
The Effect of the Sterile Insect Technique on Vibrational Communication: The Case of Bagrada hilaris (Hemiptera: Pentatomidae)
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects Rearing and Irradiation
2.2. Experimental Set-up and Signal Recording
2.3. Data Analysis
2.3.1. Terminology of Vibrational and Behavioral Parameters
2.3.2. Statistical Analysis
3. Results
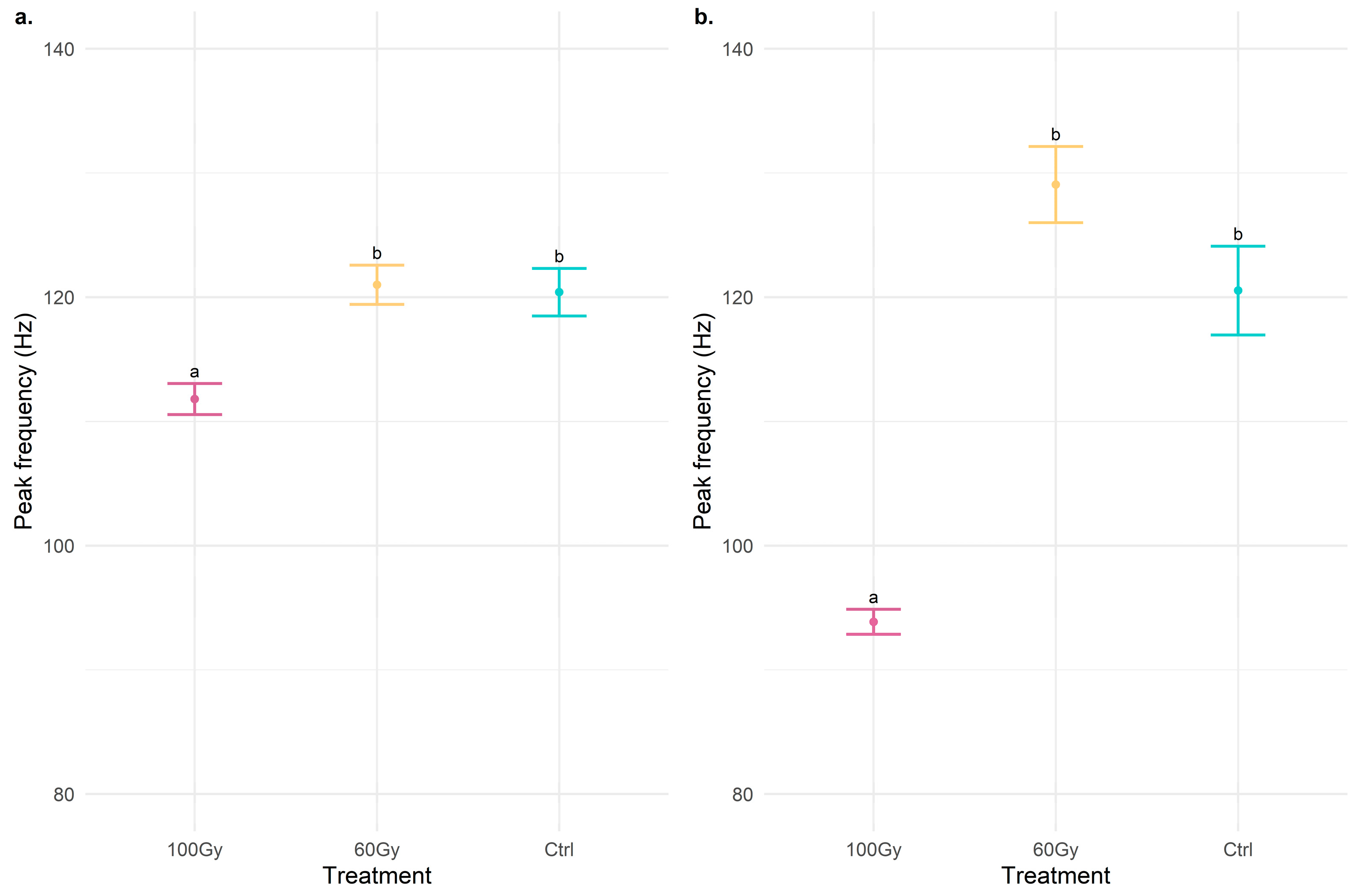
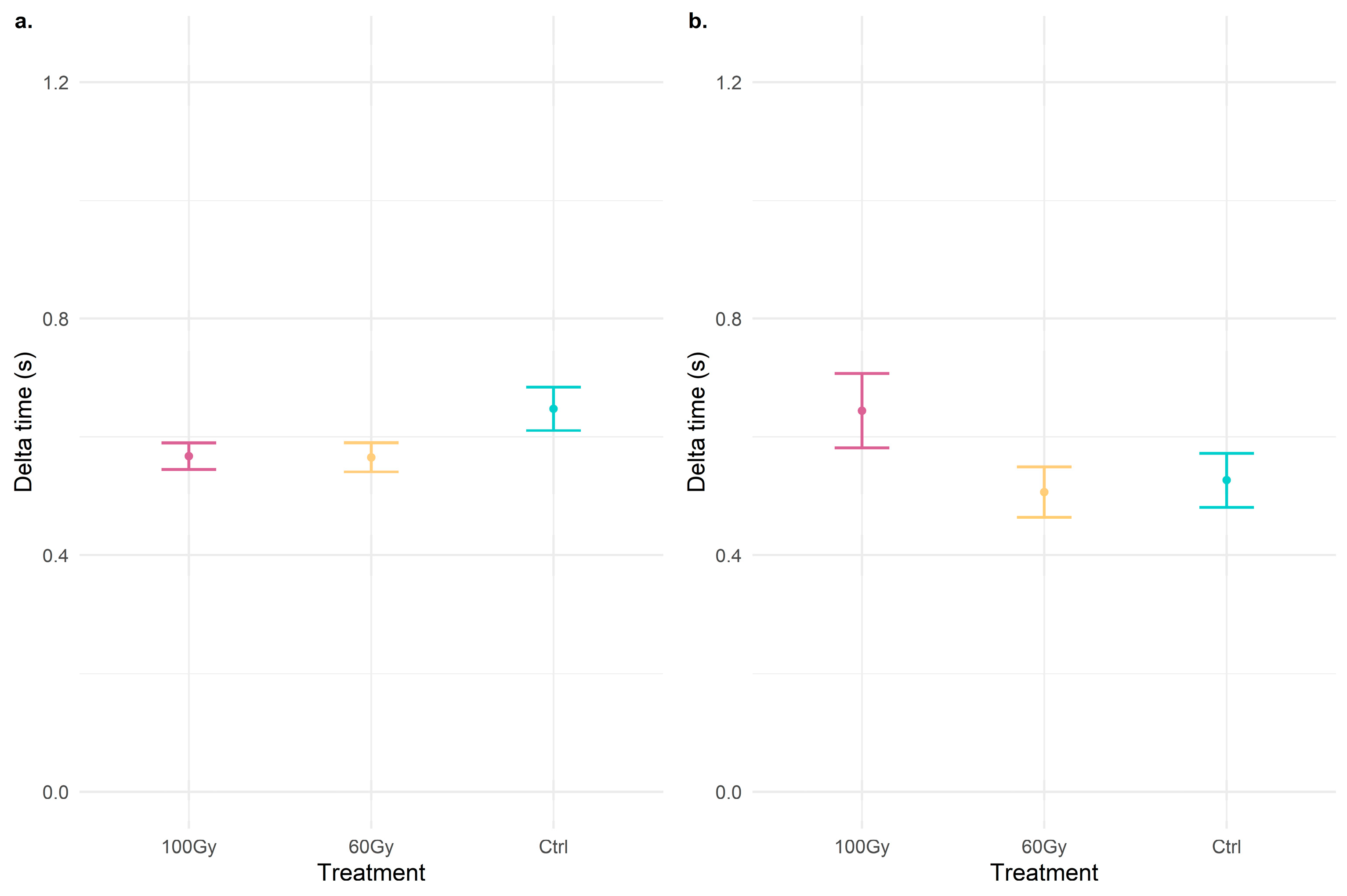
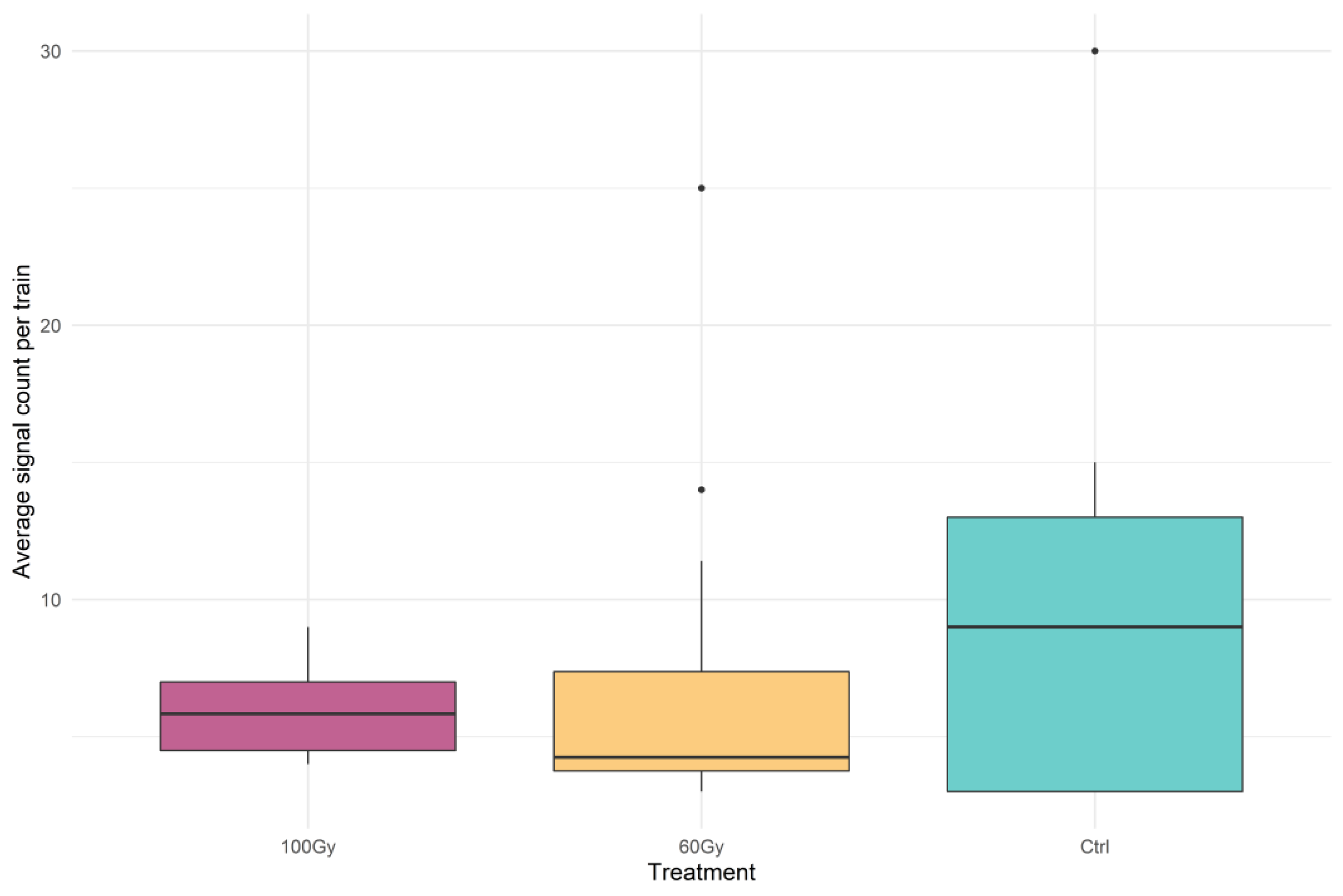
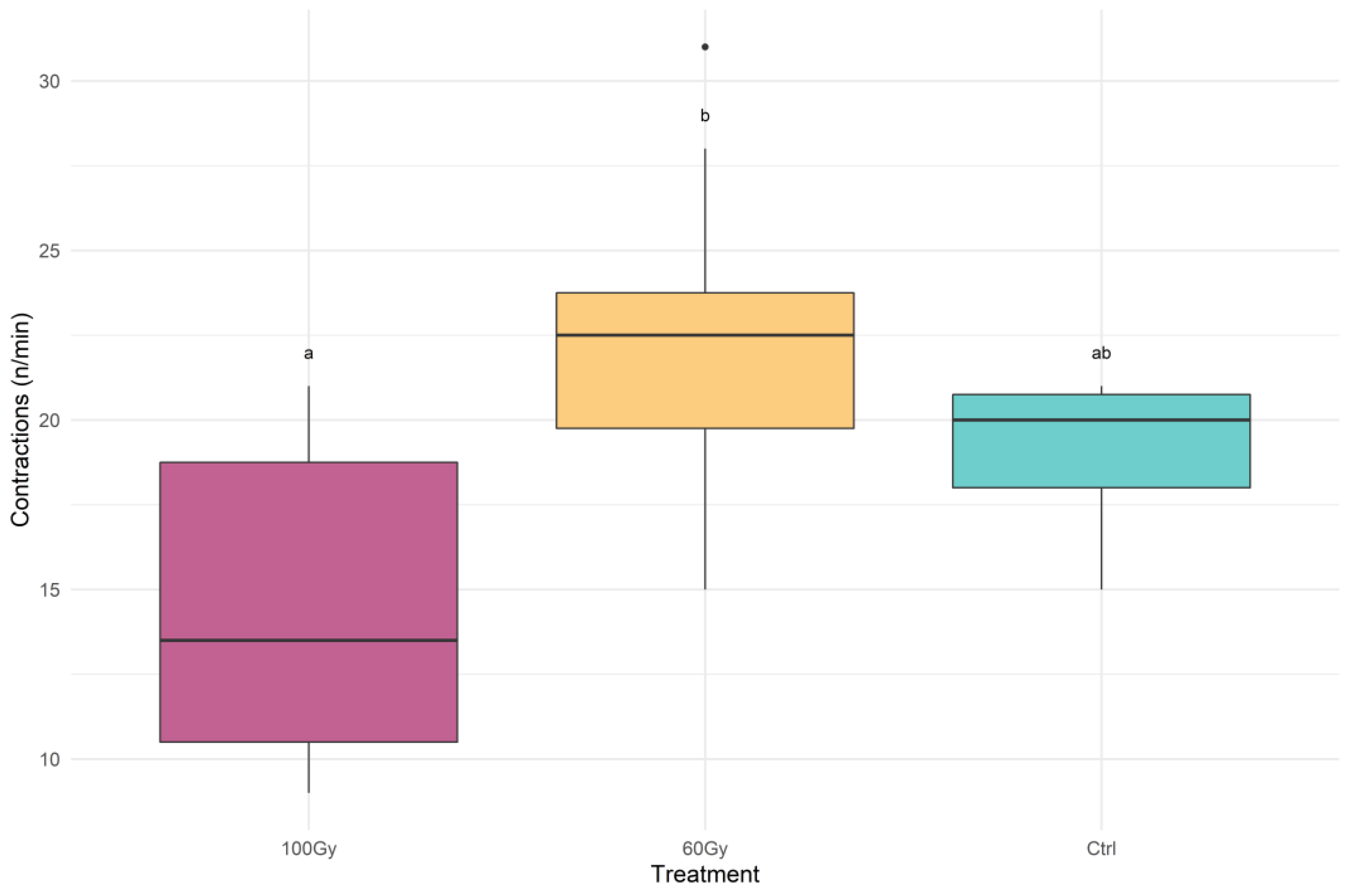
3.1. Vibrational Communication
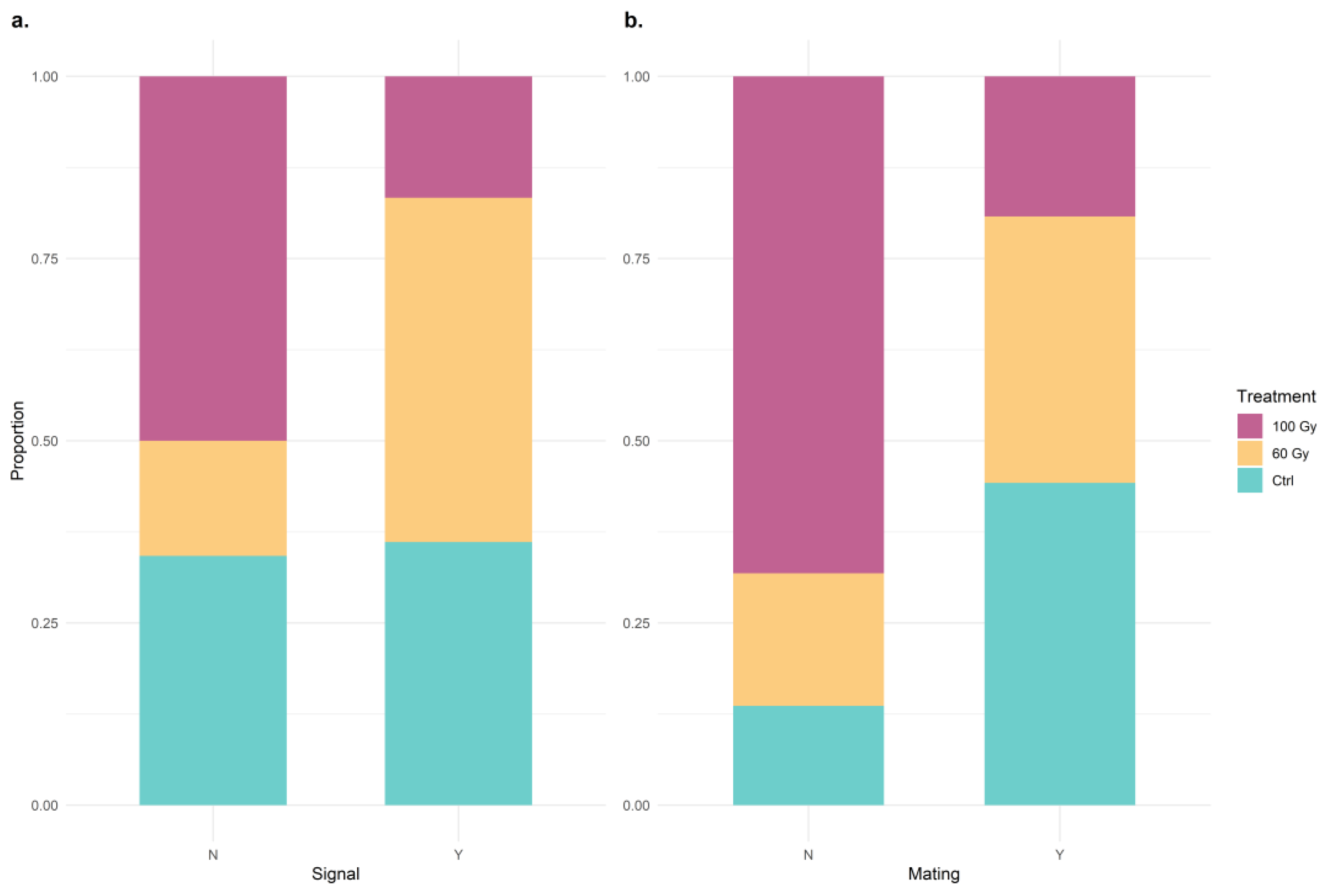
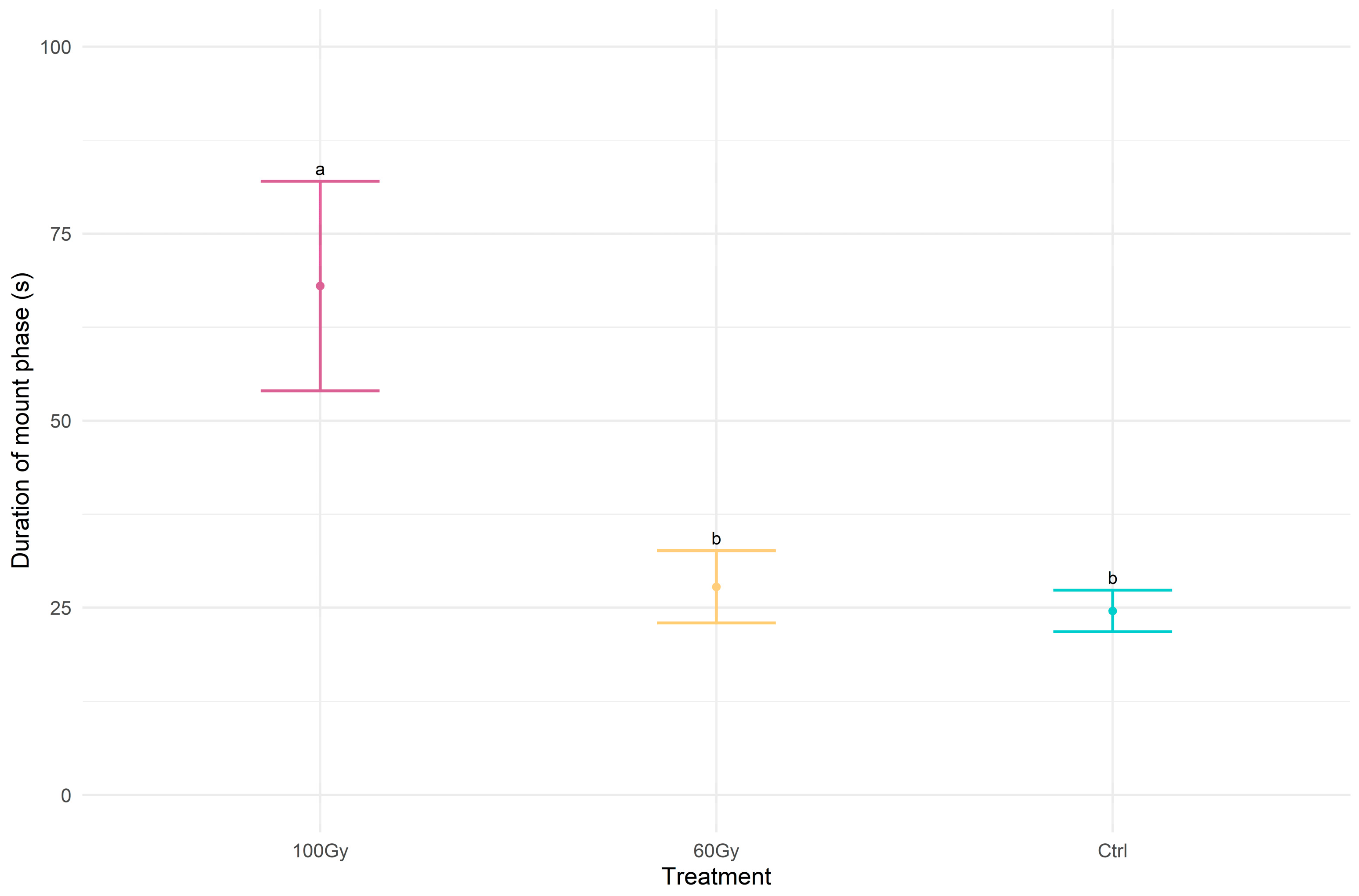
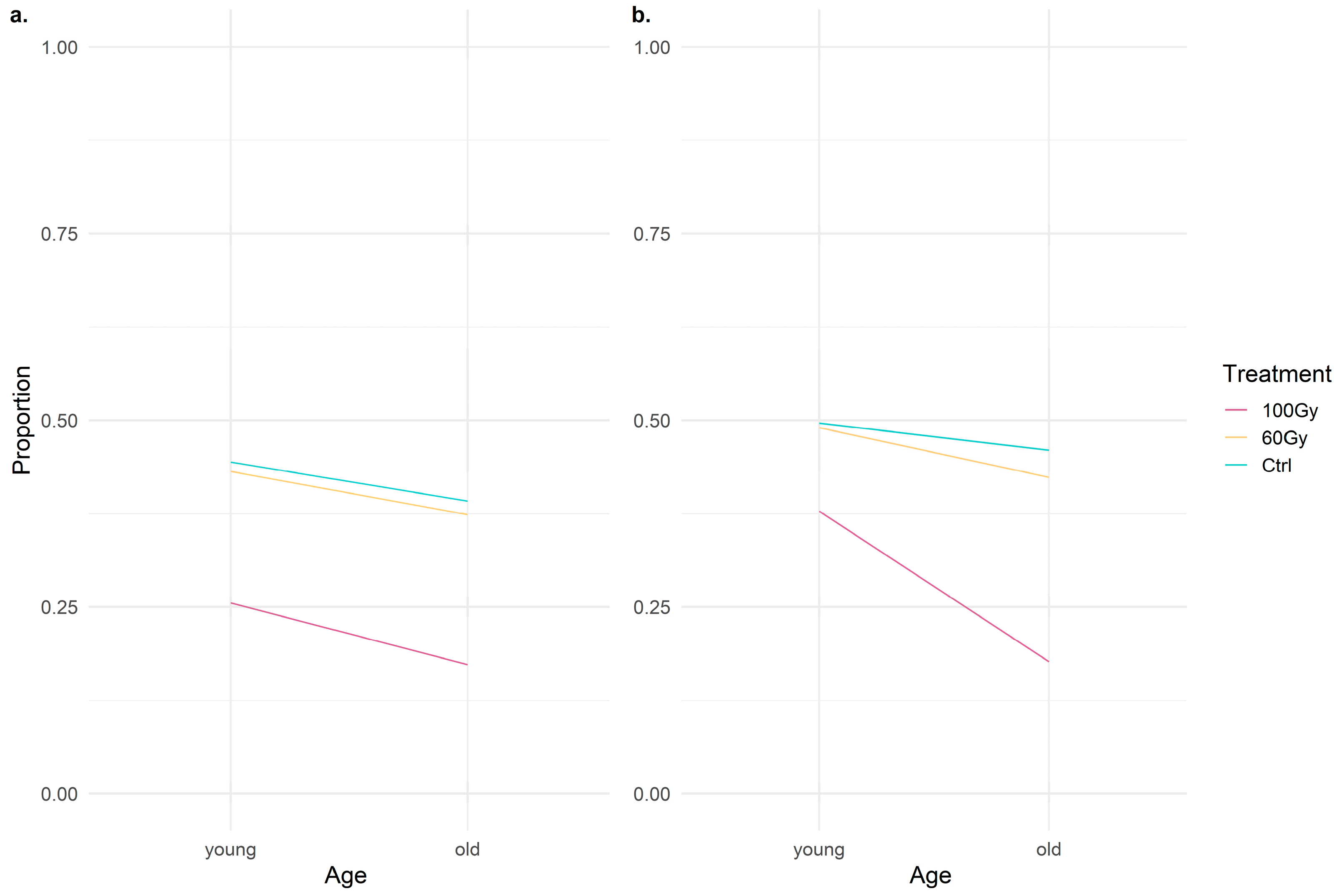
3.2. Pre-Copulatory and Mating Behavior
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ Warning on Invasive Alien Species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Sage, R.F. Global Change Biology: A Primer. Glob. Chang. Biol. 2020, 26, 3–30. [Google Scholar] [CrossRef] [PubMed]
- Singh Harvir, V.S. Malik Biology of Painted Bug (Bagrada cruciferarum). Indian J. Agric. Sci. 1993, 63, 672. [Google Scholar]
- Carvajal, M.A.; Alaniz, A.J.; Núñez-Hidalgo, I.; González-Césped, C. Spatial Global Assessment of the Pest Bagrada hilaris (Burmeister) (Heteroptera: Pentatomidae): Current and Future Scenarios: Global Assessment of the Pest Bagrada hilaris: Current and Future Scenarios. Pest. Manag. Sci. 2019, 75, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.A.; Palumbo, J.C.; Perring, T.M.; May, C. Bagrada hilaris (Hemiptera: Pentatomidae), An Invasive Stink Bug Attacking Cole Crops in the Southwestern United States. J. Integr. Pest Manag. 2013, 4, C1–C7. [Google Scholar] [CrossRef] [Green Version]
- Bundy, C.S.; Grasswitz, T.R.; Sutherland, C. First Report of the Invasive Stink Bug Bagrada hilaris (Burmeister) (Heteroptera: Pentatomidae) from New Mexico, with Notes on Its Biology. Southwest. Entomol. 2012, 37, 411–414. [Google Scholar] [CrossRef]
- Torres-Acosta, R.I.; Sánchez-Peña, S.R. Geographical Distribution of Bagrada hilaris (Hemiptera: Pentatomidae) in Mexico. J. Entomol. Sci. 2016, 51, 165–167. [Google Scholar] [CrossRef]
- Faúndez, E.I. From Agricultural to Household Pest: The Case of the Painted Bug Bagrada hilaris (Burmeister) (Heteroptera: Pentatomidae) in Chile. J. Med. Entomol. 2018, 55, 1365–1368. [Google Scholar] [CrossRef]
- Colazza, S.; Guarino, S.; Peri, E. Bagrada hilaris (Burmeister) (Heteroptera: Pentatomidae) a Pest of Capper in the Island of Pantelleria [Capparis spinosa L.; Sicily]. Inf. Fitopatol. 2004, 54, 30–34. [Google Scholar]
- Huang, T.-I.; Reed, D.A.; Perring, T.M.; Palumbo, J.C. Feeding Damage by Bagrada hilaris (Hemiptera: Pentatomidae) and Impact on Growth and Chlorophyll Content of Brassicaceous Plant Species. Arthropod-Plant Interact. 