
Climate and Environmental Changes and Their Potential Effects on the Dynamics of Chagas Disease: Hybridization in Rhodniini (Hemiptera, Triatominae)

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Crosses
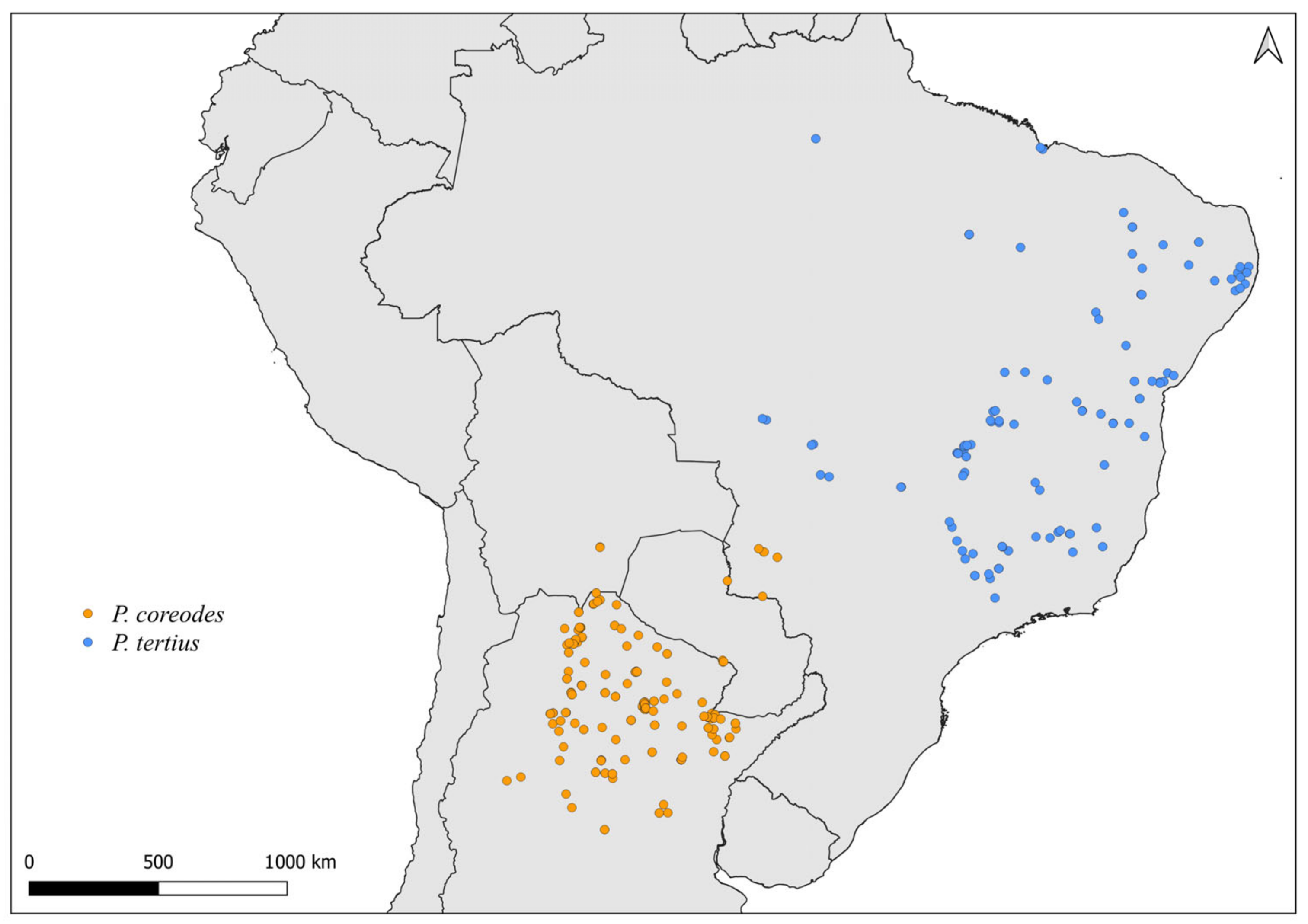
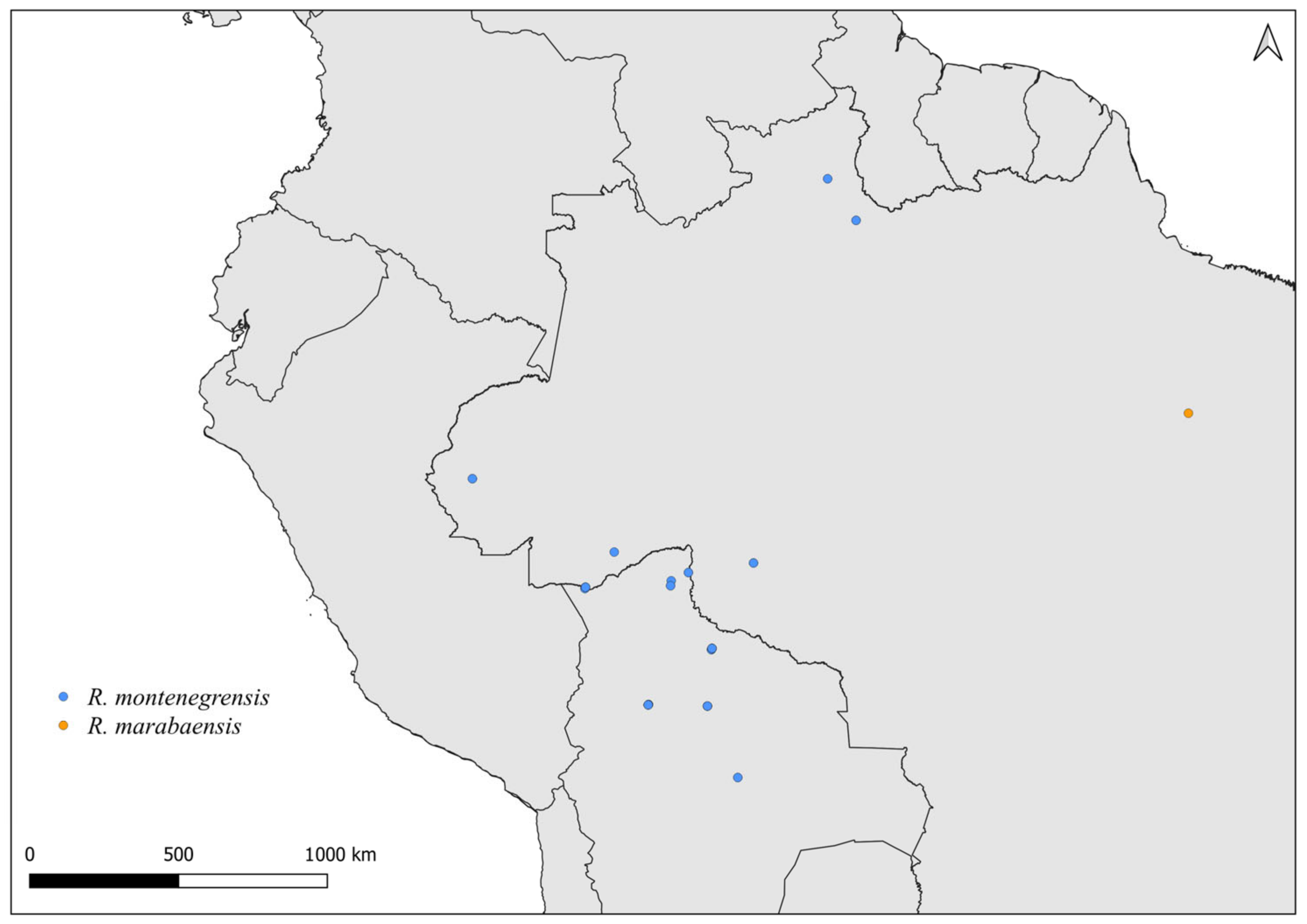
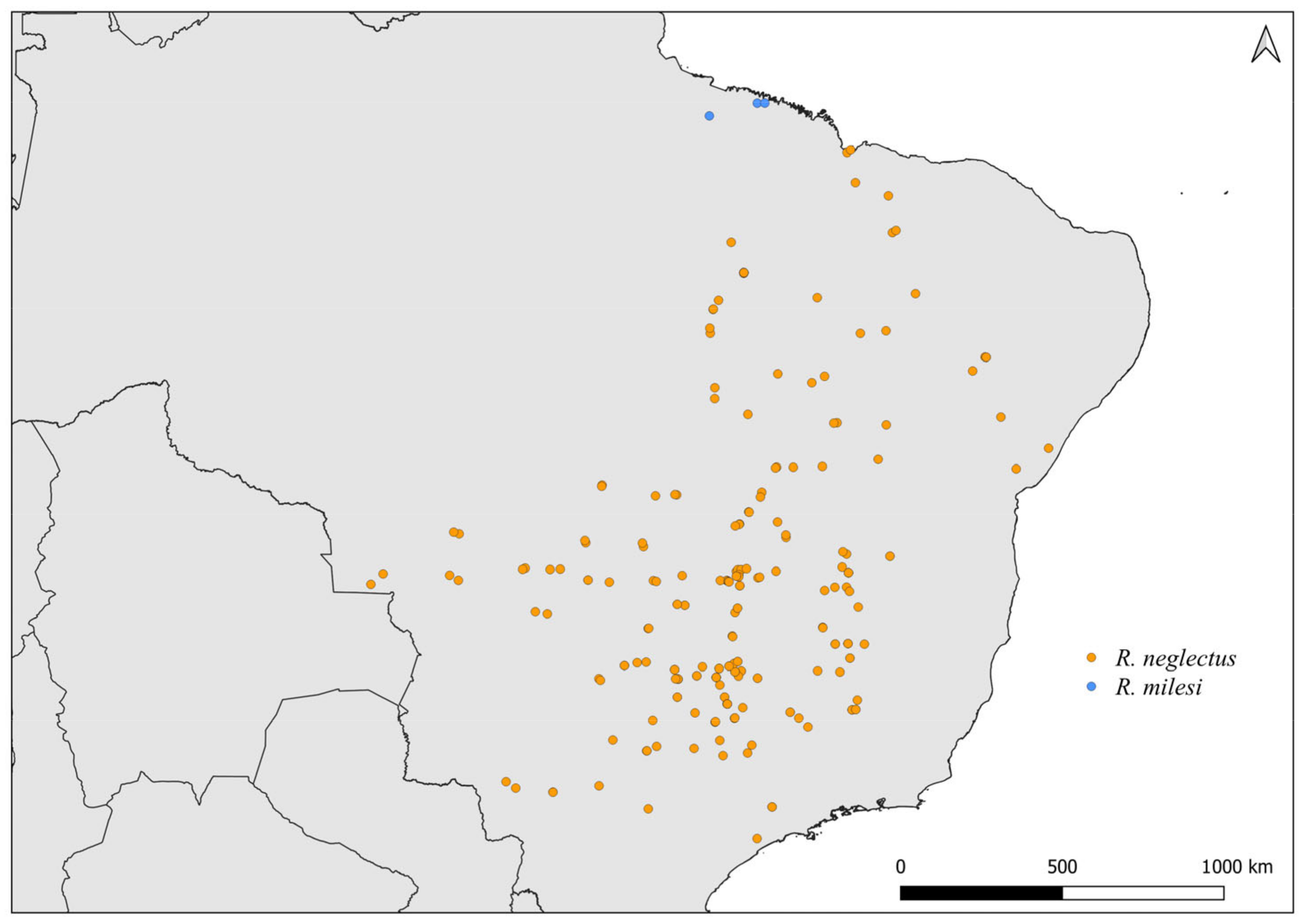
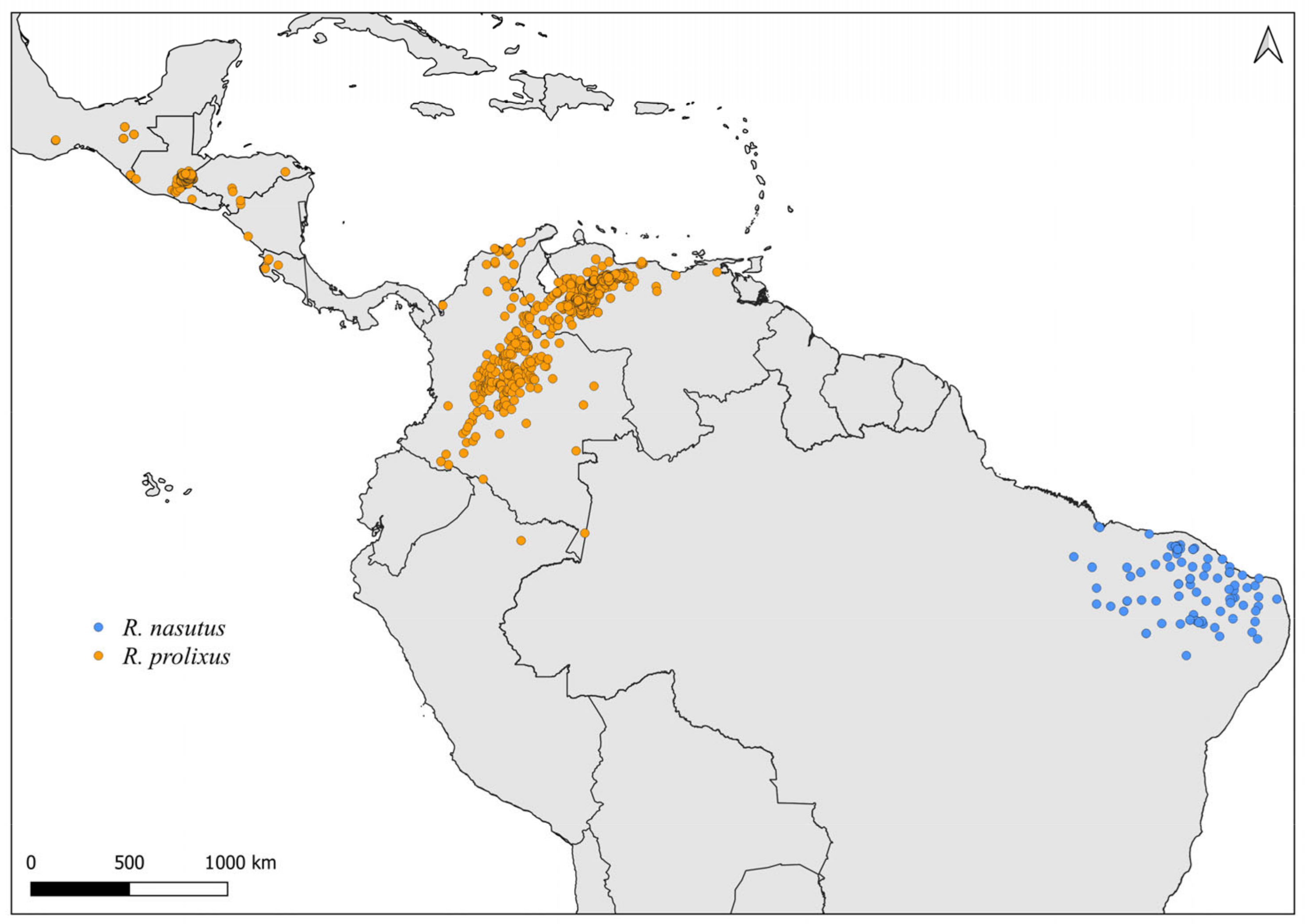
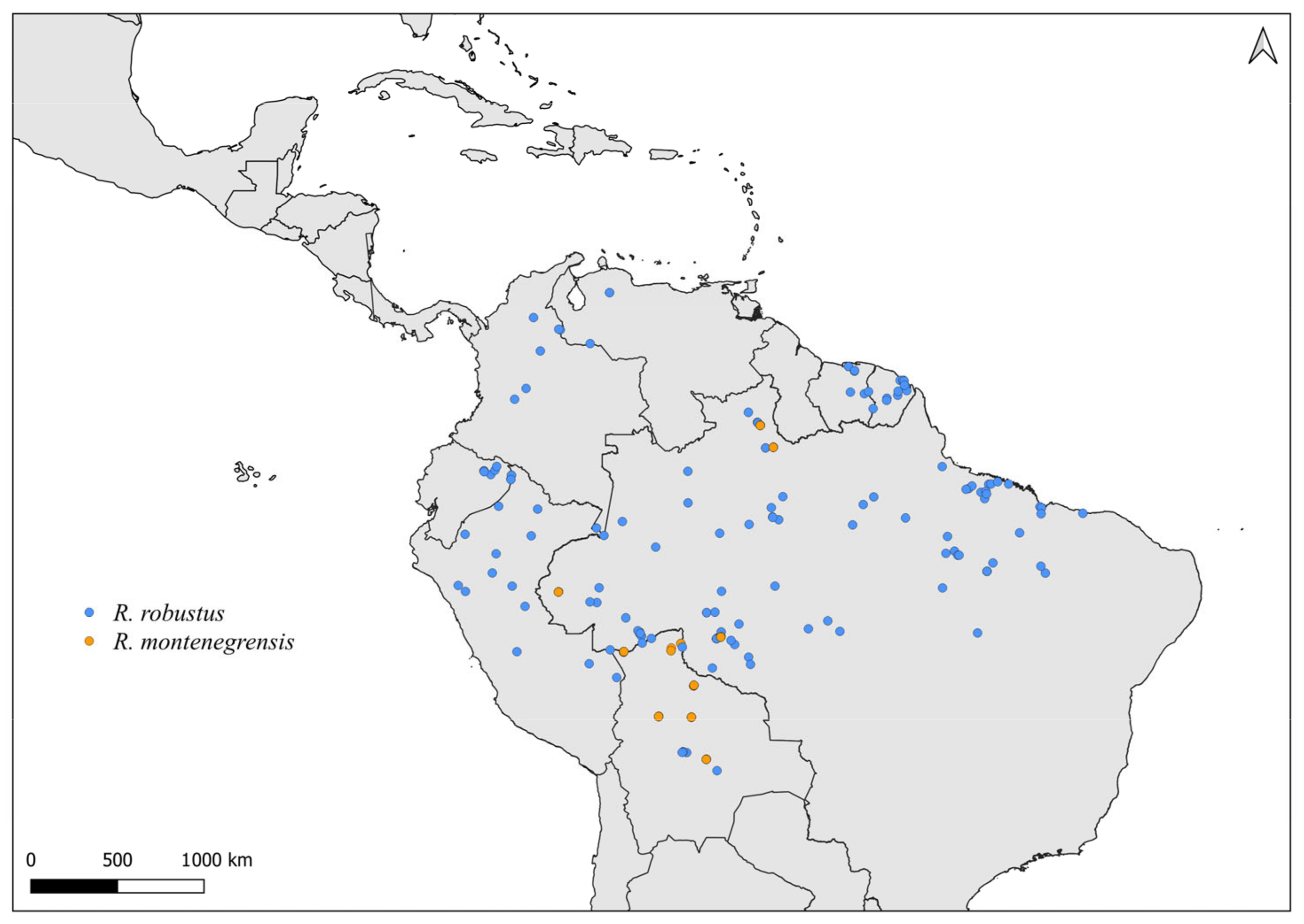
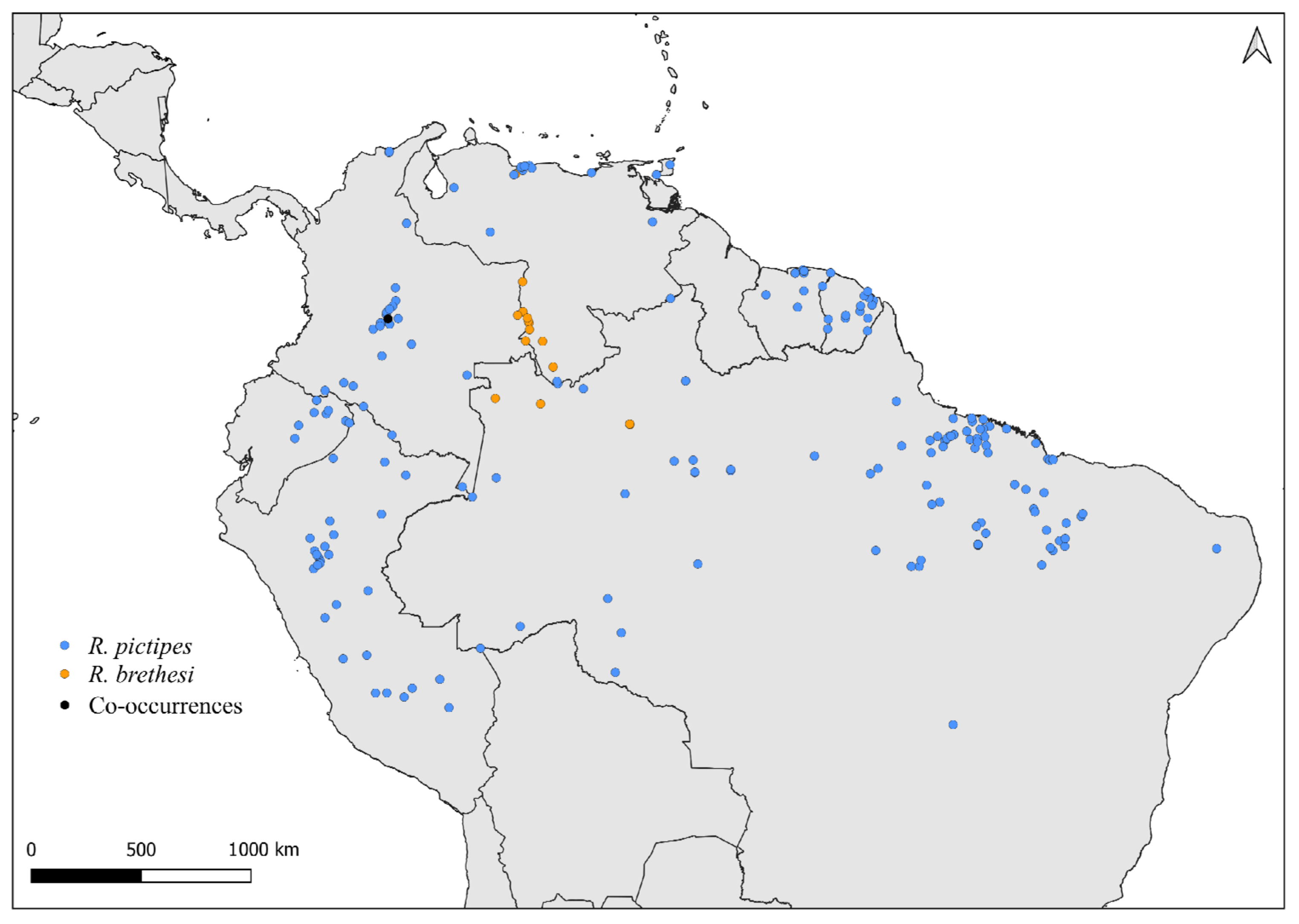
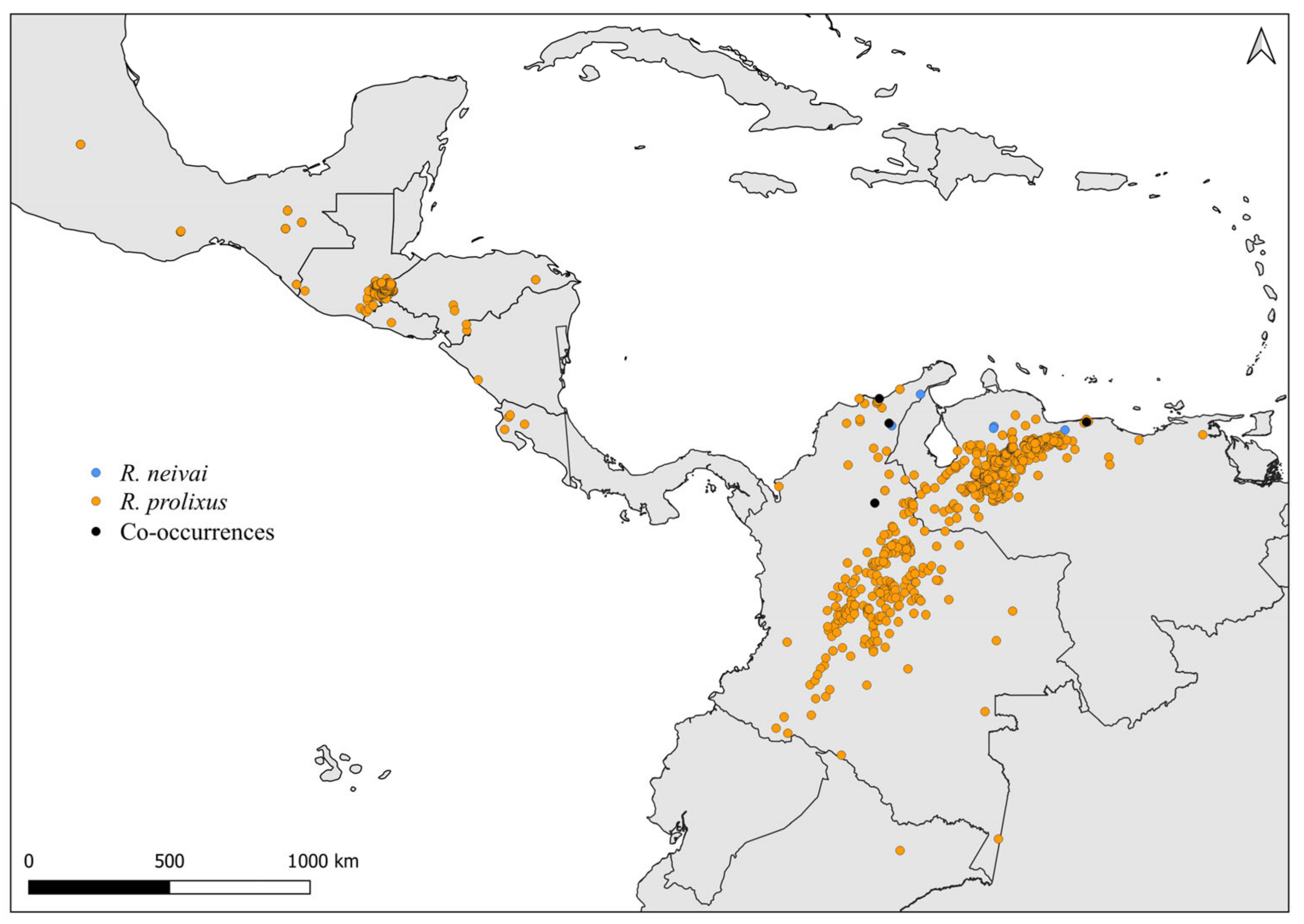
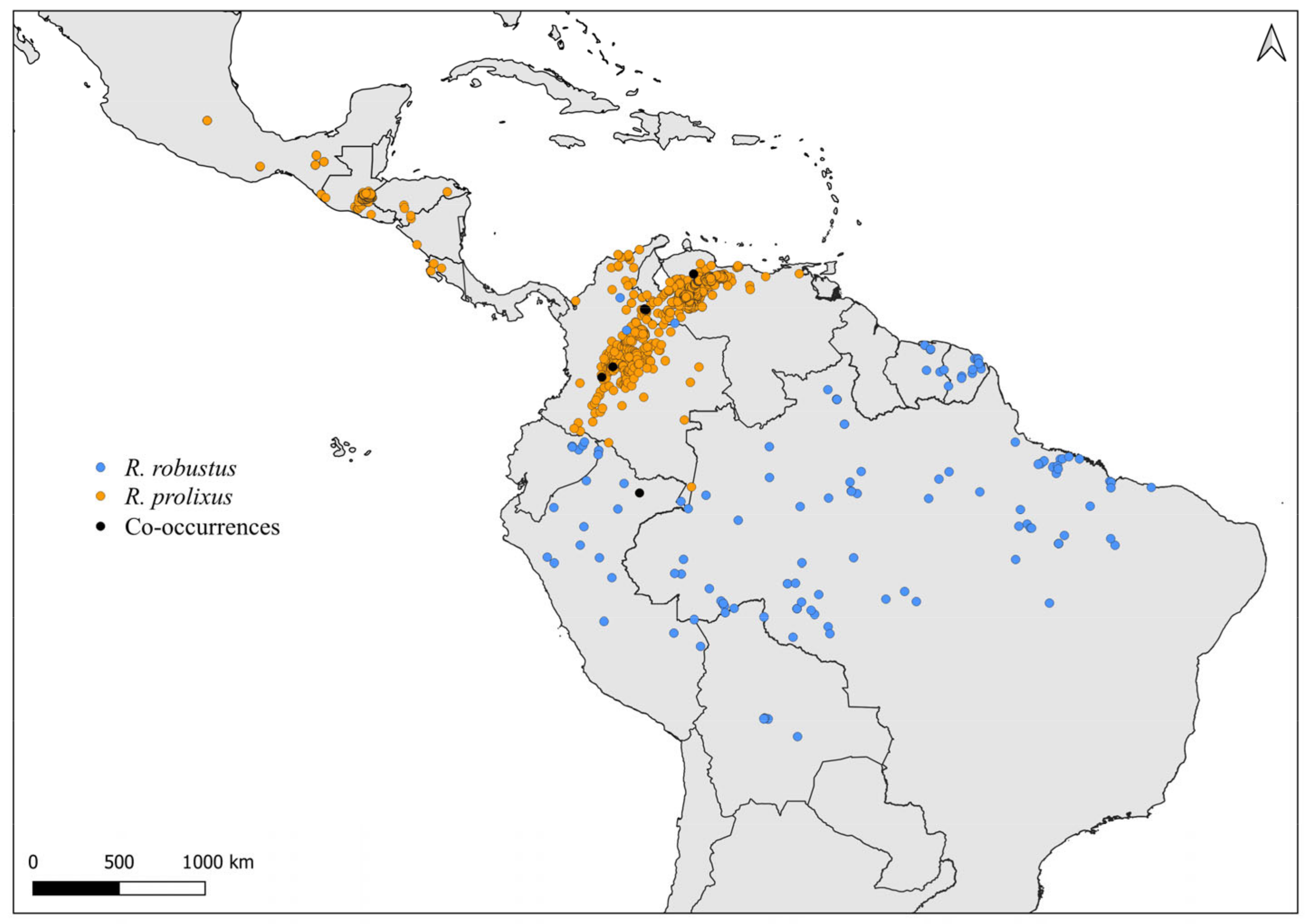
2.2. Distribution Maps
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Available online: https://www.who.int/health-topics/neglected-tropical-diseases#tab=tab_1 (accessed on 18 January 2023).
