.; Bugg, R.L.; Daane, K.M. Attractiveness of Common Insectary and Harvestable Floral Resources to Beneficial Insects. Biol. Control 2011, 56, 76–84. [Google Scholar] [CrossRef]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea v. 1; CambridgeUniversity Press: Cambridge, UK, 1964. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea v. 2; CambridgeUniversity Press: Cambridge, UK, 1968. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea v. 3; CambridgeUniversity Press: Cambridge, UK, 1972. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea v. 4; CambridgeUniversity Press: Cambridge, UK, 1976. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea v. 5; CambridgeUniversity Press: Cambridge, UK, 1980. [Google Scholar]
- Goulet, H.; Huber, J.T. (Eds.) Hymenoptera of the World: An Identification Guide to Families; Centre for Land and Biological Resources Research: Ottawa, ON, Canada, 1993. [Google Scholar]
- Triplehorn, C.A.; Johnson, N.F.; Borror, D.J. Borror and Delong’s Introduction to the Study of Insects, 7th ed.; Thompson Brooks/Cole: Belmont, CA, USA, 2005. [Google Scholar]
- Gloor, D.; Blick, T.; Nentwig, W.; Kropf, C.; Hänggi, A. Spiders of Europe. Available online: https://www.araneae.nmbe.ch (accessed on 6 February 2023).
- SAS Institute Inc. JMP v7; SAS Institute Inc.: Cary, NC, USA. Available online: https://www.jmp.com/en_ph/company/aboutus.html (accessed on 6 February 2023).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Dafni, H.; Lensky, Y.; Fahn, A. Flower and Nectar Characteristics of Nine Species of Labiatae and Their Influence on Honeybee Visits. J. Apic. Res. 1988, 27, 103–114. [Google Scholar] [CrossRef]
- Wilson, H.; Miles, A.F.; Daane, K.M.; Altieri, M.A. Host Plant Associations of Anagrus spp. (Hymenoptera: Mymaridae) and Erythroneura elegantula (Hemiptera: Cicadellidae) in Northern California. Environ. Entomol. 2016, 45, 602–615. [Google Scholar] [CrossRef] [PubMed]
- Nickel, H.; Holzinger, W. Rapid Range Expansion of Ligurian Leafhopper, Eupteryx decemnotata Rey, 1891 (Hemiptera: Cicadellidae), a Potential Pest of Garden and Greenhouse Herbs, in Europe. Russ. Entomol. J. 2006, 15, 295–301. [Google Scholar]
- Tasi, A.; Lucky, A. Ligurian Leafhopper Eupteryx decemnotata (Rey) (Insecta: Hemiptera: Cicadellidae: Typhlocybinae): EENY750/IN1290, 3/2020. EDIS 2020, 2020, 4. [Google Scholar] [CrossRef]
- Orr, D.B.; Pleasants, J.M. The Potential of Native Prairie Plant Species to Enhance the Effectiveness of the Ostrinia Nubilalis Parasitoid Macrocentrus Grandii. J. Kans. Entomol. Soc. 1996, 69, 133–143. [Google Scholar]
- Mačukanović-Jocić, M.; Stevanović, Z.D.; Mladenović, M.; Jocić, G. Flower Morphophysiology of Selected Lamiaceae Species in Relation to Pollinator Attraction. J. Apic. Res. 2011, 50, 89–101. [Google Scholar] [CrossRef]
- Aronne, G.; Malara, P. Fiber-Optic Refractometer for in Vivo Sugar Concentration Measurements of Low-Nectar-Producing Flowers. New Phytol. 2019, 224, 987–993. [Google Scholar] [CrossRef]
- Samin, N.; Asgari, S. A study on the fauna of Scelionid wasps (Hymenoptera: Platygastroidea: Scelionidae) in the Isfahan province, Iran. Arch. Biol. Sci. 2012, 64, 1073–1077. [Google Scholar] [CrossRef]
- Garcia-Marí, F.; Vercher, R.; Costa-Comelles, J.; Marzal, C.; Villalba, M. Establishment of Citrostichus phyllocnistoides (Hymenoptera: Eulophidae) as a Biological Control Agent for the Citrus Leafminer Phyllocnistis citrella (Lepidoptera: Gracillariidae) in Spain. Biol. Control 2004, 29, 215–226. [Google Scholar] [CrossRef]