2014, 8, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Guarino, S.; Peri, E.; Colazza, S.; Luchi, N.; Michelozzi, M.; Loreto, F. Impact of the Invasive Painted Bug Bagrada hilaris on Physiological Traits of Its Host Brassica Oleracea Var Botrytis. Arthropod-Plant Interact. 2017, 11, 649–658. [Google Scholar] [CrossRef]
- Infantino, A.; Tomassoli, L.; Peri, E.; Colazza, S. Viruses, Fungi and Insect Pests Affecting Caper. Eur. J. Plant Sci. Biotechnol. 2007, 1, 170–179. [Google Scholar]
- Palumbo, J.C.; Perring, T.M.; Millar, J.G.; Reed, D.A. Biology, Ecology, and Management of an Invasive Stink Bug, Bagrada hilaris, in North America. Annu. Rev. Entomol. 2016, 61, 453–473. [Google Scholar] [CrossRef] [PubMed]
- Dhingra Swaran, S. Relative Toxicity of Some Important Insecticides with Particular Reference to Change in Susceptibility Level of Bagrada cruciferarum Kirk. during the Last Quarter Century. J. Entomol. Res. 1998, 22, 307–311. [Google Scholar]
- Guarino, S.; Peri, E.; Lo Bue, P.; La Pillo, A.; Colazza, S. Impiego Di Insetticidi Di Origine Vegetale per Il Contenimento Delle Popolazioni Di Bagrada hilaris Nei Cappereti Panteschi. Inf. Fitopatol. 2007, 7, 53–58. [Google Scholar]
- Hendrichs, J.; Bloem, K.; Hoch, G.; Carpenter, J.E.; Greany, P.; Robinson, A.S. Improving the Cost-Effectiveness, Trade and Safety of Biological Control for Agricultural Insect Pests Using Nuclear Techniques. Biocontrol Sci. Technol. 2009, 19, 3–22. [Google Scholar] [CrossRef]