- Hotez, P.J.; Alvarado, M.; Basáñez, M.C.; Bolliger, I.; Bourne, R.; Boussinesq, M.; Broker, S.J.; Brown, A.S.; Buckle, G.; Budke, C.M.; et al. The global burden of disease study 2010: Interpretation and implications for the neglected tropical diseases. PLoS Negl. Trop. Dis. 2014, 8, e2865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Available online: https://apps.who.int/iris/handle/10665/77950 (accessed on 18 January 2023).
- World Health Organization. Available online: http://apps.who.int/iris/bitstream/handle/10665/44440/9789241564090_eng.pdf?sequence=1 (accessed on 18 January 2023).
- Martins-Melo, F.R.; Ramos, A.N., Jr.; Alencar, C.H.; Heukelbach, J. Prevalence of Chagas disease in Brazil: A systematic review and meta-analysis. Acta Trop. 2014, 130, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molyneux, D.H.; Savioli, L.; Engels, D. Neglected tropical diseases: Progress towards addressing the chronic pandemic. Lancet 2017, 389, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Conteh, L.; Engels, T.; Molyneux, D.H. Socioeconomic aspects of neglected tropical diseases. Lancet 2010, 375, 239–247. [Google Scholar] [CrossRef]
- Hotez, P.J. Ten global “hotspots” for the neglected tropical diseases. PLoS Negl. Trop. Dis. 2014, 8, e2496. [Google Scholar] [CrossRef]
- Cable, J.; Barber, I.; Boag, B.; Ellison, A.R.; Morgan, E.R.; Murray, K.; Pascoe, E.L.; Sait, S.M.; Wilson, A.J.; Booth, M. Global change, parasite transmission and disease control: Lessons from ecology. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160088. [Google Scholar] [CrossRef] [Green Version]
- Tidman, R.; Abela-Ridder, B.; Castañeda, R.R. The impact of climate change on neglected tropical diseases: A systematic review. Trans. R. Soc. Trop. Med. Hyg. 2021, 115, 147–168. [Google Scholar] [CrossRef]
- Booth, M. Climate Change and the Neglected Tropical Diseases. Adv. Parasitol. 2018, 100, 39–126. [Google Scholar]
- Rassi-Junior, A.; Rassi, A.; Marin-Neto, J.A. Chagas disease. Lancet 2010, 375, 1388–1402. [Google Scholar] [CrossRef]
- World Health Organization. Available online: http://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 18 January 2023).
