
Population Genetic Diversity of Two Blue Oat Mite Species on Triticum Hosts in China

Key Laboratory of Agricultural Biosafety and Green Production of Upper Yangtze River, Key Laboratory of Entomology and Pest Control Engineering, College of Plant Protection, Southwest University, Chongqing 400716, China
*
Authors to whom correspondence should be addressed.
Insects 2023, 14(4), 377; https://doi.org/10.3390/insects14040377
Submission received: 1 March 2023
/
Revised: 5 April 2023
/
Accepted: 10 April 2023
/
Published: 12 April 2023
(This article belongs to the Collection Integrated Pest Management of Crop)
Abstract
:Simple Summary
Blue oat mite species mainly include Penthaleus major, P. falcatus, P. tectus, and P. minor. Among them, P. major, P. falcatus and P. tectus are important pest mites on gramineous crops, which often occur in low-temperature and high-humidity environments. These three mites are difficult to distinguish in the field due to their small size and similar shape. Furthermore, they exhibit different resistance to chemical pesticides, making it challenging to control their outbreak. Worryingly, we only saw reports of frequent occurrence of P. major in winter wheat regions in China, and no other species of blue oat mites were found. Understanding the distribution pattern, genetic diversity, and genetic differentiation among populations of blue oat mite species in winter wheat regions of China is essential for developing effective control programs. In this study, we evaluated the distribution of blue oat mites in the major wheat regions in China and assessed the level of genetic diversity and genetic structure of blue oat mites from winter wheat regions of China using mitochondrial cytochrome c oxidase I (COI) sequences.
Abstract
Blue oat mite species, including Penthaleus major and P. tectus, are pests widely distributed across China that cause damage to winter wheat. This study evaluated the genetic diversity of P. major and P. tectus on Triticum hosts collected from 23 geographic locations based on mitochondrial cytochrome c oxidase subunit I (COI) sequences. We identified nine haplotypes in 438 P. major individuals from 21 geographic locations and five haplotypes in 139 P. tectus individuals from 11 geographic locations. Meanwhile, P. major exhibits high values of haplotype diversity (Hd) and nucleotide diversity (Pi) (Hd = 0.534 > 0.5 and Pi = 0.012 > 0.005), representing a large stable population with a long evolutionary history. P. tectus shows low values of Hd and Pi (Hd = 0.112 < 0.5 and Pi = 0 < 0.005), which suggest recent founder events. Moreover, demographic analysis suggested that P. major and P. tectus have not undergone a recent population expansion. The lowest genetic variation was observed in Xiangzhou (XZ-HB), Zaoyang (ZY-HB), Siyang (SY-JS), and Rongxian (RX-SC), with only one species and one haplotype identified in over 30 individuals. Robust genetic differentiation was found in P. major compared to P. tectus, which provides a theoretical basis for the widespread distribution of P. major in China.
1. Introduction
Blue oat mites (Penthaleus spp.) are significant agricultural pests in temperate regions around the world, attacking various pastures, vegetables, and crop plants [1,2,3,4,5,6]. These mites cause severe damage to plants by directly penetrating the plants’ epidermal cells and removing the cellular contents [7,8]. Among the three blue oat mites, P. major was first described by Dugés in France. In addition, Womersley gives this mite its current name, P. major, by recognizing the synonymy among previously named specimens [9]. After that, two other asexual mites were found, including P. falcatus and P. tectus [8,10,11,12].
Previous research has shown that P. major, P. falcatus, and P. tectus were diploid and reproduced by thelytokous parthenogenesis [10,11]. A high-level clonal variation was found within populations of P. major from southeastern Australia [11,13,14], while P. falcatus and P. tectus have fewer allozyme clones, six and four, respectively [11]. P. major has been studied as a model species to investigate how genetic diversity is maintained in asexual organisms [15].
The extent of genetic variability of a population determines its ability to adapt to the environment [16,17]. Numerous correlations between genetic variation and fitness traits, such as growth, survival, and resistance have been reported in the literature [18,19,20]. Recently, the genetic diversity of different geographical populations of pests has been widely studied based on mitochondrial cytochrome c oxidase subunit I (COI) sequences, including Aedes albopictus [21] and Culicoides mahasarakhamensee [22].
In China, two pest mites are widely reported on wheat, including P. major Dugés (Acari: Penthaleidae) and Petrobia latens Müller (Acari: Tetranychidae). P. major is mainly distributed in 29–37° N, including southern Hebei, Shanxi, Shandong, Henan, Anhui, Jiangsu, Zhejiang, and Sichuan. P. latens is mainly distributed in 34–43° N, including Liaoning, Beijing, Gansu, Qinghai, Xinjiang, Xizang, Hebei, Henan, Shandong, Shanxi, Shaanxi, and Anhui [23]. Most domestic research has primarily focused on the harm of mites on winter wheat and a series of control methods [24]. Foreign studies show that there are at least three different species in the same genus for blue oat mites, and there is a phenomenon of simultaneous damage [4,25]. Furthermore, different types of pest mites have significantly different resistance to chemical agents [11,26].
Winter wheat is widely planted in China and is always damaged by pest mites. However, the specific species of mites on wheat are still unclear in China. Thus, rigorous research is needed to learn about the distribution pattern and genetic diversity of mites in different parts of the country. Therefore, in this study, we aimed to investigate the damage of blue oat mites in three major winter wheat regions, including Southwestern Winter Wheat Zone (SW), Yellow and Huai River Facultative Winter Wheat Zone (YH), and Middle and Low Yangtze Valley Winter Wheat Zone (YV). Furthermore, we analyzed the genetic diversity of blue oat mites by mitochondrial COI sequences. This study aimed to examine the genetic variation and structure of blue oat mites on Triticum hosts from different locations across China using mitochondrial DNA. Additionally, phylogenetic relationships of haplotypes with published haplotypes of COI worldwide were investigated.
2. Materials and Methods
2.1. Investigation on Wheat Mites
This study conducted a field survey of important pest mites on winter wheat in major regions across China in December 2018 and February–March 2019. The survey covered 10 provinces and one municipality, namely, Sichuan, Guizhou, Yunnan, Jiangsu, Anhui, Hubei, Henan, Shanxi, Shandong, Shaanxi, Hebei, and Chongqing. Specifically, 35 survey sites were included, namely Rongxian, Mianzhu, Santai, Nanchong, Tongnan, Liangping, Rongchang, Qianxinan, Liupanshui, Baoshan, Lijiang, Suining, Donghai, Xuzhou, Jiawang, Siyang, Funnan, Bozhou, Changfeng, Xiaoxian, Dangyang, Xiangzhou, Zhongxiang, Zaoyang, Dancheng, Shanzhou, Tanghe, Luohe, Linyi, Ruicheng, Gaotang, Weinan, Xingtai, Handan, and Yuncheng.
2.2. Specimen Collection
A total of 577 adult mites were collected from Triticum hosts at 23 locations across China between December 2018 and February–March 2019. According to previous academic reports, the 23 locations were divided into three natural geographic groups, respectively, Southwestern China (SW), Middle and Lower Yangtze Valleys (YV), and Yellow and Huai Valleys (YH). The mites were stored in 95% ethanol or at −20 °C until DNA extraction. The sampling locations were distributed in seven provinces, including Sichuan province, Jiangsu province, Anhui province, Henan province, Shandong province, Hubei province, Shanxi province, and one municipality, Chongqing.
2.3. DNA Extraction and Mitochondrial COI Gene Amplification
DNA was extracted from individual female adult mites using 30 μL STE buffer in a 1.5 mL centrifuge tube. To the extract, 2 μL proteinase K (20 mg/mL) was added, and the mixture was incubated at 37 °C for 30 min, followed by heating to 95 °C for 5 min [27,28]. COI fragments were amplified using COI universal primer pair, LCO1490: 5′-GGTCAACAAATCATAAAGATATTGG-3′ and HCO2198: 5′-TAAACTTCAGGGTGACCAAAAAATCA-3′ [29]. Polymerase chain reaction (PCR) mixture (25 μL) contained 2.0 μL supernatant of the lysate, 1.0 μL of each primer (10 μM), 2.0 μL of dNTPs (2.5 mM), 2.0 μL of Mg2+ solution (25 mM), 2.5 μL of 10 × PCR buffer (Mg2+ free), 14.5 μL nuclease-free water, and 0.25 μL of rTaq polymerase (TaKaRa). The PCR reactions procedure was as follows: initial denaturing for 5 min at 95 °C; 35 cycles of 95 °C for 60 s, 57 °C for 60 s, and 72 °C for 90 s; and a final extension at 72 °C for 5 min. The size and the quality of PCR products were examined by 1.0% TAE agarose gel electrophoresis and sequenced in both directions by Sangon (Shanghai, China).