- Tsagkarakis, A.E.; Kalaitzaki, A.P.; Lykouressis, D.P. Phyllocnistis citrella and Its Parasitoids in Three Citrus Species in Greece. Phytoparasitica 2013, 41, 23–29. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Athanassiou, C.G.; Stathas, G.J.; Tomanović, Ž. Aphid Parasitoids (Hymenoptera: Braconidae: Aphidiinae) on Citrus: Seasonal Abundance, Association with the Species of Host Plant, and Sampling Indices. Phytoparasitica 2002, 30, 365–377. [Google Scholar] [CrossRef]
- Waloff, N.; Jervis, M.A. Communities of Parasitoids Associated with Leafhoppers and Planthoppers in Europe. In Advances in Ecological Research; Macfadyen, A., Ford, E.D., Eds.; Academic Press: Cambridge, MA, USA, 1987; Volume 17, pp. 281–376. [Google Scholar]
- Puig, J.; Gavara, J.; Ávalos Masó, J.A.; Soler, J.M.; García-Marí, F. Abundance of natural enemies in citrus of the Iberian Peninsula: Geographical distribution and seasonal trend. IOBC-WPRS Bull. 2018, 132, 68–75. [Google Scholar]
- Cotes, B.; González, M.; Benítez, E.; De Mas, E.; Clemente-Orta, G.; Campos, M.; Rodríguez, E. Spider Communities and Biological Control in Native Habitats Surrounding Greenhouses. Insects 2018, 9, 33. [Google Scholar] [CrossRef]
- Cardoso, P.; Pekár, S.; Jocqué, R.; Coddington, J.A. Global Patterns of Guild Composition and Functional Diversity of Spiders. PLoS ONE 2011, 6, e21710. [Google Scholar] [CrossRef]
- Bogya, S. Spiders (Araneae) as Polyphagous Natural Enemies in Orchards. Ph.D. Thesis, Wangenigen University & Research, Wangenigen, The Netherlands, 1999. [Google Scholar]
- Cherry, R.; Dowell, R.V. Predators of Citrus Blackfly [Hom.: Aleyrodidae]. Entomophaga 1979, 24, 385–391. [Google Scholar] [CrossRef]
- van den Berg, M.A.; Dippenaar-Shoeman, A.S.; Deacon, V.E.; Anderson, S.H. Interactions between Citrus Psylla, Trioza erytreae (Hem. Triozidae), and Spiders in an Unsprayed Citrus Orchard in the Transvaal Lowveld. Entomophaga 1992, 37, 599–608. [Google Scholar] [CrossRef]
- Smith, D.; Beattie, G.A.; Broadley, R. Citrus Pests and Their Natural Enemies: Integrated Pest Management in Australia; Department of Primary Industries: Brisbane, Australia, 1997. [Google Scholar]
- Amalin, D.M.; Reiskind, J.; Peña, J.E.; McSorley, R. Predatory Behavior of Three Species of Sac Spiders Attacking Citrus Leafminer. J. Arachnol. 2001, 29, 72–81. [Google Scholar] [CrossRef]
- Mansour, F.; Whitecomb, W.H. The Spiders of a Citrus Grove in Israel and Their Role as Biocontrol Agents Of Ceroplastes floridensis [Homoptera: Coccidae]. Entomophaga 1986, 31, 269–276. [Google Scholar] [CrossRef]
- Kaspi, R. Attraction of Female Chiracanthium mildei (Araneae: Clubionidae) to Olfactory Cues from Male Mediterranean Fruit Flies Ceratitis capitata (Diptera: Tephritidae). BioControl 2000, 45, 463–468. [Google Scholar] [CrossRef]
- Bogya, S.; Szinetár, C.S.; Markó, V. Species Composition of Spider (Araneae) Assemblages in Apple and Pear Orchards in the Carpathian Basin. Acta Phytopathol. Entomol. Hung. 1999, 34, 99–121. [Google Scholar]
- Ribeiro, C.; Cardoso, P.; Franco, J.C. Abundance and diversity of spiders in lemon orchards with different weed management systems. IOBC-WPRS Bull. 2006, 29, 167–177. [Google Scholar]
- Ghavami, S.; Amooz, S.G. Spider (Order Araneae) Fauna of Citrus Orchards in Northern Part of Iran. J. Appl. Biol. Sci. 2008, 2, 73–76. [Google Scholar]
- Barrientos, J.A.; Villalba Tordera, M.; Alvis Dávila, L.; García Mari, F. Identificación y abundancia de arañas (Araneae) en los cultivos de cítricos valencianos. Bol. San. Veg. Plagas 2010, 36, 69–85. [Google Scholar]