- Pacheco-Tucuch, F.S.; Ramirez-Sierra, M.J.; Gourbière, S.; Dumonteil, E. Public street lights increase house infestation by Triatoma dimidiata, vector of Chagas disease in the Yucatan peninsu. PLoS ONE Public Libr. Sci. 2012, 7, e36207. [Google Scholar]
- Gottdenker, N.L.; Chaves, L.F.; Calzada, J.E.; Saldaña, A.; Carrol, C.R. Host life history strategy, species diversity, and habitat influence Trypanosoma cruzi vector infection in Changing landscapes. PLoS Negl. Trop. Dis. 2012, 6, e1884. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro-Júnior, G.; Gurgel-Gonçalves, R.; Reis, R.B.; Santos, C.G.S.; Amorim, A.; Andrade, S.G.; Reis, M.G. Frequent house invasion of Trypanosoma cruzi-infected triatomines in a suburban area of Brazil. PLoS Negl. Trop. Dis. 2015, 9, e0003678. [Google Scholar]
- Rojas-Cortez, M.; Pinazo, M.J.; Garcia, L.; Arteaga, M.; Uriona, L.; Gamboa, S.; Mejía, C.; Lozano, D.; Gascon, J.; Torrico, F.; et al. Trypanosoma cruzi-infected Panstrongylus geniculatus and Rhodnius robustus adults invade households in the tropics of Cochabamba region of Bolivia. Parasites Vectors 2016, 9, 158. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.N.; O’Day, S.; Fisher-Hoch, S.; Gorchakov, R.; Patino, R.; Feria Arroyo, T.P.; Laing, S.T.; Lopez, J.E.; Ingber, A.; Jones, K.M.; et al. One Health interactions of Chagas disease vectors, canid hosts, and human residents along the Texas-Mexico border. PLoS Negl. Trop. Dis. 2016, 10, e0005074. [Google Scholar] [CrossRef] [Green Version]
- Curto de Casas, S.I.; Carcavallo, R.U.; Mena Segura, C.A.; Galíndez Girón, I. Bioclimatic factors of Triatominae distribution. Useful techniques for studies on climate change. Entomol. Y Vectores 1994, 1, 51–67. [Google Scholar]
- Galvão, C.; Jurberg, J.; Carcavallo, R.U.; Mena Segura, C.A.; Galíndez Girón, I.; Curto de Casas, S.I. Distribuição geográfica e dispersão alti-latitudinal de alguns gêneros e espécies da tribo Triatomini Jeannel, 1919 (Hemiptera, Reduviidae, Triatominae). Mem. Inst. Oswaldo Cruz 1998, 93, 33–37. [Google Scholar] [CrossRef]
- Rocklöv, J.; Dubrow, R. Climate change: An enduring challenge for vector-borne disease prevention and control. Nat. Immunol. 2020, 21, 479–483. [Google Scholar] [CrossRef]
- Reiter, P. Climate change and mosquito-borne disease. Environ. Health Perspect. 2001, 109, 141–161. [Google Scholar]
- Chaves, L.F.; Koenraadt, C.J. Climate change and highland malaria: Fresh air for a hot debate. Q. Rev. Biol. 2010, 85, 27–55. [Google Scholar] [CrossRef] [Green Version]
- Campbell-Lendrum, D.; Manga, L.; Bagayoko, M.; Sommerfeld, J. Climate change and vector-borne diseases: What are the implications for public health research and policy? Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20130552. [Google Scholar] [CrossRef] [Green Version]
- Ceccarelli, S.; Rabinovich, J.E. Global climate change effects on Venezuela’s vulnerability to Chagas disease is linked to the geographic distribution of five triatomine species. J. Med. Entomol. 2015, 52, 1333–1343. [Google Scholar] [CrossRef] [PubMed]
- Medone, P.; Ceccarelli, S.; Parham, P.E.; Figureuera, A.; Rabinovich, J.E. The impact of climate change on the geographical distribution of two vectors of Chagas disease: Implications for the force of infection. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20130560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido, R.; Bacigalupo, A.; Peña-Gómez, F.; Bustamente, R.O.; Cattan, P.E.; Gorla, D.; Botto-Mahan, C. Potential impact of climate change on the geographical distribution of two wild vectors of Chagas disease in Chile: Mepraia spinolai and Mepraia gajardoi. Parasites Vector 2019, 12, 478. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Castro, O.; Moo-Llanes, D.A.; Ramsey, J.M. Impact of climate change on vector transmission of Trypanosoma cruzi (Chagas, 1909) in North America. Med. Vet. Entomol. 2018, 32, 84–101. [Google Scholar] [CrossRef] [PubMed]
- Garza, M.; Feria Arroyo, T.P.; Casillas, E.A.; Sanchez-Cordero, V.; Rivaldi, C.L.; Sarkar, S. Projected Future Distributions of Vectors of Trypanosoma cruzi in North America under Climate Change Scenarios. PLoS Negl. Trop. Dis. 2014, 8, e2818. [Google Scholar] [CrossRef]
- Lambert, R.C.; Kolivras, K.N.; Resler, L.M.; Brewster, C.C.; Paulson, S.L. The potential for emergence of Chagas disease in the United States. Geospat. Health 2008, 2, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.; Dornak, L.L.; Almeida, C.E.; Peterson, A.T. Distributional potential of the Triatoma brasiliensis species complex at present and under scenarios of future climate conditions. Parasites Vectors 2014, 7, 238. [Google Scholar] [CrossRef]
- Tapia-Garay, V.; Figueroa, D.P.; Maldonado, A.; Frías-Laserre, D.; Gonzalez, C.R.; Parra, A.; Canals, L.; Apt, W.; Alvarado, S.; Caceres, D.; et al. Assessing the risk zones of Chagas disease in Chile, in a world marked by global climatic change. Mem. Inst. Oswaldo Cruz 2018, 113, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Eberhard, F.E.; Cunze, S.; Kochmann, J.; Klimpel, S. Modelling the climatic suitability of Chagas disease vectors on a global scale. Elife 2020, 9, e52072. [Google Scholar] [CrossRef]
- Graves, B.N. Climate Change and Chagas Disease in the Americas: A Qualitative Systematic Review. Available online: https://digitalcommons.library.tmc.edu/uthsph_dissertsopen/87 (accessed on 24 February 2023).