2.4. Data Analysis
The assembling and alignment of COI sequences were completed in DNASTAR LASERGENE 7.1.0 and MEGA 7.0, respectively [30,31,32]. Genetic diversity parameters were determined in DnaSP 5.10.01 [27], including the number of polymorphic sites (S), the total number of mutations (η), the number of haplotypes (H), haplotype diversity (Hd), nucleotide diversity (Pi), and the average number of nucleotide differences (K). A total of 12 haplotypes reported from Canada and Poland were used to analyze the phylogenetic relationships. Phylogenetic analyses were performed in MEGA 7.0 using the Neighbor-Joining method, while haplotype networks of P. major and P. tectus haplotypes were constructed in TCS 1.21 [33]. Pairwise FST and gene flow (Nm) between each pair of the sampled locations and pairwise FST and gene flow (Nm) among three geographic regions were calculated in ARLEQUIN 3.5 [34]. The mismatch distribution analysis was performed to detect historical population expansion events in P. major and P. tectus populations in DnaSP 5.10.01. A multimodal pattern implies that populations are at demographic equilibrium, whereas a unimodal pattern shows that populations are experiencing rapid demographic growth [35,36].
3. Results
3.1. Distribution Pattern of Wheat Pest Mites
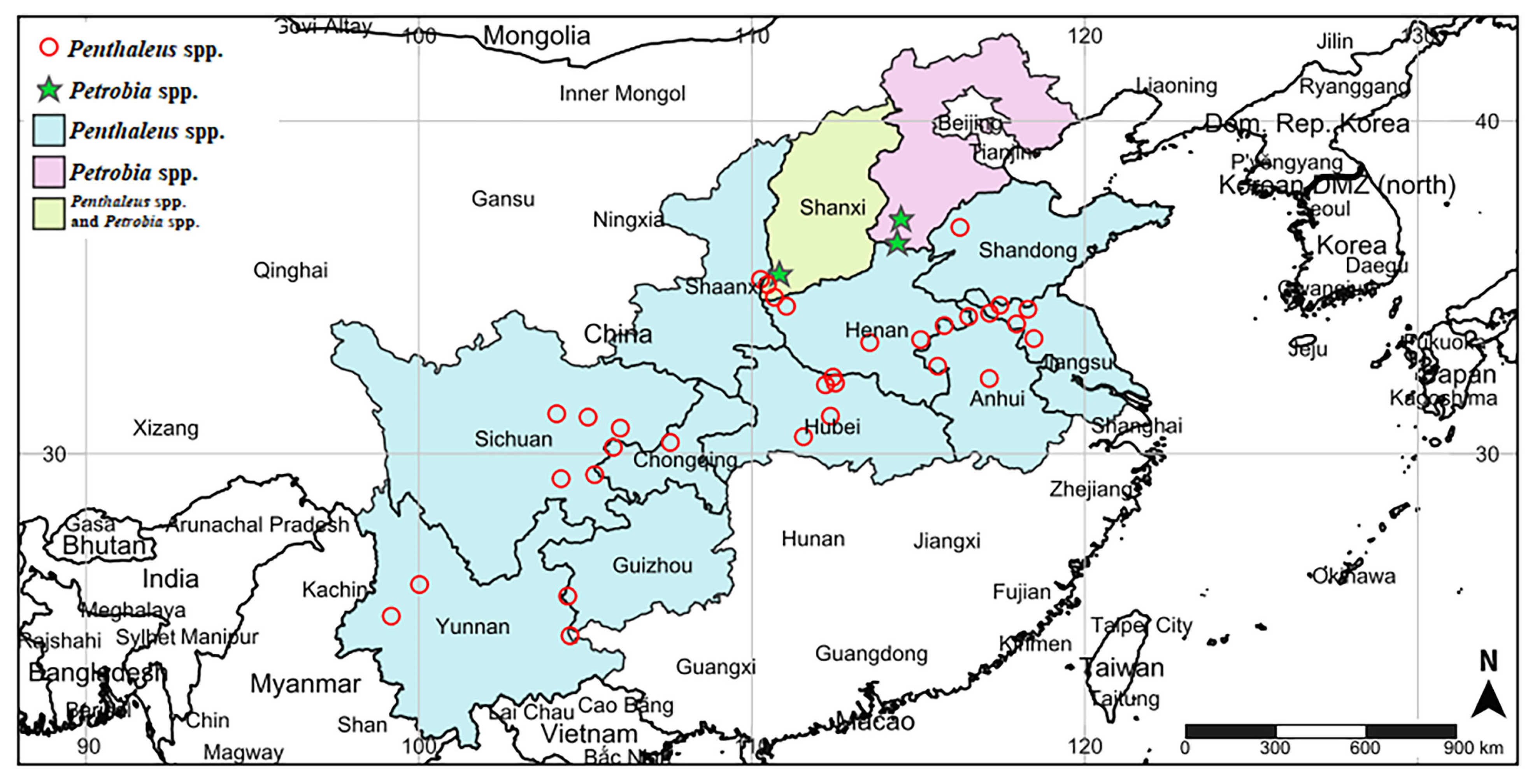
Two genera of mites were found in the main winter wheat regions of China during this survey, including Penthaleus spp. and Petrobia spp. (Figure 1). The study found that Penthaleus spp. were the main pest mites in winter wheat regions and were distributed in 11 provinces and a municipality, including Yunnan, Sichuan, Guizhou, Shaanxi, Hubei, Shanxi, Henan, Anhui, Shandong, Jiangsu, and Chongqing. The range of Petrobia spp. was small, primarily in Shanxi and Hebei. Moreover, the map revealed that both Penthaleus spp. and Petrobia spp. occurred in Shanxi Province. Penthaleus spp. was the most widely distributed; however, it was not found in Hebei. Among the 35 survey sites, Penthaleus spp. was found in 32 sites, accounting for up to 91.43%, while Petrobia spp. was only found in three sites. The population distribution range and number were significantly lower than Penthaleus spp.
3.2. Population Distribution of Blue Oat Mites
In this study, the dominant population of blue oat mite species on winter wheat in China was identified (Figure 2; Table 1). Two species of blue oat mites were found in 577 samples, including P. major and P. tectus. Among them, P. major accounted for 75.91%, more than P. tectus. P. major and P. tectus were found throughout three natural geographic groups, including Southwestern China (SW), the Middle and Lower Yangtze Valleys (YV), and the Yellow and Huai Valleys (YH). Of the 23 sites surveyed, P. major was found in 21 survey sites, with the exceptions being Rongxian (RX-SC) and Ruicheng (RC-SX). The distribution of P. tectus is similar to that of P. major, but it is not always found in the same sites. At nine sites, P. major and P. tectus co-occurred, including Santai (ST-SC), Nanchong (NC-SC), Dangyang (DY-HB), Xuzhou (XZ-JS), Donghai (DH-JS), Bozhou (BZ-AH), Changfeng (CF-AH), Shanzhou (SZ-HN), and Tanghe (TH-HN). Additionally, only P. tectus was found at two survey sites, including Rongxian (RX-SC) and Ruicheng (RC-SX).