- Klassen, W.; Curtis, C.F. History of the sterile insect technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 3–36. ISBN 9781003035572. [Google Scholar]
- Knipling, E.F. Possibilities of Insect Control or Eradication Through the Use of Sexually Sterile Males1. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Robinson, A.S. Genetic Basis of the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; CRC: Boca Raton, FL, USA, 2021. [Google Scholar]
- Lance, D.R.; McInnis, D.O. Biological basis of the sterile insect technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; p. 30. ISBN 9781003035572. [Google Scholar]
- Klassen, W.; Vreysen, M.J.B. Area-Wide Integrated Pest Management and the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; CRC Press: Boca Raton, FL, USA, 2021; pp. 75–112. ISBN 9781003035572. [Google Scholar]
- Kapranas, A.; Collatz, J.; Michaelakis, A.; Milonas, P. Review of the Role of Sterile Insect Technique within Biologically-Based Pest Control—An Appraisal of Existing Regulatory Frameworks. Entomol. Exp. Appl. 2022, 170, 385–393. [Google Scholar] [CrossRef]
- Cornelius, M.L.; Dieckhoff, C.; Hoelmer, K.A.; Olsen, R.T.; Weber, D.C.; Herlihy, M.V.; Talamas, E.J.; Vinyard, B.T.; Greenstone, M.H. Biological Control of Sentinel Egg Masses of the Exotic Invasive Stink Bug Halyomorpha halys (Stål) in Mid-Atlantic USA Ornamental Landscapes. Biol. Control 2016, 103, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Ganjisaffar, F.; Talamas, E.J.; Bon, M.