- Chagas, C. Nova tripanozomiaze humana: Estudos sobre a morfolojia e o ciclo evolutivo do Schizotrypanum cruzi n. gen., n. sp., ajente etiolojico de nova entidade morbida do homem. Mem. Inst. Oswaldo Cruz 1909, 1, 159–218. [Google Scholar] [CrossRef] [Green Version]
- Cordovez, J.M.; Rendon, L.M.; Gonzalez, C.; Guhl, F. Using the basic reproduction number to assess the effects of climate change in the risk of Chagas disease transmission in Colombia. Acta Trop. 2014, 129, 74–82. [Google Scholar] [CrossRef]
- Wiegand, K.M. A taxonomist’s experience with hybrids in the wild. Science 1935, 81, 161–166. [Google Scholar] [CrossRef]
- Chunco, A.J. Hybridization in a warmer world. Ecol. Evol. 2014, 4, 2019–2031. [Google Scholar] [CrossRef]
- Kardol, P.; Campany, C.E.; Souza, L.; Norby, R.J.; Weltzin, J.F.; Classen, A.T. Climate change effects on plant biomass alter dominance patterns and community evenness in an experimental old-field ecosystem. Glob. Change Biol. 2010, 16, 2676–2687. [Google Scholar] [CrossRef]
- Yang, H.; Wu, M.; Liu, W.; Zhang, Z.; Zhang, N.; Wan, S. Community structure and composition in response to climate change in a temperate steppe. Glob. Change Biol. 2011, 17, 452–465. [Google Scholar] [CrossRef]
- Garroway, C.J.; Bowman, J.; Cascaden, T.J.; Holloway, G.L.; Mahan, C.G.; Malcolm, J.R.; Steele, M.A.; Turner, J.; Wilson, P.J. Climate change induced hybridization in flying squirrels. Glob. Change Biol. 2010, 16, 113–121. [Google Scholar] [CrossRef]
- Wellenreuther, M.; Tynkkynen, K.; Svensson, E.I. Simulating range expansion: Male species recognition and loss of premating isolation in damselflies. Evolution 2010, 64, 242–252. [Google Scholar] [CrossRef]
- Visser, M.E.; Both, C. Shifts in phenology due to global climate change: The need for a yardstick. Proc. R. Soc. B Biol. Sci. 2005, 272, 2561–2569. [Google Scholar] [CrossRef]
- Williams, J.W.; Jackson, S.T. Novel climates, no-analog communities, and ecological surprises. Front. Ecol. Environ. 2007, 5, 475–482. [Google Scholar] [CrossRef]
- Alevi, K.C.C.; Oliveira, J.; Rocha, D.S.; Galvão, C. Trends in Taxonomy of Chagas Disease Vectors (Hemiptera, Reduviidae, Triatominae): From Linnaean to Integrative Taxonomy. Pathogens 2021, 10, 1627. [Google Scholar] [CrossRef]
- Correia, J.P.S.O.; Gil-Santana, H.R.; Dale, C.; Galvão, C. Triatoma guazu Lent and Wygodzinsky Is a Junior Synonym of Triatoma williami Galvão, Souza and Lima. Insects 2022, 13, 591. [Google Scholar] [CrossRef] [PubMed]
- Gil-Santana, H.R.; Chavez, T.; Pita, S.; Panzera, F.; Galvão, C. Panstrongylus noireaui, a remarkable new species of Triatominae (Hemiptera, Reduviidae) from Bolivia. ZooKeys 2022, 1104, 203–225. [Google Scholar] [CrossRef] [PubMed]
- Téllez-Rendón, J.; Esteban, L.; Rengifo-Correa, L.; Díaz-Albiter, H.; Huerta, H.; Dale, C. Triatoma yelapensis sp. nov. (Hemiptera: Reduviidae) from Mexico, with a Key of Triatoma Species Recorded in Mexico. Insects 2023, 1012, 14–331. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Galvão, C.; Cai, W. Rhodnius micki, a new species of Triatominae (Hemiptera, Reduviidae) from Bolivia. ZooKeys 2021, 1012, 71–93. [Google Scholar] [CrossRef] [PubMed]
- Justi, S.A.; Galvão, C.; Schrago, C.G. Geological changes of the Americas and their influence on the diversification of the Neotropical kissing bugs (Hemiptera: Reduviidae: Triatominae). PLoS Negl. Trop. Dis. 2016, 10, e0004527. [Google Scholar] [CrossRef] [Green Version]
- Hernández, C.; Alvarado, M.; Salgado-Roa, F.C.; Ballesteros, N.; Rueda-M, N.; Oliveira, J.; Alevi, K.C.C.; da Rosa, J.A.; Urbano, P.; Salazar, C.; et al. Phylogenetic relationships and evolutionary patterns of the genus Psammolestes Bergroth, 1911 (Hemiptera: Reduviidae: Triatominae). BMC Ecol. Evol. 2022, 22, 30. [Google Scholar] [CrossRef]
- Justi, S.A.; Galvão, C. The Evolutionary Origin of Diversity in Chagas Disease Vectors. Trends Parasitol. 2017, 33, 42–52. [Google Scholar] [CrossRef] [Green Version]
- Lent, H.; Wygodzynsky, P. Revision of the Triatominae (Hemiptera—Reduviidae) and their significance as vectors of Chagas’ disease. Bull. Am. Mus. Nat. Hist. 1979, 163, 123–520. [Google Scholar]
- Hernández, C.; Rosa, J.A.; Vallejo, G.A.; Guhl, F.; Ramirez, J.D. Taxonomy, Evolution and Biogeography of the Rhodniini Tribe (Hemiptera: Reduviidae). Diversity 2020, 12, 97. [Google Scholar] [CrossRef] [Green Version]
- Abad-Franch, F.; Pavan, M.G.; Jaramillo, N.; Palomeque, F.S.; Dale, C.; Chaverra, D.; Monteiro, F.A. Rhodnius barretti, a new species of Triatominae (Hemiptera: Reduviidae) from western Amazonia. Mem. Inst. Oswaldo Cruz 2013, 108, 92–99. [Google Scholar] [CrossRef]
- Carvalheiro, J.R.; Barretto, M.P. Estudos sobre reservatórios e vectores silvestres do Trypanosoma cruzi. LX—Tentativas de cruzamento de Rhodnius prolixus Stal, 1859 com Rhodnius neglectus Lent, 1954 (Hemiptera, Reduviidae). Rev. Soc. Bras. Med. Trop. 1976, 18, 17–23. [Google Scholar]
- Galíndez, G.L.; Barazarte, R.; Márquez, J.; Oviedo, M.; Márquez, Y.; Morón, L.; Carcavallo, R. Relaciones reproductivas entre Rhodnius prolixus Stal y Rhodnius robustus Larrousse (Hemiptera, Reduviidae, Triatominae) bajo condiciones de laboratorio. Entomol. Y Vectores 1994, 1, 3–13. [Google Scholar]
- Díaz, S.; Panzera, F.; Jaramillo-O, N.; Pérez, R.; Fernández, R.; Vallejo, G.; Saldaña, A.; Calzada, J.E.; Triana, O.; Gómez-Palacio, A. Genetic, Cytogenetic and Morphological Trends in the Evolution of the Rhodnius (Triatominae: Rhodniini) Trans-Andean Group. PLoS ONE 2014, 9, e87493. [Google Scholar] [CrossRef] [Green Version]
- Ravazi, A.; Oliveira, J.; Campos, F.F.; Madeira, F.F.; Reis, Y.V.; Oliveira, A.B.B.; Azeredo-Oliveira, M.T.V.; Rosa, J.A.; Galvão, C.; Alevi, K.C.C. Trends in evolution of the Rhodniini tribe (Hemiptera, Triatominae): Experimental crosses between Psammolestes tertius Lent & Jurberg, 1965 and P. coreodes Bergroth, 1911 and analysis of the reproductive isolating mechanisms. Parasites Vectors 2021, 14, 350. [Google Scholar]
- Mayr, E. Animal Species and Evolution; Harvard University Press: Cambridge, MA, USA, 1963. [Google Scholar]
- Rosa, J.A.; Barata, J.M.S.; Barelli, N.; Santos, J.L.F.; Belda Neto, F.M. Sexual distinction between 5th instar nymphs of six species (Hemiptera: Reduviidae). Mem. Inst. Oswaldo Cruz 1992, 87, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Ibarra, J.Á.; Grant-Guillén, Y.; Delgadillo-Aceves, I.N.; Zumaya-Estrada, F.A.; Rocha-Chávez, G.; Salazar-Schettino, P.M.; Alejandre-Aguilar, R.; Villalobos, G.; Nogueda-Torres, B. Biological and genetic aspects of crosses between phylogenetically close species of Mexican Triatomines (Hemiptera: Reduviidae). J. Med. Entomol. 2011, 48, 705–707. [Google Scholar] [CrossRef] [Green Version]
- Olaia, N.; Alevi, K.C.C.; Oliveira, J.; Cacini, G.L.; Souza, E.D.S.; Pinotti, H.; Silva, L.A.; Rosa, J.A. Biology of Chagas disease vectors: Biological cycle and emergence rates of Rhodnius marabaensis Souza et al., 2016 (Hemiptera, Reduviidae, Triatominae) under laboratory conditions. Parasitol. Res. 2021, 120, 2939–2945. [Google Scholar] [CrossRef]
- Geographic Information System. Available online: http://qgis.org (accessed on 18 January 2023).