3.3. Haplotype Diversity, Nucleotide Diversity, and Haplotype distribution
The COI gene fragment, which was amplified from 577 individuals in 23 different locations across three central winter wheat regions, ranged from 657 to 658 bp (Table 2). Of the 438 individuals of P. major, nine haplotypes (MH1-MH9) were identified with 22 polymorphic nucleotide sites. Nineteen of these sites were parsimony-informative, and three were singleton sites. The A/T content was significantly higher (70.30%) than the C/G content (29.70%). Meanwhile, 139 P. tectus individuals generated five haplotypes (TH1-TH5) with four polymorphic nucleotide sites. Four of these sites were parsimony-informative sites and none were singleton sites. The A/T content was significantly higher (69.90%) than the C/G content (30.10%).
The Hd of P. major ranged from 0.000 in the seven populations (NC-SC, XZ-JS, SY-JS, BZ-AH, XZ-HB, ZY-HB, TH-HN) to 0.708 CF-AH population (Table 3). Haplotype diversity of P. major was identified in 12 of the 21 locations. For the other nine locations, only one haplotype was found. The Hd of P. tectus ranged from 0.000 in the six populations (RX-SC, ST-SC, NC-SC, XZ-JS, SZ-HN, TH-HN) to 0.467 CF-AH population. Additionally, haplotype diversity of P. tectus was only observed in three of the 11 locations. For the other eight locations, only one haplotype was found. Furthermore, for all individuals, P. major had large Hd and Pi values (Hd > 0.5 and Pi > 0.005), indicating a large stable population with an extended evolutionary history. In contrast, P. tectus had smaller Hd and Pi values (Hd < 0.5 and Pi < 0.005), implying recent founder events [37,38,39,40].
3.4. Phylogenetic Relationship and Haplotype Network
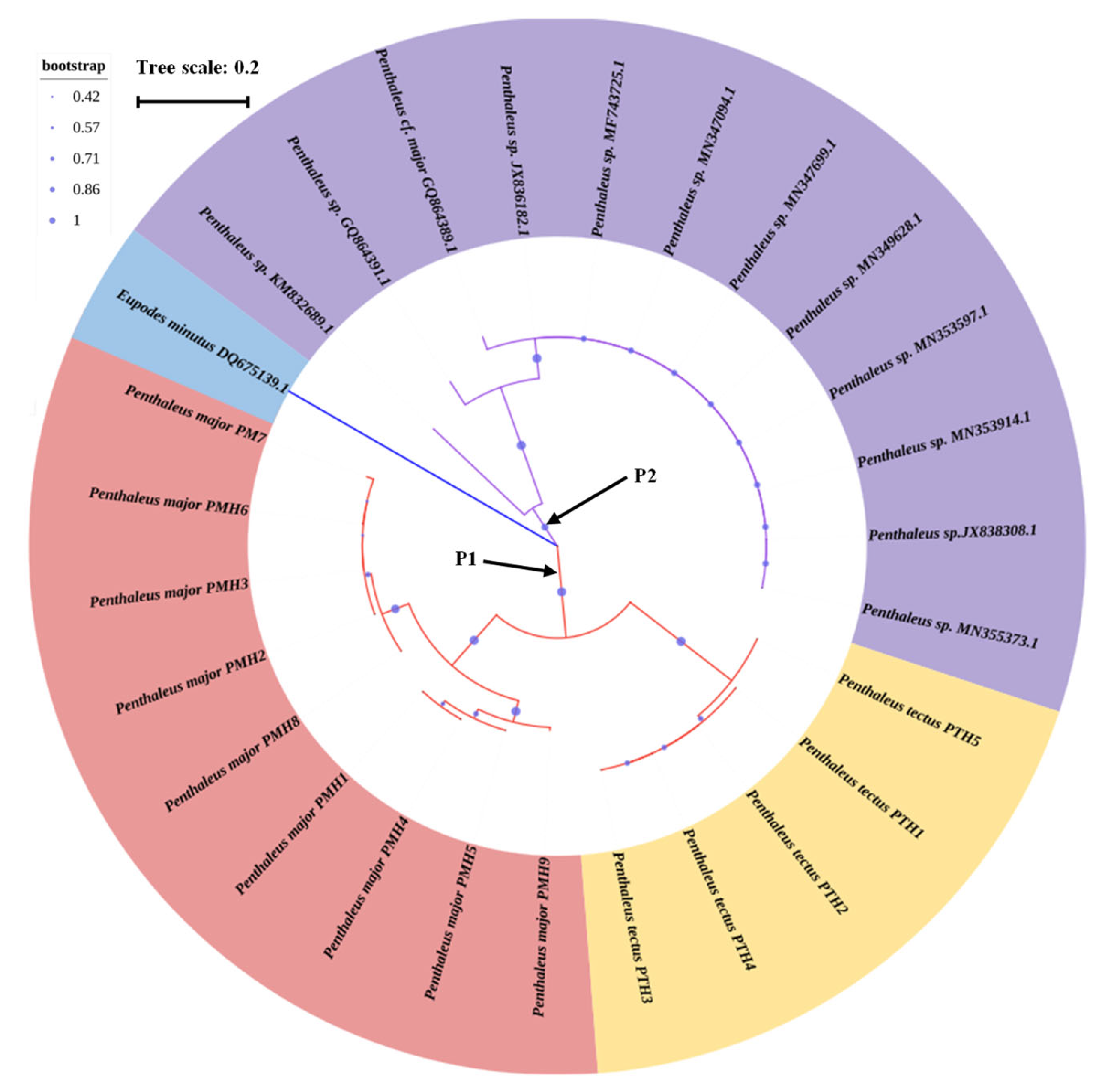
The Neighbor-Joining method was used to construct the phylogenetic tree with COI sequences of P. major, P. tectus, and 12 published COI sequences of Penthaleus sp. (Figure 3). Among the 26 haplotypes of Penthaleus sp., Clade P1 (PMH1-PMH9 and PTH1-PTH5 from China) and Clade P2 (other samples from Canada and Poland) were identified. The COI sequence of Eupodes minutus with GenBank accession number DQ675139.1 was chosen as the outgroup. The phylogenetic tree showed that the Penthaleus sp. in China were divided into two branches distinct from those in Canada and Poland, while the 14 haplotypes in China were split into two branches, representing P. major (PMH1-PMH9) and P. tectus (PTH1-PTH5), respectively.
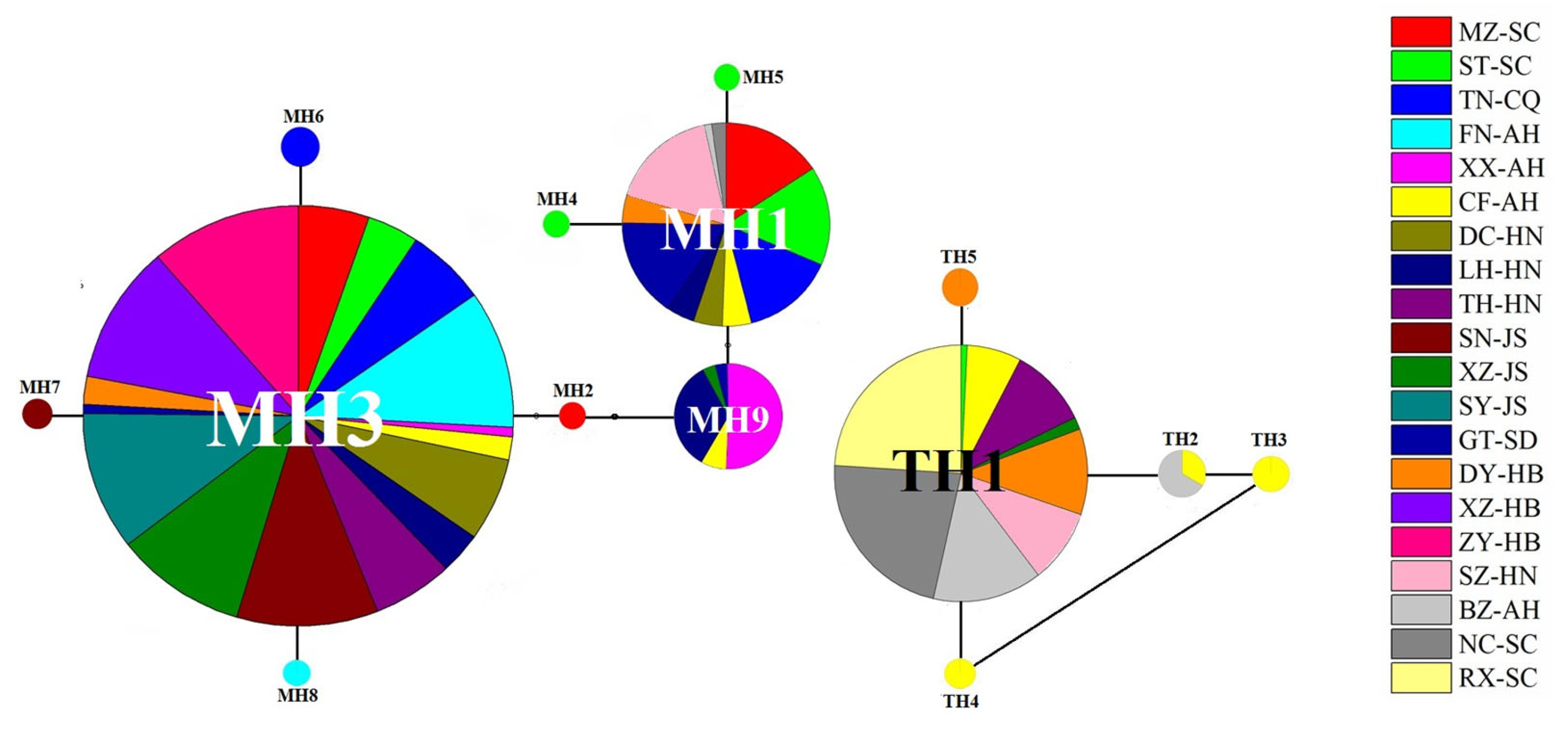
The haplotypes of P. major and P. tectus populations are shown by the TCS network, with varying colors and sizes of circles representing different sampling sites and the number of individuals detected. MH3 is the most crucial haplotype for P. major, and TH1 is the most abundant haplotype for P. tectus (Figure 4).
