

Comparison of Preference for Chemicals Associated with Fruit Fermentation between Drosophila melanogaster and Drosophila suzukii and between Virgin and Mated D. melanogaster

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Y-Tube Attraction Test
2.3. EAG Experiments
2.4. Statistical Analysis
3. Results
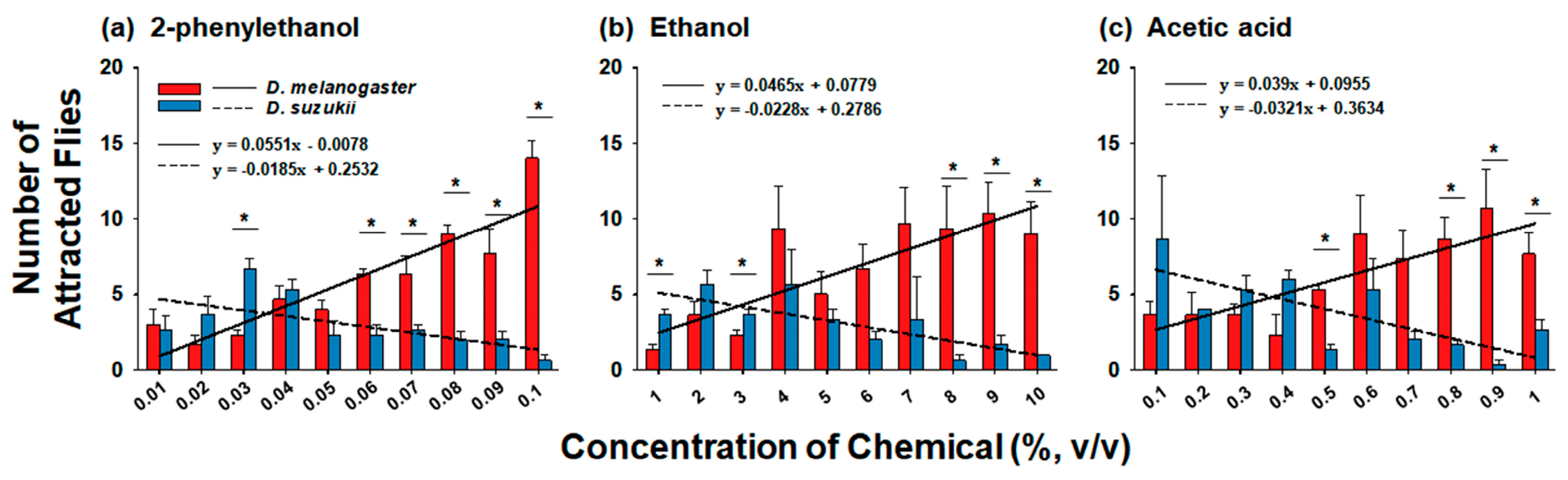
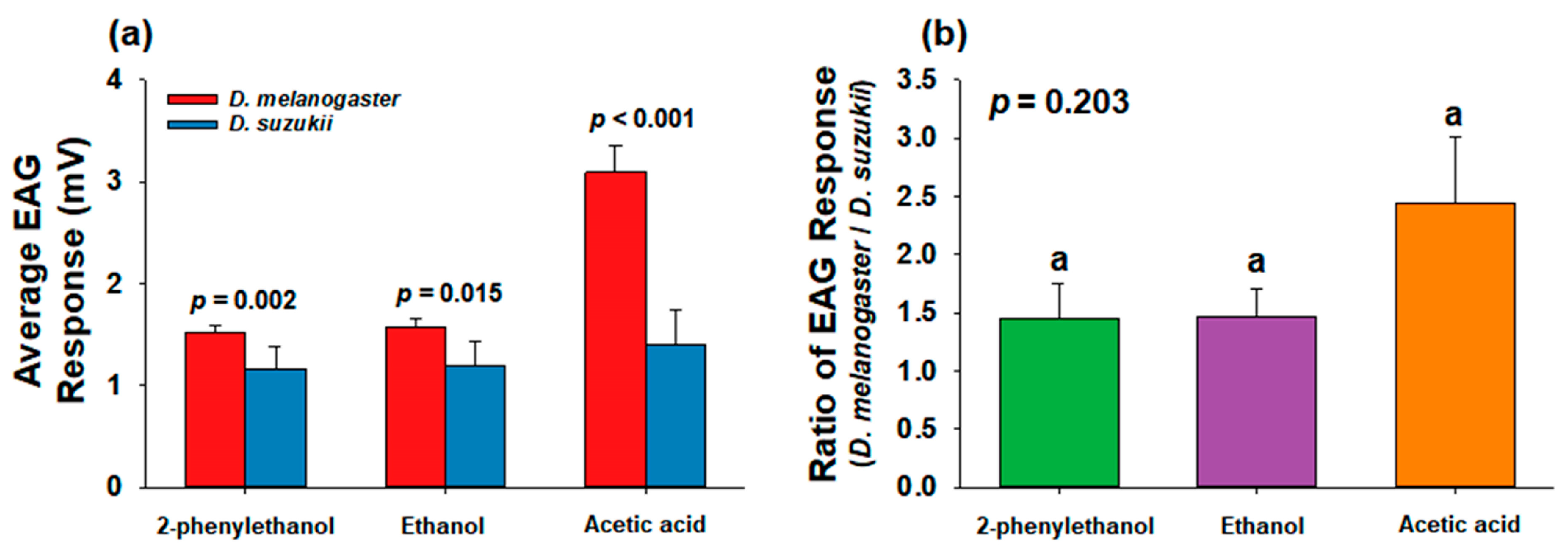
3.1. Comparison of Preference for Chemicals at Various Concentrations between D. melanogaster and D. suzukii
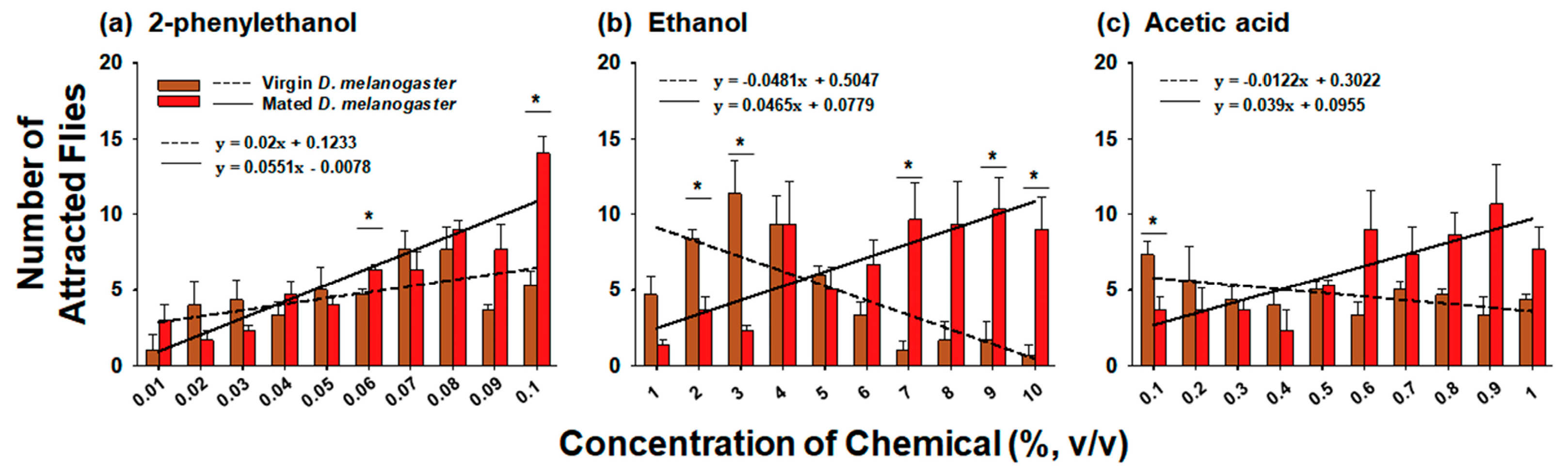
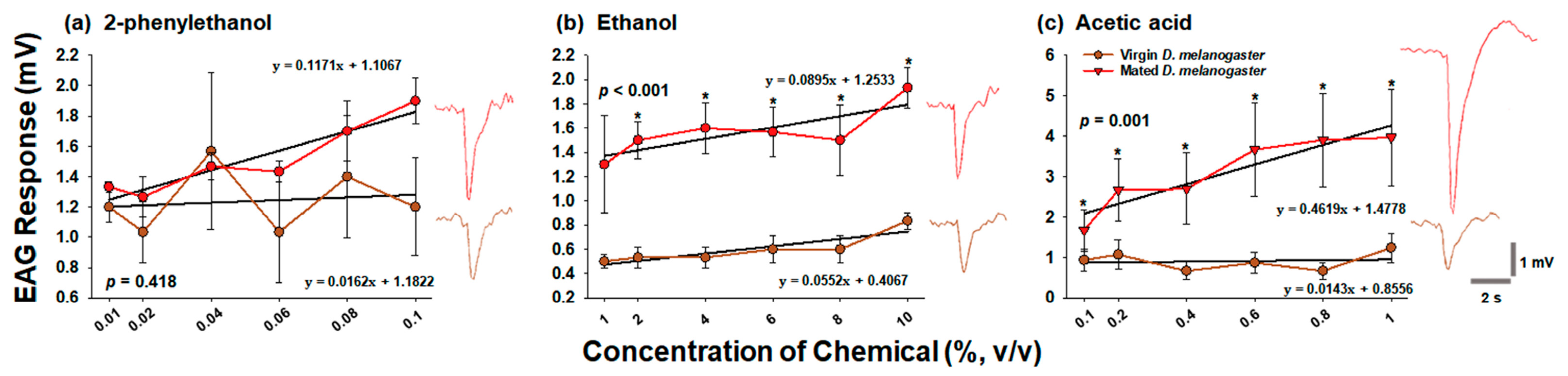
3.2. Comparison of Preference for Chemicals at Various Concentrations between Virgin and Mated D. melanogaster
4. Discussion
4.1. Comparison between D. melanogaster and D. suzukii
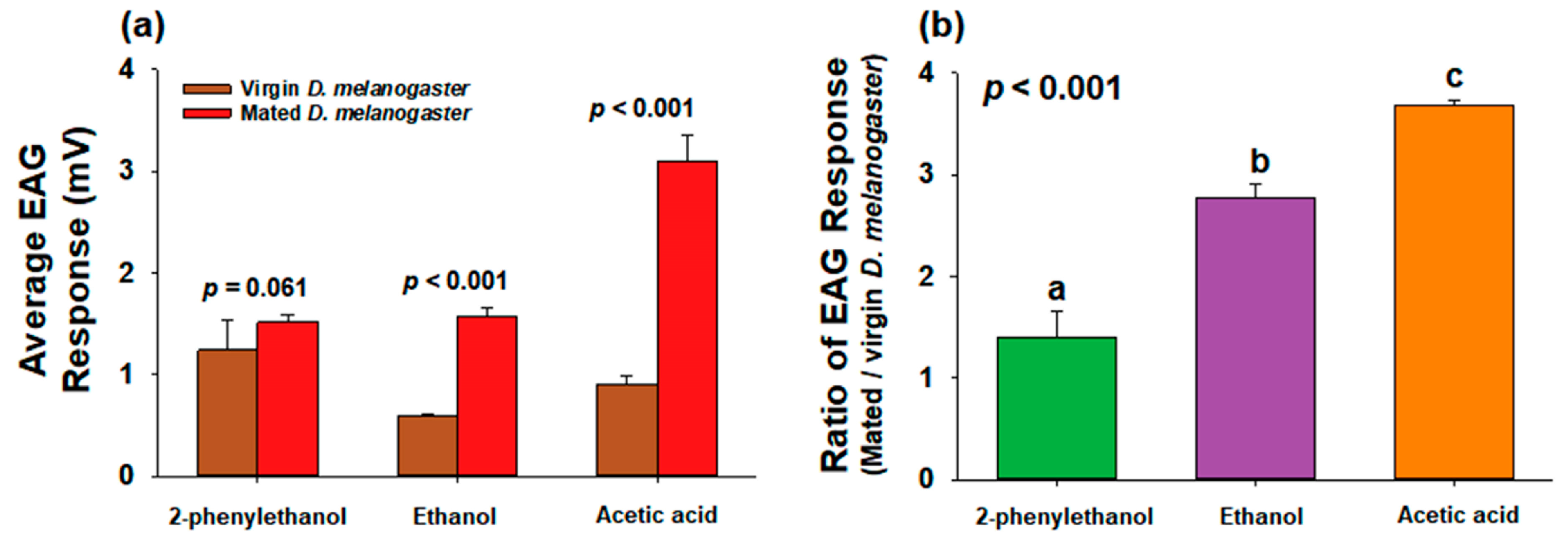
4.2. Comparison between Virgin and Mated D. melanogaster
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Keesey, I.W.; Zhang, J.; Depetris-Chauvin, A.; Obiero, G.F.; Gupta, A.; Gupta, N.; Vogel, H.; Knaden, M.; Hansson, B.S. Functional olfactory evolution in Drosophila suzukii and the subgenus Sophophora. Iscience 2022, 25, 104212. [Google Scholar] [CrossRef] [PubMed]
- Seetharam, A.S.; Stuart, G.W. Whole genome phylogeny for 21 Drosophila species using predicted 2b-RAD fragments. PeerJ 2013, 1, e226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Park, K.C.; Baker, T.C. Identification of odors from overripe mango that attract vinegar flies, Drosophila melanogaster. J. Chem. Ecol. 2003, 29, 899–909. [Google Scholar] [CrossRef]
- Rombaut, A.; Guilhot, R.; Xuéreb, A.; Benoit, L.; Chapuis, M.P.; Gibert, P.; Fellous, S. Invasive Drosophila suzukii facilitates Drosophila melanogaster infestation and sour rot outbreaks in the vineyards. R. Soc. Open Sci. 2017, 4, 170117. [Google Scholar] [CrossRef] [Green Version]