-C.; Brown, B.V.; Gonzalez, L.; Perring, T.M. Trissolcus hyalinipennis Rajmohana & Narendran (Hymenoptera, Scelionidae), a Parasitoid of Bagrada hilaris (Burmeister) (Hemiptera, Pentatomidae), Emerges in North America. J. Hymenopt. Res. 2018, 65, 111–130. [Google Scholar] [CrossRef] [Green Version]
- Martel, G.; Augé, M.; Talamas, E.; Roche, M.; Smith, L.; Sforza, R.F.H. First Laboratory Evaluation of Gryon Gonikopalense (Hymenoptera: Scelionidae), as Potential Biological Control Agent of Bagrada hilaris (Hemiptera: Pentatomidae). Biol. Control 2019, 135, 48–56. [Google Scholar] [CrossRef]
- Power, N.; Ganjisaffar, F.; Perring, T.M. Evaluation of the Physiological Host Range for the Parasitoid Ooencyrtus mirus, a Potential Biocontrol Agent of Bagrada hilaris. Insects 2020, 11, 432. [Google Scholar] [CrossRef]
- Cayol, J.P. Changes in Sexual Behavior and Life History Traits of Tephritid Species Caused by Mass-Rearing Processes. In Fruit Flies (Tephritidae); CRC Press: Boca Raton, FL, USA, 2001; pp. 861–878. ISBN 978-0-429-12467-9. [Google Scholar]
- Cristofaro, M.; Sforza, R.F.H.; Roselli, G.; Paolini, A.; Cemmi, A.; Musmeci, S.; Anfora, G.; Mazzoni, V.; Grodowitz, M. Effects of Gamma Irradiation on the Fecundity, Fertility, and Longevity of the Invasive Stink Bug Pest Bagrada hilaris (Burmeister) (Hemiptera: Pentatomidae). Insects 2022, 13, 787. [Google Scholar] [CrossRef] [PubMed]