3.5. Population Differentiation and Genetic Structure
Population pairwise fixation index (FST) and gene flow (Nm) investigated the diversity in 21 P. major populations and 11 P. tectus populations (Table 4 and Table 5). The pairwise FST tests of P. major populations indicated significant differentiation in 120 of 210 location pairs based on mitochondrial COI gene.
Gene flow between SW and YV of P. major and between SW and YH of P. tectus show a relatively low degree due to the low Nm value (Nm < 1), suggesting that genetic drift might result in genetic differentiation (Table 6 and Table 7). For three P. major natural geographic groups, higher gene flow (Nm > 1) was found between SW and YH, while for P. major and P. tectus, high gene flow (Nm > 3) was found between YV and YH. In addition, for P. tectus, high gene flow (Nm = 4.23) was also found between SW and YV.
AMOVA results of P. major indicated significant molecular genetic variations among the locations within geographic regions (55.74%) (Table 8). The variation within sites was 42.20%. Only 2.05% variation was found among geographic regions. However, for P. tectus, the AMOVA results indicated that a significant portion of the molecular genetic variation was found within locations (88.43%). The variation among the sites within geographic regions was 9.21%. Only 2.35% variation was found among geographic areas.
3.6. Demographic Analysis
Demographic analysis showed that the overall mismatch distribution for all sampled localities showed the multimodal profile (Figure 5), suggesting that P. major populations in China might not have undergone a sudden demographic expansion. The actual observation results are compatible with the predicted constant model when assuming a constant population size for P. tectus, demonstrating that the population status is consistent with the expected model assumption. Altogether, results suggested that the population size of P. major and P. tectus have not changed over time.
4. Discussion
Blue oat mites, which are widely distributed in southern Australia, are an important agricultural pest species in Australia [4,5]. Electrophoresis results suggest that there are differences between species in terms of clonal diversity [11,13,14]. Previous research has also showed that Penthaleus spp. respond differently to pesticides [26,38]. Therefore, clarifying the distribution range and population density of different species is essential for adequate control.
In this study, Penthaleus spp. and Petrobia spp. were found on winter wheat, consistent with previous research reports [23]. A large number of blue oat mites (Penthaleus spp.) were found in three major winter wheat regions of China, including P. major and P. tectus. The distribution range and population density of P. major were broader and higher than P. tectus, suggesting that controlling pest mites on wheat is very difficult because different species have different resistance levels. The risk is that the sympatric occurrence of different species will make control more difficult. We also found high levels of genetic diversity in the P. major population by analyzing mitochondrial cytochrome oxidase subunit I sequences. The distribution proportion of P. major was significantly higher than P. tectus, and the total number of P. major was as high as 75.91% in 577 samples, consistent with the study of Australian scholars [41]. P. tectus only occurs in limited areas and is recorded only from Australia and South Africa [12]. In China, P. tectus was first recorded in this study and was widely distributed only in a few samples, such as Rongxian (RX-SC) and Nanchong (NC-SC).
The number of haplotypes, haplotype diversity (Hd), and nucleotide diversity (Pi) are used for assessing diversity in population-level COI sequencing surveys [42,43,44,45]. These metrics are useful for biodiversity assessment because they can be influenced by a variety of factors [46]. In the current study, the Hd and Pi values suggested that P. tectus may not be an ancient and stable population compared to P. major. The Hd and Pi among all P. tectus samples were lower than those of P. major. The Hd of P. major in Changfeng (CF-AH) was higher than in this study of other locations. In addition, the Pi of P. major in Changfeng (CF-AH) was up to 0.014. Among the P. tectus populations, the Hd and Pi of Changfeng (CF-AH) were the highest (Hd = 0.467, π = 0.001). Compared to P. tectus, the genetic variation of P. major was at a relatively high level, which was confirmed by allozyme loci [13,14].
Clonal diversity is often high in obligate asexual organisms and most research on asexual organisms has focused on the role of environmental heterogeneity in promoting clonal diversity [13,46,47]. The blue oat mite P. major is asexual and lacks a sexual relative. Populations consist of numerous clonal genotypes and clone frequencies change over time and space [13]. Previous studies have also provided direct evidence for negative frequency-dependent selection maintaining clonal variation in P. major [15]. In our research, haplotype 3 (MH3) was found in 17 P. major populations and haplotype 1 (TH1) was the most abundant in P. tectus populations. The different haplotypes represent different genetic backgrounds, and clone competition, differential resource utilization, and pesticide resistance likely cause the haplotypes’ diversity [48,49,50]. A larger haplotype variation was found in P. major than P. tectus populations, indicating that it has a stronger evolutionary potential for adaptation to future environmental changes and chemical pesticides.
We observed significant differentiation of P. major among regions, locations within regions, and within locations. The percentage of variation was only 2.05% between regions, but as much as 42.20% within locations. For P. tectus, the percentage of variation was up to 88.43% within locations. AMOVA analysis indicated a low level of genetic differentiation among regions of P. major (FCT = 0.02, p = 0.3323) and P. tectus (FCT = 0.02, p = 0.2063), which agrees with previous studies [14]. This may be due to the long-range movement of diapause eggs through winds or human assistance in summer [5,51].
5. Conclusions
In this study, the occurrence pattern of blue oat mites was clarified in three major winter wheat regions. The results indicated that the occurrence of blue oat mites was particularly serious in the Yellow and Huai River winter wheat zone of China, especially during the early spring season. Among the pest mites, P. major was found to be the most common in winter wheat, followed by P. tectus. Although P. tectus had not been previously reported in China, this study found that it occurred in all three wheat regions, and there was an evident co-occurrence between the P. major and P. tectus. Furthermore, the COI sequence analysis revealed that P. major boasts more genotypes and higher genetic diversity than P. tectus. It suggested that P. major may have stronger adaptability. Further studies should focus on exploring the tolerance of P. major and P. tectus to temperature, the fitness on different host plants, and the resistance to various chemicals. This way, we can develop a more reasonable scheme for regional precise control of these mites.