- Cha, D.H.; Adams, T.; Werle, C.T.; Sampson, B.J.; Adamczyk, J.J., Jr.; Rogg, H.; Landolt, P.J. A four-component synthetic attractant for Drosophila suzukii (Diptera: Drosophilidae) isolated from fermented bait headspace. Pest Manag. Sci. 2014, 70, 324–331. [Google Scholar] [CrossRef]
- Bengtsson, M.; Jaastad, G.; Knudsen, G.; Kobro, S.; Bäckman, A.C.; Pettersson, E.; Witzgall, P. Plant volatiles mediate attraction to host and non-host plant in apple fruit moth, Argyresthia conjugella. Entomol. Exp. Appl. 2006, 118, 77–85. [Google Scholar] [CrossRef]
- Mattheis, J.P.; Buchanan, D.A.; Fellman, J.K. Volatile compounds emitted by sweet cherries (Prunus avium Cv. Bing) during fruit development and ripening. J. Agric. Food Chem. 1992, 40, 471–474. [Google Scholar] [CrossRef]
- Levey, D.J. The evolutionary ecology of ethanol production and alcoholism. Integr. Comp. Biol. 2004, 44, 284–289. [Google Scholar] [CrossRef]
- Palanca, L.; Gaskett, A.C.; Günther, C.S.; Newcomb, R.D.; Goddard, M.R. Quantifying variation in the ability of yeasts to attract Drosophila melanogaster. PLoS ONE 2013, 8, e75332. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, M.; Franke, K.; Fischer, K. Feeding on ripening and over-ripening fruit: Interactions between sugar, ethanol and polyphenol contents in a tropical butterfly. J. Exp. Biol. 2017, 220, 3127–3134. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, A.; Alquézar, B.; Peña, L. Fruit aromas in mature fleshy fruits as signals of readiness for predation and seed dispersal. New Phytol. 