- Bakri, A.; Mehta, K.; Lance, D.R. Sterilizing Insects with Ionizing Radiation. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; CRC Press: Boca Raton, FL, USA, 2021; pp. 355–398. ISBN 978-1-00-303557-2. [Google Scholar]
- Wang, L.-M.; Li, N.; Zhang, M.; Tang, Q.; Lu, H.-Z.; Zhou, Q.-Y.; Niu, J.-X.; Xiao, L.; Peng, Z.-Y.; Zhang, C.; et al. The Sex Pheromone Heptacosane Enhances the Mating Competitiveness of Sterile Aedes aegypti Males. Parasites Vectors 2023, 16, 102. [Google Scholar] [CrossRef]
- Cocroft, R.B.; Rodríguez, R.L. The Behavioral Ecology of Insect Vibrational Communication. BioScience 2005, 55, 323. [Google Scholar] [CrossRef]
- Guarino, S.; De Pasquale, C.; Peri, E.; Alonzo, G.; Colazza, S. Role of Volatile and Contact Pheromones in the Mating Behaviour of Bagrada hilaris (Heteroptera: Pentatomidae). Eur. J. Entomol. 2008, 105, 613–617. [Google Scholar] [CrossRef] [Green Version]
- Scala, M.; Peccerillo, C.; Fouani, J.; Nieri, R.; Mazzoni, V.; Baser, N.; Verrastro, V.; Anfora, G. Characterization of Substrate-Borne Vibrational Communication of Bagrada hilaris. In Proceedings of the III International Conference on Biotremology, Piran, Slovenia, 19–22 September 2022; p. 37. [Google Scholar]