Author Contributions
Conceptualization, X.W., Y.L. and H.L.; methodology, X.W. and Y.L.; software, X.W.; formal analysis, X.W.; investigation, X.W., W.W., M.W. and Y.Q.; data curation, H.L.; writing—original draft preparation, X.W. and Y.L.; writing—review and editing, Y.L. and H.L.; visualization, X.W. and Y.Q.; supervision, H.L.; project administration, Y.L. and H.L.; funding acquisition, Y.L. and H.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key R&D Program of China (2022YFC2601000), the National Natural Science Foundation of China (32072483), the Fundamental Research Funds for the Central Universities (SWU-KQ22019), the Scientific Projects of Science and Technology Department of Tibet, China (ZX202101ZY0006N), and Chongqing Municipal Training Program of Innovation and Entrepreneurship for Undergraduates (S202110635027).
Data Availability Statement
The data generated during the study have already been reported in the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chada, H.L. Biology of the winter grain mite and its control in small grains. J. Econ. Entomol. 1956, 49, 515–520. [Google Scholar] [CrossRef]
- Narayan, D.S. Morphological, biological and ecological studies on the winter grain mite, Penthaleus major (Dugés), Penthaleidae; Acarina. Part 1. J. Zool. Soc. India 1962, 14, 45–63. [Google Scholar]
- Wallace, M.M.H.; Mahon, J.A. The distribution of Halotydeus destructor and Penthaleus major (Acari: Eupodidae) in Australia in relation to climate and land use. Aust. J. Zool. 1971, 19, 65–76. [Google Scholar] [CrossRef]
- Robinson, M.T.; Hoffmann, A.A. The pest status and distribution of three cryptic blue oat mite species (Penthaleus spp.) and redlegged earth mite (Halotydeus destructor) in southeastern Australia. Exp. Appl. Acarol. 2001, 25, 699–716. [Google Scholar] [CrossRef] [PubMed]
- Umina, P.A.; Hoffmann, A.A.; Weeks, A.R. Biology, ecology and control of the Penthaleus species complex (Acari: Penthaleidae). Exp. Appl. Acarol. 2004, 34, 211–237. [Google Scholar] [CrossRef] [PubMed]
- Murray, D.A.H.; Clarke, M.B.; Ronning, D.A. Estimating invertebrate pest losses in six major Australian grain crops. Aust. J. Entomol. 2013, 52, 227–241. [Google Scholar] [CrossRef]
- Qin, T.K.; Halliday, R.B. The Australian species of Chromotydaeus Berlese and Penthaleus C. L. Koch (Acarina, Penthaleidae). J. Nat. Hist. 1996, 30, 1833–1848. [Google Scholar] [CrossRef]
- Qin, T.K.; Halliday, R.B. Revision of the Australian and South African species of Halotydeus (Acarina: Penthaleidae). Bull. Entomol. Res. 1996, 86, 441–450. [Google Scholar] [CrossRef]
- Womersley, H. On the name of the “blue oat mite” of Australia. Bull. Entomol. Res. 1935, 26, 163. [Google Scholar] [CrossRef]
- Weeks, A.R.; Fripp, Y.J.; Hoffmann, A.A. Genetic structure of Halotydeus destructor and Penthaleus major populations in Victoria (Acari: Penthaleidae). Exp. Appl. Acarol. 1995, 19, 633–646. [Google Scholar] [CrossRef]
- Weeks, A.R.; Hoffmann, A.A. The biology of Penthaleus species in southeastern Australia. Entomol. Exp. Appl. 1999, 92, 179–189. [Google Scholar] [CrossRef]
- Halliday, R.B. Systematics and biology of Penthaleus tectus sp. n. (Acari: Penthaleidae), a recently discovered pest of grain crops in eastern Australia. Aust. J. Entomol. 2005, 44, 144–149. [Google Scholar] [CrossRef]
- Weeks, A.R.; Hoffmann, A.A. Intense selection of mite clones in a heterogeneous environment. Evolution 1998, 52, 1325–1333. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.T.; Weeks, A.R.; Hoffmann, A.A. Geographic patterns of clonal diversity in the earth mite species Penthaleus major with particular emphasis on species margins. Evolution 2002, 56, 1160–1167. [Google Scholar] [CrossRef] [PubMed]
- Weeks, A.R.; Hoffmann, A.A. Frequency-dependent selection maintains clonal diversity in an asexual organism. Proc. Natl. Acad. Sci. USA 2008, 105, 17872–17877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankham, R.; Briscoe, D.A.; Ballou, J.D. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Allendorf, F.W.; Luikart, G. Conservation and the Genetics of Populations; Wiley: New York, NY, USA, 2009. [Google Scholar]
- Reed, D.H.; Frankham, R. Correlation between fitness and genetic diversity. Conserv. Biol. 2003, 17, 230–237. [Google Scholar] [CrossRef]
- Vandewoestijne, S.; Schtickzelle, N.; Baguette, M. Positive correlation between genetic diversity and fitness in a large, well-connected metapopulation. BMC Biol. 2008, 6, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoa, N.P.; Hamzah, A.; Nguyen, N.H. Genetic variation and correlated changes in reproductive performance of a red tilapia line selected for improved growth over three generations. Anim. Reprod. Sci. 2017, 184, 94–101. [Google Scholar] [CrossRef]
- Carrazco-Montalvo, A.; Ponce, P.; Villota, S.D.; Quentin, E.; Muñoz-Tobar, S.; Coloma, J.; Cevallos, V. Establishment, Genetic Diversity, and Habitat Suitability of Aedes albopictus Populations from Ecuador. Insects 2022, 13, 305. [Google Scholar] [CrossRef]
- Pramual, P.; Jomkumsing, P.; Wongpakam, K.; Vaisusuk, K.; Chatan, W.; Gomontean, B. Population genetic structure and population history of the biting midge Culicoides mahasarakhamense (Diptera: Ceratopogonidae). Insects 2022, 13, 724. [Google Scholar] [CrossRef]
- Wang, J.Q. Acute symptoms identification methods and prevention methods of mite pests on wheat. Hortic. Seed 2017, 4, 53–55. [Google Scholar]
- He, X.Y. Occurrence and control technology of wheat main pests in Taihe county in 2020. Anhui Agric. Sci. Bull. 2021, 5, 115–117. [Google Scholar] [CrossRef]
- Hill, M.P.; Hoffmann, A.A.; Mccoll, S.A.; Umina, P.A. Distribution of cryptic blue oat mite species in Australia: Current and future climate conditions. Agric. For. Entomol. 2012, 14, 127–137. [Google Scholar] [CrossRef]
- Umina, P.A.; Hoffmann, A.A. Tolerance of cryptic species of blue oat mites (Penthaleus spp.) and the redlegged earth mite (Halotydeus destructor) to pesticides. Aust. J. Exp. Agric. 1999, 39, 621–628. [Google Scholar] [CrossRef]
- Wang, J. The life tables of laboratory population of Frankliniella occidentalis and Thrips tabaci on purple cabbage. Acta. Phytophyl. Sin. 2011, 27, 300–302. [Google Scholar]
- Li, X.; Zhang, Z.; Zhang, J.; Huang, J.; Wang, L.; Li, Y.; Hafeez, M.; Lu, Y. Population genetic diversity and structure of Thrips tabaci (Thysanoptera: Thripidae) on Allium hosts in China, inferred from mitochondrial COI gene sequences. J. Econ. Entomol. 2020, 113, 1426–1435. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 1673–1680. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Sudhir, K.; Glen, S.; Koichiro, T. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H. Arlequin suite ver3.5: A new series of programs to perform population genetics analyses under linux and windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. Dnasp v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slatkin, M.; Hudson, R.R. Pairwise comparisons of mitochondrial DNA sequences in stable and exponentially growing populations. Genetics 1991, 129, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.R.; Harpending, H. Population growth makes waves in the distribution of pairwise genetic differences. Mol. Biol. Evol. 1992, 9, 552–569. [Google Scholar] [CrossRef]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987. [Google Scholar]