- Vogels, C.B.F.; Fros, J.J.; Göertz, G.P.; Piljman, G.P.; Koenraadt, C.J.M. Vector competence of northern European Culex pipiens biotypes and hybrids for West Nile virus is differentially affected by temperature. Parasites Vectors 2016, 9, 393. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Ibarra, J.A.; Nogueda-Torres, B.; Salazar-Schettino, P.M.; Cabrera-Bravo, M.; Vences-Blanco, M.O.; Rocha-Chávez, G. Transmission Capacity of Trypanosoma cruzi (Trypanosomatida: Trypanosomatidae) by Three Subspecies of Meccus phyllosomus (Heteroptera: Reduviidae) and Their Hybrids. J. Med. Entomol. 2016, 53, 928–934. [Google Scholar] [CrossRef]
- Martínez-Ibarra, J.A.; Nogueda-Torres, B.; García-Lin, J.C.; Arroyoreys, D.; Salazar-Montaño, L.F.; Hernández-Navarro, J.A.; Díaz-Sánchez, C.G.; Toro-Arreola, E.S.D.; Rocha-Chávez, G. Importance of Hybrids of Meccus phyllosomus mazzottii, and M. p. pallidipennis, and M. p. phyllosomus to the Transmission of Trypanosoma cruzi in Mexico. Jpn. J. Infect. Dis. 2016, 69, 202–206. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Ibarra, J.A.; Nogueda-Torres, B.; Salazar-Montaño, L.F.; García-Lino, J.C.; Arroyo-Reyes, D.; Hernández-Navarro, J.A. Comparison of biological fitness in crosses between subspecies of Meccus phyllosomus (Hemiptera: Reduviidae: Triatominae) in southern Mexico. Insect. Sci. 2017, 24, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Meraz-Medina, T.; Nogueda-Torres, B.; Ceballos-Rodríguez, R.S.; Godínez-Aceves, K.C.; Martínez-Ibarra, J.A. Enhancing fitness in offspring of crosses between two triatomine species. J. Vector Ecol. 2019, 44, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Reis, Y.V.; Alevi, K.C.C. Hybridization in Phlebotominae (Diptera: Psychodidae): A mini-review. Infect. Genet. Evol. 2020, 85, 104593. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ibarra, J.A.; Cárdenas-Sosa, M.A.; Montañez-Valdez, O.D.; Michel-Parra, J.G.; Nogueda-Torres, B. Biological parameters and estimation of the vectorial capacity of two subspecies of Triatoma protracta (Uhler) and their laboratory hybrids in Mexico. J. Vector Ecol. 2021, 46, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, H.J.; Segovia, M.; Londoño, J.C.; Ortegoza, J.; Rodríguez, M.; Martínez, C.E. Panstrongylus geniculatus and four other species of triatomine bug involved in the Trypanosoma cruzi enzootic cycle: High risk factors for Chagas’ disease transmission in the Metropolitan District of Caracas, Venezuela. Parasites Vectors 2014, 7, 602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, A.; Roque, A.L.R. Domestic and wild mammalian reservoirs. In American Trypanosomiasis: Chagas Disease One Hundred Years of Research, 1st ed.; Telleria, J., Tibyarenc, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2010; pp. 249–276. [Google Scholar]
- Peterson, J.K.; Graham, A.L.; Dobson, A.P.; Chávez, O.T. Rhodnius prolixus Life History Outcomes Differ when Infected with Different Trypanosoma cruzi I Strains. Am. J. Trop. Med. Hyg. 2015, 93, 564–572. [Google Scholar] [CrossRef]
- Zeledón, R.; Marín, F.; Calvo, N.; Lugo, E.; Valle, S. Distribution and ecological aspects of Rhodnius pallescens in Costa Rica and Nicaragua and their epidemiological implications. Mem. Inst. Oswaldo. Cruz 2006, 101, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Grijalva, M.J.; Villacis, A.G.; Ocana-Mayorga, S.; Yumiseva, C.A.; Baus, E.G. Limitations of selective deltamethrin application for triatomine control in central coastal Ecuador. Parasites Vectors 2011, 4, 20. [Google Scholar] [CrossRef] [Green Version]
- Grijalva, M.J.; Palomeque-Rodriguez, F.S.; Costales, J.A.; Davila, S.; Arcos-Teran, L. High household infestation rates by synanthropic vectors of Chagas disease in southern Ecuador. J. Med. Entomol. 2005, 42, 68–74. [Google Scholar] [CrossRef]
- Silveira, A.C.; Rezende, D.F. Epidemiologia e controle da transmissăo vetorial da doença de Chagas no Brasil. Rev. Soc. Bras. Med. Trop. 1994, 27, 11–22. [Google Scholar]
- Lima, M.M.; Coutinho, C.F.; Gomes, T.F.; Oliveira, T.G.; Duarte, R.; Borges-Pereira, J.; Bóia, M.N.; Sarquis, O. Risk presented by Copernicia prunifera palm trees in the Rhodnius nasutus distribution in a Chagas disease-endemic area of the Brazilian northeast. Am. J. Trop. Med. Hyg. 2008, 79, 750–754. [Google Scholar] [CrossRef] [Green Version]
- López, A.G.; Cardozo, M.; Oscherov, E.B.; Crocco, L.B. Dynamics of feeding and defecation behavior of Triatoma infestans hybrids. Parasitol. Res. 2020, 119, 2775–2781. [Google Scholar] [CrossRef]
- Vicente, R.D.; Madeira, F.F.; Borsatto, K.C.; Garcia, A.C.C.; Cristal, D.C.; Delgado, L.M.G.; Bittinelli, I.S.; Mello, D.V.; Reis, Y.V.; Ravazi, A.; et al. Morphological, Cytological and Molecular Studies and Feeding and Defecation Pattern of Hybrids from Experimental Crosses between Triatoma sordida and T. rosai (Hemiptera, Triatominae). Pathogens 2022, 11, 1302. [Google Scholar] [CrossRef]
- Galvão, C.; Gurgel-Gonçalves, R. Vetores conhecidos no Brasil. In Vetores da Doença de Chagas no Brasil, 1st ed.; Galvão, C., Ed.; SciELO: Curitiba, Brazil, 2014; pp. 88–170. [Google Scholar]
- Anderson, E. Introgressive Hybridization, 1st ed.; Wiley & Sons: New York, NY, USA, 1949. [Google Scholar]
- Anderson, E.; Hubricht, L. Hybridization in Tradescantia. III. The evidence for introgressive hybridization. Am. J. Bot. 1938, 25, 396–402. [Google Scholar] [CrossRef]
- Fontaine, M.C.; Pease, J.B.; Steele, A.; Waterhouse, R.M.; Neafsey, D.E.; Sharakhov, I.V.; Jiang, X.; Hall, A.B.; Catruccia, F.; Kakani, E.; et al. Extensive introgression in a malaria vector species complex revealed by phylogenomics. Science 2014, 347, 1258524. [Google Scholar] [CrossRef] [Green Version]
- Vicente, J.L.; Clarkson, C.S.; Caputo, B.; Gomes, B.; Pombi, M.; Sousa, C.A.; Antao, T.; Dinis, J.; Bottà, G.; Mancini, E.; et al. Massive introgression drives species radiation at the range limit of Anopheles gambiae. Sci. Rep. 2017, 7, 46451. [Google Scholar] [CrossRef] [Green Version]
- Souza, R.C.M.; Gorla, D.E.; Chame, M.; Jaramillo, N.; Monroy, C.; Diotaiuti, L. Chagas disease in the context of the 2030 agenda: Global warming and vectors. Mem. Inst. Oswaldo Cruz 2022, 117, e200479. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Study Area | Projection Periods | Key Findings | Reference |
|---|---|---|---|---|
| Triatoma sanguisuga, T. barberi, T. peninsularis, T. protracta, T. rubida, T. brailovskyi, T. gerstaeckeri, T. longipennis, T. mazzottii, T. mexicana, T. pallidipennis, T. phyllosoma, T. picturata, T. recurva, T. dimidiata, Eratyrus cuspidatus, Paratriatoma hirsuta and P. lecticularia | North America | 2050 2070 | 1. Significant alterations in risk of vector-borne T. cruzi transmission; 2. Impacts of climate change on T. cruzi vector species and their geographical locations; 3. Need for effective vector surveillance to aid in CD prevention and control. | [28] |
| T. gerstaeckeri and T. sanguisuga | North America | 2050 | 1. A potential change in the distribution of both species is predicted due to climate change, indicating an increase in T. cruzi transmission risk by the year 2050; 2. Predicted shift of CD risk due to climate change. | [29] |