2013, 197, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Dudley, R. Ethanol, fruit ripening, and the historical origins of human alcoholism in primate frugivory. Integr. Comp. Biol. 2004, 44, 315–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montooth, K.L.; Siebenthall, K.T.; Clark, A.G. Membrane lipid physiology and toxin catabolism underlie ethanol and acetic acid tolerance in Drosophila melanogaster. J. Exp. Biol. 2006, 209, 3837–3850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Lee, S.; Kim, Y.H.; Kim, Y.H. Comparative analyses of susceptibility to chemicals associated with fermentation between Drosophila melanogaster and Drosophila suzukii. Entomol. Res. 2018, 48, 514–521. [Google Scholar] [CrossRef]
- Kim, Y.; Goh, G.; Kim, Y.H. Expression of antimicrobial peptides associated with different susceptibilities to environmental chemicals in Drosophila suzukii and Drosophila melanogaster. Pestic. Biochem. Physiol. 2022, 187, 105210. [Google Scholar] [CrossRef]
- Devineni, A.V.; Heberlein, U. Preferential ethanol consumption in Drosophila models features of addiction. Curr. Biol. 2009, 19, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Ogueta, M.; Cibik, O.; Eltrop, R.; Schneider, A.; Scholz, H. The influence of Adh function on ethanol preference and tolerance in adult Drosophila melanogaster. Chem. Senses. 2010, 35, 813–822. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, P.; Kim, A.Y.; Jung, J.K.; Donahue, K.M.; Jung, C.; Choi, M.Y.; Koh, Y.H. The biochemical adaptations of spotted wing drosophila (Diptera: Drosophilidae) to fresh fruits reduced fructose concentrations and glutathione-S transferase activities. J. Econ. Entomol. 2016, 109, 973–981. [Google Scholar] [CrossRef]