- Goodall-Copestake, W.; Tarling, G.; Murphy, E. On the comparison of population-level estimates of haplotype and nucleotide diversity: A case study using the gene cox1 in animals. Heredity 2012, 109, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.T.; Hoffmann, A.A. Additional tests on the effects of pesticides on cryptic species of blue oat mite (Penthaleus spp.) and the redlegged earth mite (Halotydeus destructor). Aust. J. Exp. Agric. 2000, 40, 671–678. [Google Scholar] [CrossRef]
- Fox, J.A.; Dybdahl, M.F.; Jokela, J.; Lively, C.M. Genetic structure of coexisting sexual and clonal subpopulations in a freshwater snail (Potamopyrgus antipodarum). Evolution 1996, 50, 1541–1548. [Google Scholar]
- Herbert, P.D.N.; Crease, T.J. Clonal coexistence in Daphnia pulex (Leydig): Another planktonic paradox. Science 1980, 207, 1363–1365. [Google Scholar] [CrossRef]
- Turner, B.J.; Elder, J.F.J.; Laughlin, T.F.; Davis, W.P.; Taylor, D.S. Extreme clonal diversity and divergence in populations of a selfing hermaphroditic fish. Proc. Natl. Acad. Sci. USA 1992, 89, 10643–10647. [Google Scholar] [CrossRef] [Green Version]
- Vrijenhoek, R.C. Factors affecting clonal diversity and coexistence. Amer. Zool. 1979, 19, 787–797. [Google Scholar] [CrossRef] [Green Version]
- Levene, H. Genetic equilibrium when more than one ecological niche is available. Am. Nat. 1953, 87, 331–333. [Google Scholar] [CrossRef]
- Strobeck, C. Sufficient conditions for polymorphism with N niches and M mating groups. Am. Nat. 1974, 108, 152–156. [Google Scholar] [CrossRef]
- Cao, L.J.; Gao, Y.F.; Gong, Y.J.; Chen, J.C.; Chen, M.; Hoffmann, A.; Wei, S.J. Population analysis reveals genetic structure of an invasive agricultural Thrips pest related to invasion of greenhouses and suitable climatic space. Evol. Appl. 2019, 12, 1868–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.J.; Cai, W.Z.; Luo, J.Y.; Zhang, S.; Li, W.D.; Wang, C.Y.; Lv, L.M.; Cui, J.J. Population genetic structure and expansion patterns of the cotton pest Adelphocoris fasciaticollis. J. Pest. Sci. 2018, 91, 539–550. [Google Scholar] [CrossRef]
- Zamoum, T.; Simon, J.C.; Crochard, D.; Ballanger, Y.; Lapchin, L.; Vanlerberghe-Masutti, F.; Guillemaud, T. Does insecticide resistance alone account for the low genetic variability of asexually reproducing populations of the peach-potato aphid Myzus persicae? Heredity 2005, 94, 630–639. [Google Scholar] [CrossRef]
- RidsdillSmith, T.J.; Annells, A.J. Seasonal occurrence and abundance of redlegged earth mite Halotydeus destructor (Acari: Penthaleidae) in annual pastures of southwestern Australia. Bull. Entomol. Res. 1997, 87, 413–423. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Investigation map of pest mites on wheat in main winter wheat areas of China, including survey areas and specific survey sites. The picture was created using the online website SimpleMappr (https://www.simplemappr.net accessed on 16 December 2022) with latitude and longitude data.
Figure 1.
Investigation map of pest mites on wheat in main winter wheat areas of China, including survey areas and specific survey sites. The picture was created using the online website SimpleMappr (https://www.simplemappr.net accessed on 16 December 2022) with latitude and longitude data.

Figure 2.
Location map of the studied wheat populations of P. major and P. tectus in Southwestern China (SW), Middle and Lower Yangtze Valleys (YV), and Yellow and Huai Valleys (YH), which are the three major winter wheat areas of China. The picture was created using the online website SimpleMappr (https://www.simplemappr.net accessed on 16 December 2022) with latitude and longitude data. The haplotype network diagram was created in PowerPoint.
Figure 2.
Location map of the studied wheat populations of P. major and P. tectus in Southwestern China (SW), Middle and Lower Yangtze Valleys (YV), and Yellow and Huai Valleys (YH), which are the three major winter wheat areas of China. The picture was created using the online website SimpleMappr (https://www.simplemappr.net accessed on 16 December 2022) with latitude and longitude data. The haplotype network diagram was created in PowerPoint.

Figure 3.
The Neighbor-Joining phylogenetic relationship of 9 haplotypes of P. major, 5 haplotypes of P. tectus, and 12 haplotypes of P. sp. The scale bar indicates the average number of nucleotide substitutions per site. The picture was first drawn in MEGA 7.0, and then the picture was adjusted through the online website (https://itol.embl.de/ accessed on 24 March 2023).
Figure 3.
The Neighbor-Joining phylogenetic relationship of 9 haplotypes of P. major, 5 haplotypes of P. tectus, and 12 haplotypes of P. sp. The scale bar indicates the average number of nucleotide substitutions per site. The picture was first drawn in MEGA 7.0, and then the picture was adjusted through the online website (https://itol.embl.de/ accessed on 24 March 2023).

Figure 4.
The TCS network for haplotypes of P. major and P. tectus populations based on the sequence of the COI gene. The haplotype network diagram was constructed in TCS 1.21, and then the color was added in PowerPoint.
Figure 4.
The TCS network for haplotypes of P. major and P. tectus populations based on the sequence of the COI gene. The haplotype network diagram was constructed in TCS 1.21, and then the color was added in PowerPoint.

Figure 5.
Mismatch distribution analysis of P. major and P. tectus from 23 locations based on COI gene.
Figure 5.
Mismatch distribution analysis of P. major and P. tectus from 23 locations based on COI gene.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Sample collection information for P. major and P. tectus used in this study.
| Region | Pop. ID | Population | Longitude (°E) | Latitude (°N) | Number of P. major | Number of P. tectus |
|---|---|---|---|---|---|---|
| SW (SC, CQ) | RX-SC | Sichuan, Rongxian | 104°27′ | 29°25′ | 31 | 0 |
| MZ-SC | Sichuan, Mianzhu | 104°14′ | 31°20′ | 30 | 0 | |
| ST-SC | Sichuan, Santai | 105°08′ | 31°10′ | 27 | 1 | |
| NC-SC | Sichuan, Nanchong | 106°05′ | 30°77′ | 2 | 29 | |
| TN-CQ | Chongqing, Tongnan | 105°84′ | 30°19′ | 32 | 0 | |
| YV (HB) | DY-HB | Hubei, Dangyang | 111°55′ | 30°51′ | 10 | 16 |
| XZ-HB | Hubei, Xiangzhou | 112°21′ | 32°07′ | 30 | 0 | |
| ZY-HB | Hubei, Zaoyang | 112°51′ | 32°12′ | 32 | 0 | |
| ZX-HB | Hubei, Zhongxiang | 112°36′ | 31°13′ | 4 | 0 | |
| YH (JS, AH, HN, SD, SX) | SN-JS | Jiangsu, Suining | 117°95′ | 33°90′ | 31 | 0 |
| XZ-JS | Jiangsu, Xuzhou | 117°14′ | 34°22′ | 28 | 2 | |
| SY-JS | Jiangsu, Siyang | 118°47′ | 33°46′ | 30 | 0 | |
| DH-JS | Jiangsu, Donghai | 118°29′ | 34°34′ | 3 | 1 | |
| FN-AH | Anhui, Funan | 115°58′ | 32°63′ | 31 | 0 | |
| BZ-AH | Anhui, Bozhou | 115°78′ | 33°85′ | 1 | 20 | |
| CF-AH | Anhui, Changfeng | 117°13′ | 32°26′ | 13 | 13 | |
| XX-AH | Anhui, Xiaoxian | 116°51′ | 34°12′ | 29 | 0 | |
| DC-HN | Henan, Dancheng | 115°07′ | 33°43′ | 22 | 0 | |
| SZ-HN | Henan, Shanzhou | 111°04′ | 34°43′ | 17 | 12 | |
| TH-HN | Henan, Tanghe | 112°44′ | 32°30′ | 17 | 13 | |
| LH-HN | Henan, Luohe | 113°54′ | 33°34′ | 31 | 0 | |
| GT-SD | Shandong, Gaotang | 116°25′ | 36°80′ | 18 | 0 | |
| RC-SX | Shanxi, Ruicheng | 110°68′ | 34°70′ | 0 | 1 |
The SW, YV, and YH stand for Southwestern China, Middle and Lower Yangtze Valleys, and Yellow and Huai Valleys, respectively.