| P. lecticularia, T. protracta and T. sanguisuga | United States | 2030 | 1. The increase in temperature is expected to promote the expansion of the vector range by the year 2030; 2. Potential for CD to emerge in the country; 3. It is believed that interdisciplinary collaboration is necessary to increase the accuracy of future disease risk predictions. | [30] |
| Panstrongylus geniculatus, E. mucronatus, Rhodnius prolixus, R. robustus and T. maculata | Venezuela | 2020 2060 2080 | 1. Global climate change is predicted to slightly decrease the overall vulnerability of the population to T. cruzi vector species; 2. It is believed that fewer citizens will be exposed to T. cruzi vectors over the next 50–70 years; 3. Predictions can enhance the ability to prevent and control CD transmission in Venezuela in the future. | [25] |
| R. prolixus and T. infestans | VenezuelaArgentina | 2050 | 1. Forecasts reveal the increased expansion of R. prolixus to new areas and a decrease in the current geographical range of T. infestans; 2. Overall study conclusions reflect a lower incidence of CD infections by 2050. | [26] |
| T. brasiliensis brasiliensis, T. b. macromelasoma, T. juazeirensis, T. sherlocki and T. melanica | Brazil | 2020 2050 | 1. Projection of little change in species’ distributions under future climate change scenarios; 2. T. b. brasiliensis has the greatest distributional potential to colonize new areas; 3. Conclusions may help guide proactive surveillance and control activities to reduce the risk of future CD transmission. | [31] |
| T. infestans | Chile | 2070 | 1. The distribution under two global climate change scenarios showed low variation, with a minimal reduction tendency in suitable areas; 2. Climate change appears to play a major role in the reemergence of CD and of the vector in the country; 3. The impact of temperature and precipitation on the distribution of T. infestans indicates the need for aggressive vector control efforts in Chile. | [32] |
| M.epraia spinolai and M. gajardoi | Chile | 2070 | 1. Under future climate conditions, these species could modify their potential geographical range; 2. The suitable areas for both species may be greater than currently known, generating new challenges in terms of vector control and prevention; 3. Preventive measures to avoid accidental human vectorial transmission by wild vectors of T. cruzi become critical, considering the uncertainty of the future suitable areas projected in this study. | [27] |
| P. geniculatus, P. megistus, R. brethesi, R. ecuadoriensis, R. prolixus, T. b. brasiliensis, T. dimidiata, T. infestans, T. maculata, T. rubrofasciata and T. sordida | Different countries from five continents: Africa, Asia, Europe, Oceania and America. | 2030 | 1. Global species distribution modeling revealed several regions with current suitable climatic conditions; 2. R. brethesi, R. ecuadoriensis and T. maculata are limited to one or a few areas with mostly tropical climate; 3. T. b. brasiliensis, P. geniculatus, P. megistus, R. prolixus, T. dimidiata and T. rubrofasciata find suitable climate conditions in a broad range of tropical and subtropical regions; 4. T. sordida and T. infestans possess a broad potential range in temperate regions; 5. It may be beneficial to establish national and international vector surveillance programs to monitor the spread of vectors and to register CD as a reportable disease. | [33] |
| Species | GBIF Dataset |
|---|---|
| Psammolestes coreodes Bergroth, 1911 | https://doi.org/10.15468/dl.vhbk9r |
| Psammolestes tertius Lent & Jurberg, 1966 | https://doi.org/10.15468/dl.d78jfs |
| Rhodnius brethesi Matta, 1919 | https://doi.org/10.15468/dl.hctjyp |
| Rhodnius colombiensis Mejia, Galvão & Jurberg, 1999 | https://doi.org/10.15468/dl.muvwy2 |
| Rhodnius ecuadoriensis Lent & León, 1958 | https://doi.org/10.15468/dl.bbc7ft |
| Rhodnius marabaensis Souza et al., 2016 | https://doi.org/10.15468/dl.v32gdm |
| Rhodnius milesi Carcavallo, Rocha, Galvão & Jurberg, 2001 | https://doi.org/10.15468/dl.ga2kks |
| Rhodnius montenegrensis da Rosa et al., 2012 | https://doi.org/10.15468/dl.7gjaws |
| Rhodnius nasutus Stål, 1859 | https://doi.org/10.15468/dl.e8vm73 |
| Rhodnius neglectus Lent, 1954 | https://doi.org/10.15468/dl.9w7xdu |
| Rhodnius neivai Lent, 1953 | https://doi.org/10.15468/dl.zghcuf |
| Rhodnius pictipes Stål, 1872 | https://doi.org/10.15468/dl.tknw8g |
| Rhodnius prolixus Stål, 1859 | https://doi.org/10.15468/dl.ggavgu |
| Rhodnius robustus Larrousse, 1927 | https://doi.org/10.15468/dl.bcps8k |
| Rhodniini | Crossing Experiments | Eggs | Hybrids | Hatch Rate |
|---|---|---|---|---|
| Psammolestes | P. coreodes ♀ × P. tertius ♂ | 54 * | 23 * | 43% * |
| P. tertius ♀ × P. coreodes ♂ | 117 * | 30 * | 26% * | |
| Rhodnius | R. marabaensis ♀ × R. montenegrensis ♂ | 214 | 128 | 60% |
| R. montenegrensis ♀ × R. marabaensis ♂ | 436 | 36 | 08% | |
| R. brethesi ♀ × R. pictipes ♂ | 268 | 16 | 06% | |
| R. pictipes ♀ × R. brethesi ♂ | 162 | 00 | 00% | |
| R. colombiensis ♀ × R. ecuadoriensis ♂ | 170 | 45 | 06% | |
| R. ecuadoriensis ♀ × R. colombiensis ♂ | 42 | 00 | 00% | |
| R. montenegrensis ♀ × robustus ♂ | 565 | 331 | 59% | |
| R. robustus ♀ × R. montenegrensis ♂ | 611 | 484 | 57% | |
| R. neivai ♀ × R. prolixus ♂ | 70 | 22 | 31% | |
| R. prolixus ♀ × R. neivai ♂ | 368 | 00 | 00% | |
| R. prolixus ♀ × R. nasutus ♂ | 318 | 71 | 22% | |
| R. nasutus ♀ × R. prolixus ♂ | 316 | 14 | 04% | |
| R. milesi ♀ × R. neglectus ♂ | 357 | 81 | 23% | |
| R. neglectus ♀ × R. milesi ♂ | 366 | 314 | 86% | |
| R. robustus ♀ × R. prolixus ♂ | 622 | 447 | 72% | |
| R. prolixus ♀ × R. robustus ♂ | 437 | 221 | 51% | |
| R. taquarussuensis (syn. R. neglectus) ♀ × R. neglectus ♂ | 540 | 496 | 92% | |
| R. neglects ♀ × R. taquarussuensis (syn. R. neglectus) ♂ | 737 | 636 | 85% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ravazi, A.; Oliveira, J.d.; Madeira, F.F.; Nunes, G.M.; Reis, Y.V.d.; Oliveira, A.B.B.d.; Azevedo, L.M.S.; Galvão, C.; Azeredo-Oliveira, M.T.V.d.; Rosa, J.A.d.; et al. Climate and Environmental Changes and Their Potential Effects on the Dynamics of Chagas Disease: Hybridization in Rhodniini (Hemiptera, Triatominae). Insects 2023, 14, 378. https://doi.org/10.3390/insects14040378
Ravazi A, Oliveira Jd, Madeira FF, Nunes GM, Reis YVd, Oliveira ABBd, Azevedo LMS, Galvão C, Azeredo-Oliveira MTVd, Rosa JAd, et al. Climate and Environmental Changes and Their Potential Effects on the Dynamics of Chagas Disease: Hybridization in Rhodniini (Hemiptera, Triatominae). Insects. 2023; 14(4):378. https://doi.org/10.3390/insects14040378
Chicago/Turabian StyleRavazi, Amanda, Jader de Oliveira, Fernanda Fernandez Madeira, Giovana Menezes Nunes, Yago Visinho dos Reis, Ana Beatriz Bortolozo de Oliveira, Luísa Martins Sensato Azevedo, Cleber Galvão, Maria Tercília Vilela de Azeredo-Oliveira, João Aristeu da Rosa, and et al. 2023. "Climate and Environmental Changes and Their Potential Effects on the Dynamics of Chagas Disease: Hybridization in Rhodniini (Hemiptera, Triatominae)" Insects 14, no. 4: 378. https://doi.org/10.3390/insects14040378