- Belloni, V.; Galeazzi, A.; Bernini, G.; Mandrioli, M.; Versace, E.; Haase, A. Evolutionary compromises to metabolic toxins: Ammonia and urea tolerance in Drosophila suzukii and Drosophila melanogaster. Physiol. Behav. 2018, 191, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Karageorgi, M.; Bräcker, L.B.; Lebreton, S.; Minervino, C.; Cavey, M.; Siju, K.P.; Grunwald Kadow, I.C.G.; Gompel, N.; Prud’homme, B. Evolution of Multiple Sensory Systems Drives Novel Egg-Laying Behavior in the Fruit Pest Drosophila suzukii. Curr. Biol. 2017, 27, 847–853. [Google Scholar] [CrossRef] [Green Version]
- Durkin, S.M.; Chakraborty, M.; Abrieux, A.; Lewald, K.M.; Gadau, A.; Svetec, N.; Peng, J.; Kopyto, M.; Langer, C.B.; Chiu, J.C.; et al. Behavioral and genomic sensory adaptations underlying the pest activity of Drosophila suzukii. Mol. Biol. Evol. 2021, 38, 2532–2546. [Google Scholar] [CrossRef] [PubMed]
- Keesey, I.W.; Knaden, M.; Hansson, B.S. Olfactory specialization in Drosophila suzukii supports an ecological shift in host preference from rotten to fresh fruit. J. Chem. Ecol. 2015, 41, 121–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cloonan, K.R.; Abraham, J.; Angeli, S.; Syed, Z.; Rodriguez-Saona, C. Advances in the chemical ecology of the spotted wing drosophila (Drosophila suzukii) and its applications. J. Chem. Ecol. 2018, 44, 922–939. [Google Scholar] [CrossRef] [PubMed]
- Dalton, D.T.; Walton, V.M.; Shearer, P.W.; Walsh, D.B.; Caprile, J.; Isaacs, R. Laboratory survival of Drosophila suzukii under simulated winter conditions of the Pacific Northwest and seasonal field trapping in five primary regions of small and stone fruit production in the United States. Pest Manag. Sci. 2011, 67, 1368–1374. [Google Scholar] [CrossRef]
- Clymans, R.; Van Kerckvoorde, V.; Bangels, E.; Akkermans, W.; Alhmedi, A.; De Clercq, P.; Beliën, T.; Bylemans, D. Olfactory preference of Drosophila suzukii shifts between fruit and fermentation cues over the season: Effects of physiological status. Insects 2019, 10, 200. [Google Scholar] [CrossRef] [Green Version]