Table 2.
Variation analysis of COI gene sequences of P. major and P. tectus.
| Species | Length of Sequences (bp) | C | V | PI | S | A + T (%) |
|---|---|---|---|---|---|---|
| P. major | 657–658 | 636 | 22 | 19 | 3 | 70.30 |
| P. tectus | 657–658 | 654 | 4 | 4 | None | 69.90 |
C: Constant sites; V: variable sites; PI: Parsimony-informative sites; S: Singleton sites.
Table 3.
Genetic diversity of P. major and P. tectus from different locations based on mitochondrial COI gene.
Table 3.
Genetic diversity of P. major and P. tectus from different locations based on mitochondrial COI gene.
| Species | Pop.ID | H | Haplotype (No. of Specimens) | Hd | Pi | K |
|---|---|---|---|---|---|---|
| P. major | MZ-SC | 3 | MH1 (14), MH2 (1), MH3 (15) | 0.549 | 0.014 | 9.274 |
| ST-SC | 4 | MH1 (14), MH3 (11), MH4 (1), MH5 (1) | 0.584 | 0.014 | 9.111 | |
| NC-SC | 1 | MH1 (2) | 0 | 0 | 0 | |
| TN-CQ | 3 | MH1 (13), MH3 (17), MH6 (2) | 0.567 | 0.014 | 9.085 | |
| DY-HB | 2 | MH1 (4), MH3 (6) | 0.513 | 0.014 | 9.231 | |
| XZ-HB | 1 | MH3 (30) | 0 | 0 | 0 | |
| ZY-HB | 1 | MH3 (32) | 0 | 0 | 0 | |
| ZX-HB | 1 | MH1 (4) | 0 | 0 | 0 | |
| SN-JS | 2 | MH3 (30), MH7 (1) | 0.065 | 0 | 0.065 | |
| XZ-JS | 1 | MH3 (28) | 0 | 0 | 0 | |
| SY-JS | 1 | MH3 (30) | 0 | 0 | 0 | |
| DH-JS | 1 | MH3 (3) | 0 | 0 | 0 | |
| FN-AH | 2 | MH3 (30), MH8 (1) | 0.065 | 0 | 0.065 | |
| BZ-AH | 1 | MH1 (1) | 0 | 0 | 0 | |
| CF-AH | 3 | MH1 (4), MH3 (5), MH9 (4) | 0.708 | 0.014 | 8.917 | |
| XX-AH | 2 | MH3 (2), MH9 (27) | 0.114 | 0.003 | 1.825 | |
| DC-HN | 2 | MH1 (4), MH3 (18) | 0.280 | 0.008 | 5.040 | |
| SZ-HN | 2 | MH1 (15), MH9 (2) | 0.221 | 0 | 0.441 | |
| TH-HN | 1 | MH3 (17) | 0 | 0 | 0 | |
| LH-HN | 3 | MH1 (4), MH3 (9), MH9 (18) | 0.581 | 0.011 | 7.277 | |
| GT-SD | 3 | MH1 (14), MH3 (2), MH9 (2) | 0.288 | 0.007 | 4.275 | |
| P. tectus | RX-SC | 1 | TH1 (31) | 0 | 0 | 0 |
| ST-SC | 1 | TH1 (1) | 0 | 0 | 0 | |
| NC-SC | 1 | TH1 (29) | 0 | 0 | 0 | |
| DY-HB | 2 | TH1 (14), TH5 (2) | 0.199 | 0.000 | 0.199 | |
| XZ-JS | 1 | TH1 (2) | 0 | 0 | 0 | |
| DH-JS | 1 | TH1 (1) | 0 | 0 | 0 | |
| BZ-AH | 2 | TH1 (18), TH2 (2) | 0.138 | 0.000 | 0.138 | |
| CF-AH | 4 | TH1 (9), TH2 (1), TH3 (2), TH4 (1) | 0.467 | 0.001 | 0.686 | |
| SZ-HN | 1 | TH1 (12) | 0 | 0 | 0 | |
| TH-HN | 1 | TH1 (13) | 0 | 0 | 0 | |
| RC-SX | 1 | TH1 (1) | 0 | 0 | 0 |
Table 4.
Pairwise FST (below diagonal) and gene flow Nm (above diagonal) among 21 populations of P. major in winter wheat of China.
Table 4.
Pairwise FST (below diagonal) and gene flow Nm (above diagonal) among 21 populations of P. major in winter wheat of China.
| Locations | MZ- SC | ST- SC | NC- SC | TN- CQ | DY- HB | XZ- HB | ZY- HB | ZX- HB | SN- JS | XZ- JS | SY- JS | DH-JS | FN- AH | BZ- AH | CF- AH | XX- AH | DC- HN | SZ- HN | TH- HN | LH- HN | GT- SD |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MZ-SC | inf | 0.84 | inf | inf | 0.30 | 0.29 | 0.75 | 0.30 | 0.31 | 0.30 | 0.91 | 0.30 | 9.01 | inf | 0.41 | 1.62 | 0.31 | 0.40 | 2.80 | 0.64 | |
| ST-SC | −0.01 | 3.32 | 7.33 | 83.08 | 0.18 | 0.17 | 0.40 | 0.17 | 0.18 | 0.18 | 0.45 | 0.17 | inf | inf | 0.75 | 0.70 | 0.51 | 0.23 | inf | 1.36 | |
| NC-SC | 0.23 | 0.07 | 0.55 | 0.56 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | inf | 10.62 | inf | inf | inf | inf | inf | inf | |
| TN-CQ | −0.02 | 0.03 | 0.31 | inf | 0.41 | 0.40 | 1.15 | 0.41 | 0.43 | 0.41 | 1.50 | 0.41 | 1.36 | 7.56 | 0.31 | 3.13 | 0.25 | 0.55 | 1.50 | 0.46 | |
| DY-HB | −0.06 | 0.00 | 0.31 | −0.07 | 0.21 | 0.20 | 1.11 | 0.21 | 0.22 | 0.21 | 1.67 | 0.21 | 2.00 | inf | 0.22 | 4.75 | 0.14 | 0.33 | 1.70 | 0.38 | |
| XZ-HB | 0.45 | 0.59 | 1.00 | 0.38 | 0.55 | inf | inf | inf | inf | inf | inf | inf | 0.00 | 0.10 | 0.02 | 1.17 | 0.00 | inf | 0.11 | 0.02 | |
| ZY-HB | 0.46 | 0.60 | 1.00 | 0.38 | 0.56 | 0.00 | inf | 249.8 | inf | inf | inf | 249.8 | 0.00 | 0.10 | 0.02 | 1.12 | 0.00 | inf | 0.11 | 0.02 | |
| ZX-HB | 0.25 | 0.39 | 1.00 | 0.18 | 0.18 | 0.00 | 0.00 | inf | inf | inf | inf | inf | 0.00 | 0.33 | 0.04 | inf | 0.00 | inf | 0.23 | 0.06 | |
| SN-JS | 0.46 | 0.59 | 1.00 | 0.38 | 0.55 | 0.00 | 0.00 | −0.14 | inf | inf | inf | inf | 0.00 | 0.10 | 0.02 | 1.16 | 0.00 | inf | 0.11 | 0.02 | |
| XZ-JS | 0.44 | 0.58 | 1.00 | 0.37 | 0.53 | 0.00 | 0.00 | 0.00 | 0.00 | inf | inf | inf | 0.00 | 0.11 | 0.02 | 1.24 | 0.00 | inf | 0.11 | 0.03 | |
| SY-JS | 0.45 | 0.59 | 1.00 | 0.38 | 0.55 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | inf | inf | 0.00 | 0.10 | 0.02 | 1.17 | 0.00 | inf | 0.11 | 0.02 | |
| DH-JS | 0.22 | 0.36 | 1.00 | 0.14 | 0.13 | 0.00 | 0.00 | 0.00 | −0.02 | 0.00 | 0.00 | inf | 0.00 | 0.38 | 0.04 | inf | 0.00 | inf | 0.23 | 0.06 | |
| FN-AH | 0.46 | 0.59 | 1.00 | 0.38 | 0.55 | 0.00 | 0.00 | −0.14 | 0.00 | 0.00 | 0.00 | −0.02 | 0.00 | 0.10 | 0.02 | 1.16 | 0.00 | inf | 0.11 | 0.02 | |
| BZ-AH | 0.03 | −0.23 | 0.00 | 0.16 | 0.11 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | inf | inf | 0.15 | inf | 0.00 | inf | inf | |