- Castañé, C.; Arnó, J.; Gabarra, R.; Alomar, O. Plant Damage to Vegetable Crops by Zoophytophagous Mirid Predators. Biol. Control 2011, 59, 22–29. [Google Scholar] [CrossRef]
- Bosco, L.; Tavella, L. Distribution and Abundance of Species of the Genus Orius in Horticultural Ecosystems of Northwestern Italy. Bull. Insect. 2013, 66, 297–307. [Google Scholar]
- Alomar, O.; Gabarra, R.; González, O.; Arnó, J. Selection of insectary plants for ecological infrastructure in Mediterranean vegetable crops. IOBC-WPRS Bull. 2006, 29, 5–8. [Google Scholar]
- Elekcïoğlu, N.Z. Species composition of Coccinellidae (Coleoptera) and their preys in Adana (Turkey) with observations on potential host medicinal and aromatic plants. Appl. Ecol. Environ. Res. 2020, 18, 369–388. [Google Scholar] [CrossRef]
- Pappas, M.L.; Broufas, G.D.; Koveos, D.S. Effects of Various Prey Species on Development, Survival and Reproduction of the Predatory Lacewing Dichochrysa prasina (Neuroptera: Chrysopidae). Biol. Control 2007, 43, 163–170. [Google Scholar] [CrossRef]
- Linn, C.A.; Griebeler, E.M. Habitat Preference of German Mantis religiosa Populations (Mantodea: Mantidae) and Implications for Conservation. Environ. Entomol. 2016, 45, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Romeu-Dalmau, C.; Piñol, J.; Agustí, N. Detecting Aphid Predation by Earwigs in Organic Citrus Orchards Using Molecular Markers. Bull. Entomol. Res. 2012, 102, 566–572. [Google Scholar] [CrossRef]
- Rodwell, L.E.; Day, J.J.; Foster, C.W.; Holloway, G.J. Daily Survival and Dispersal of Adult Rhagonycha fulva (Coleoptera: Cantharidae) in a Wooded Agricultural Landscape. Eur. J. Entomol. 2018, 115, 432–436. [Google Scholar] [CrossRef]
- Alvis, L.; Villalba, M.; Marzal, C.; García-Marí, F. Identification and abundance of Neuropteran species associated with citrus orchards in Valencia, Spain. IOBC-WPRS Bull. 2003, 26, 185–190. [Google Scholar]
- Miller, G.L.; Oswald, J.D.; Miller, D.R. Lacewings and Scale Insects: A Review of Predator/Prey Associations Between the Neuropterida and Coccoidea (Insecta: Neuroptera, Raphidioptera, Hemiptera). Ann. Entomol. Soc. Am. 2004, 97, 1103–1125. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Period April–June 2021 | Sampling Period April–June 2022 | |||
|---|---|---|---|---|
| Field | R | p | R | p |
| WV | 0.1372 | 0.0056 | 0.157 | 0.001 |
| RO(ORG) | 0.5026 | 0.0001 | 0.1447 | 0.0067 |
| RO(IPM) | 0.2631 | 0.0001 | 0.1793 | 0.0001 |
| OR | 0.2021 | 0.0005 | 0.1328 | 0.0003 |
| SG | 0.0736 | 0.0499 | 0.1344 | 0.0027 |
| SV | 0.3154 | 0.0001 | 0.0393 | 0.1738 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stathakis, T.; Economou, L.; Barda, M.; Angelioudakis, T.; Kati, V.; Karamaouna, F. Potential of Hedgerows with Aromatic Plants as Reservoirs of Natural Enemies of Pests in Orange Orchards. Insects 2023, 14, 391. https://doi.org/10.3390/insects14040391
Stathakis T, Economou L, Barda M, Angelioudakis T, Kati V, Karamaouna F. Potential of Hedgerows with Aromatic Plants as Reservoirs of Natural Enemies of Pests in Orange Orchards. Insects. 2023; 14(4):391. https://doi.org/10.3390/insects14040391
Chicago/Turabian StyleStathakis, Theodoros, Leonidas Economou, Myrto Barda, Theodoros Angelioudakis, Vaya Kati, and Filitsa Karamaouna. 2023. "Potential of Hedgerows with Aromatic Plants as Reservoirs of Natural Enemies of Pests in Orange Orchards" Insects 14, no. 4: 391. https://doi.org/10.3390/insects14040391