- Manning, A. The sexual behaviour of two sibling Drosophila species. Behaviour 1960, 15, 123–145. [Google Scholar] [CrossRef]
- Seong, K.M.; Kim, Y.; Kim, D.; Pittendrigh, B.R.; Kim, Y.H. Identification of transcriptional responsive genes to acetic acid, ethanol, and 2-phenylethanol exposure in Drosophila melanogaster. Pestic. Biochem. Physiol. 2020, 165, 104552. [Google Scholar] [CrossRef]
- Cha, D.H.; Adams, T.; Rogg, H.; Landolt, P.J. Identification and field evaluation of fermentation volatiles from wine and vinegar that mediate attraction of spotted wing drosophila, Drosophila suzukii. J. Chem. Ecol. 2012, 38, 1419–1431. [Google Scholar] [CrossRef]
- Becher, P.G.; Bengtsson, M.; Hansson, B.S.; Witzgall, P. Flying the fly: Long-range flight behavior of Drosophila melanogaster to attractive odors. J. Chem. Ecol. 2010, 36, 599–607. [Google Scholar] [CrossRef] [Green Version]
- Dweck, H.K.; Talross, G.J.; Wang, W.; Carlson, J.R. Evolutionary shifts in taste coding in the fruit pest Drosophila suzukii. eLife 2021, 10, e64317. [Google Scholar] [CrossRef]
- Ai, M.; Min, S.; Grosjean, Y.; Leblanc, C.; Bell, R.; Benton, R.; Suh, G.S. Acid sensing by the Drosophila olfactory system. Nature 2010, 468, 691–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, R.M.; Devineni, A.V.; King, I.F.; Heberlein, U. Oviposition preference for and positional avoidance of acetic acid provide a model for competing behavioral drives in Drosophila. Proc. Natl. Acad. Sci. USA 2009, 106, 11352–11357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, G.; Huang, J.H.; McMullen, J.G.; Newell, P.D.; Douglas, A.E. Physiological responses of insects to microbial fermentation products: Insights from the interactions between Drosophila and acetic acid. J. Insect Physiol. 