| CF-AH | −0.01 | −0.06 | 0.02 | 0.03 | 0.00 | 0.71 | 0.72 | 0.43 | 0.71 | 0.70 | 0.71 | 0.40 | 0.71 | −0.33 | 0.72 | 0.59 | 0.41 | 0.15 | inf | 1.59 | |
| XX-AH | 0.38 | 0.25 | −0.30 | 0.45 | 0.54 | 0.93 | 0.93 | 0.88 | 0.93 | 0.93 | 0.93 | 0.87 | 0.93 | −0.93 | 0.26 | 0.09 | 27.53 | 0.02 | 1.80 | inf | |
| DC-HN | 0.13 | 0.26 | 0.66 | 0.07 | 0.05 | 0.17 | 0.18 | −0.03 | 0.18 | 0.17 | 0.18 | −0.08 | 0.18 | 0.62 | 0.30 | 0.73 | 0.07 | 1.85 | 0.36 | 0.14 | |
| SZ-HN | 0.44 | 0.33 | 0.00 | 0.50 | 0.64 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 0.00 | 0.38 | 0.01 | 0.79 | 0.00 | 0.97 | 4.8 | |
| TH-HN | 0.38 | 0.52 | 1.00 | 0.31 | 0.43 | 0.00 | 0.00 | 0.00 | −0.02 | 0.00 | 0.00 | 0.00 | −0.02 | 1.00 | 0.62 | 0.91 | 0.12 | 1.00 | 0.14 | 0.03 | |
| LH-HN | 0.08 | 0.00 | −0.07 | 0.14 | 0.13 | 0.70 | 0.70 | 0.54 | 0.70 | 0.69 | 0.70 | 0.52 | 0.70 | −0.47 | −0.04 | 0.12 | 0.41 | 0.21 | 0.64 | 5.18 | |
| GT-SD | 0.28 | 0.16 | −0.27 | 0.35 | 0.40 | 0.91 | 0.91 | 0.81 | 0.91 | 0.91 | 0.91 | 0.80 | 0.91 | −0.88 | 0.14 | −0.04 | 0.65 | 0.05 | 0.88 | 0.05 |
Table 5.
Pairwise FST (below diagonal) and gene flow Nm (above diagonal) among 10 populations of P. tectus in winter wheat of China.
Table 5.
Pairwise FST (below diagonal) and gene flow Nm (above diagonal) among 10 populations of P. tectus in winter wheat of China.
| Locations | RX-SC | ST-SC | NC-SC | XZ-JS | BZ-AH | CF-AH | SZ-HN | TH-HN | RC-SX | DY-HB |
|---|---|---|---|---|---|---|---|---|---|---|
| RX-SC | inf | inf | inf | 2.63 | 0.58 | inf | inf | inf | 1.63 | |
| ST-SC | 0.00 | inf | inf | inf | inf | inf | inf | inf | inf | |
| NC-SC | 0.00 | 0.00 | inf | 2.83 | 0.62 | inf | inf | inf | 1.74 | |
| XZ-JS | 0.00 | 0.00 | 0.00 | inf | inf | inf | inf | inf | inf | |
| DH-JS | 0.00 | 0.00 | 0.00 | 0.00 | inf | inf | inf | inf | inf | |
| BZ-AH | 0.09 | −0.89 | 0.08 | −0.28 | 2.35 | 15.00 | 11.04 | inf | 3.76 | |
| CF-AH | 0.30 | −0.67 | 0.29 | −0.16 | 0.10 | 1.35 | 1.25 | inf | 1.3 | |
| SZ-HN | 0.00 | 0.00 | 0.00 | 0.00 | 0.02 | 0.16 | inf | inf | 5.79 | |
| TH-HN | 0.00 | 0.00 | 0.00 | 0.00 | 0.02 | 0.17 | 0.00 | inf | 4.92 | |
| RC-SX | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | −0.89 | −0.67 | 0.00 | inf | |
| DY-HB | 0.13 | −0.87 | 0.13 | −0.27 | 0.06 | 0.16 | 0.04 | 0.05 | -0.87 |
Table 6.
Pairwise FST (below diagonal) and gene flow Nm (above diagonal) among different geographic regions of P. major based on mitochondrial.
Table 6.
Pairwise FST (below diagonal) and gene flow Nm (above diagonal) among different geographic regions of P. major based on mitochondrial.
| Locations | Southwestern China (SW) | Middle and Lower Yangtze Valleys (YV) | Yellow and Huai Valleys (YH) |
|---|---|---|---|
| Southwestern China (SW) | 0.43819 | 1.08303 | |
| Middle and Lower Yangtze Valleys (YV) | 0.36327 | 3.81602 | |
| Yellow and Huai Valleys (YH) | 0.18754 | 0.06149 |
Table 7.
Pairwise FST (below diagonal) and gene flow Nm (above diagonal) among different geographic regions of P. tectus based on mitochondrial.
Table 7.
Pairwise FST (below diagonal) and gene flow Nm (above diagonal) among different geographic regions of P. tectus based on mitochondrial.
| Locations | Southwestern China (SW) | Middle and Lower Yangtze Valleys (YV) | Yellow and Huai Valleys (YH) |
|---|---|---|---|
| Southwestern China (SW) | 4.23242 | 0.84639 | |
| Middle and Lower Yangtze Valleys (YV) | 0.05577 | 4.07018 | |
| Yellow and Huai Valleys (YH) | 0.22802 | 0.05787 |
Table 8.
Analysis of molecular variance (AMOVA) of COI gene sequences in different geographic regions of P. major and P. tectus.
Table 8.
Analysis of molecular variance (AMOVA) of COI gene sequences in different geographic regions of P. major and P. tectus.
| Species | Source of Variation | df | Sum of Squares | % of Variation | Fixation Indices | p-Value |
|---|---|---|---|---|---|---|
| P. major | Among regions | 2 | 152.02 | 2.05 | FCT = 0.02 | p = 0.332 |
| Among locations within regions | 18 | 859.49 | 55.74 | FSC = 0.57 | p < 0.001 | |
| Within locations | 417 | 733.64 | 42.20 | FST = 0.58 | p < 0.001 | |
| P. tectus | Among regions | 2 | 0.53 | 2.35 | FCT = 0.02 | p = 0.206 |
| Among locations within regions | 8 | 1.04 | 9.21 | FSC = 0.09 | p = 0.239 | |
| Within locations | 128 | 8.17 | 88.43 | FST = 0.12 | p = 0.070 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, X.; Wang, W.; Qin, Y.; Wang, M.; Li, Y.; Liu, H. Population Genetic Diversity of Two Blue Oat Mite Species on Triticum Hosts in China. Insects 2023, 14, 377. https://doi.org/10.3390/insects14040377
AMA Style
Wang X, Wang W, Qin Y, Wang M, Li Y, Liu H. Population Genetic Diversity of Two Blue Oat Mite Species on Triticum Hosts in China. Insects. 2023; 14(4):377. https://doi.org/10.3390/insects14040377
Chicago/Turabian StyleWang, Xian, Wenjie Wang, Yang Qin, Mian Wang, Yaying Li, and Huai Liu. 2023. "Population Genetic Diversity of Two Blue Oat Mite Species on Triticum Hosts in China" Insects 14, no. 4: 377. https://doi.org/10.3390/insects14040377
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.