2018, 106, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Xu, S.; Li, Y.; Shi, G. Improvement of 2-phenylethanol production in Saccharomyces cerevisiae by evolutionary and rational metabolic engineering. PLoS ONE 2021, 16, e0258180. [Google Scholar] [CrossRef]
- Clark, G.S. Leaf alcohol. Perfum. Flavor. 1990, 15, 47–52. [Google Scholar]
- Zentou, H.; Zainal Abidin, Z.; Yunus, R.; Awang Biak, D.R.; Abdullah Issa, M.; Yahaya Pudza, M. A new model of alcoholic fermentation under a byproduct inhibitory effect. ACS Omega 2021, 6, 4137–4146. [Google Scholar] [CrossRef]
- Saichana, N.; Matsushita, K.; Adachi, O.; Frébort, I.; Frebortova, J. Acetic acid bacteria: A group of bacteria with versatile biotechnological applications. Biotechnol. Adv. 2015, 33, 1260–1271. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Kim, Y.; Roh, G.H.; Kim, Y.H. Comparison of Preference for Chemicals Associated with Fruit Fermentation between Drosophila melanogaster and Drosophila suzukii and between Virgin and Mated D. melanogaster. Insects 2023, 14, 382. https://doi.org/10.3390/insects14040382
Kim H, Kim Y, Roh GH, Kim YH. Comparison of Preference for Chemicals Associated with Fruit Fermentation between Drosophila melanogaster and Drosophila suzukii and between Virgin and Mated D. melanogaster. Insects. 2023; 14(4):382. https://doi.org/10.3390/insects14040382
Chicago/Turabian StyleKim, Hyemin, YeongHo Kim, Gwang Hyun Roh, and Young Ho Kim. 2023. "Comparison of Preference for Chemicals Associated with Fruit Fermentation between Drosophila melanogaster and Drosophila suzukii and between Virgin and Mated D. melanogaster" Insects 14, no. 4: 382. https://doi.org/10.3390/insects14040382