- Taylor, M.E.; Bundy, C.S.; McPherson, J.E. Unusual Ovipositional Behavior of the Stink Bug Bagrada hilaris (Hemiptera: Heteroptera: Pentatomidae). Ann. Entomol. Soc. Am. 2014, 107, 872–877. [Google Scholar] [CrossRef] [Green Version]
- Baccaro, S.; Cemmi, A.; Di Sarcina, I.; Ferrara, G. Gamma Irradiation Calliope Facility at ENEA–Casaccia Research Centre (Rome, Italy); Fusion and Technology for Nuclear Safety and Security Department Casaccia Research Centre, Ed.; 2019; p. 49. Available online: https://iris.enea.it/retrieve/dd11e37c-d730-5d97-e053-d805fe0a6f04/RT-2019-04-ENEA.pdf (accessed on 1 February 2023).
- Polajnar, J.; Maistrello, L.; Bertarella, A.; Mazzoni, V. Vibrational Communication of the Brown Marmorated Stink Bug (Halyomorpha halys): Vibrational Communication in Halyomorpha halys. Physiol. Entomol. 2016, 41, 249–259. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 2 February 2023).
- Burchett, W.W.; Ellis, A.R.; Harrar, S.W.; Bathke, A.C. Nonparametric Inference for Multivariate Data: The R Package Npmv. J. Stat. Soft. 2017, 76, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Dinno, A. Nonparametric Pairwise Multiple Comparisons in Independent Groups Using Dunn’s Test. Stata J. 2015, 15, 292–300. [Google Scholar] [CrossRef] [Green Version]
- Brooks, M.E.; Kristensen, K.; Benthem, K.J.; van Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. GlmmTMB Balances Speed and Flexibility Among Packages for Zero-Inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, Y.; Makio, I.; Genshiro, K. Akaike Information Criterion Statistics; no. 10.5555 1986: 26853; D. Reidel 81: Dordrecht, The Netherlands, 1994. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef] [Green Version]
- Parker, A.G.; Vreysen, M.J.B.; Bouyer, J.; Calkins, C.O. Sterile Insect Quality Control/Assurance. In Sterile Insect Technique; CRC Press: Boca Raton, FL, USA, 2021; pp. 399–440. ISBN 978-1-00-303557-2. [Google Scholar]
- Markow, T.A.; O’Grady, P.M. Evolutionary Genetics of Reproductive Behavior in Drosophila: Connecting the Dots. Annu. Rev. Genet. 2005, 39, 263–291. [Google Scholar] [CrossRef] [Green Version]
- Dahanukar, A.; Ray, A. Courtship, Aggression and Avoidance: Pheromones, Receptors and Neurons for Social Behaviors in Drosophila. Fly 2011, 5, 58–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO/IAEA/USDA. Product Quality Control for Sterile Mass-Reared and Released Tephritid Fruit Flies, Version 7.0; International Atomic Energy Agency: Vienna, Austria, 2019. [Google Scholar]
- Mainardi, C.E.; Peccerillo, C. Biotechnology and Biological Control Agency (BBCA) Onlus, Rome, Italy. Insects, 2023; to be submitted. [Google Scholar]
- Yamagishi, M.Y.; Tsubaki, Y. Sperm Competition in the Melon Fly, Bactrocera cucurbitae (Diptera: Tephritidae): Effects of Sperm ?Longevity? On Sperm Precedence. J. Insect Behav. 1992, 5, 599–608. [Google Scholar] [CrossRef]
- He, N.; Tsubaki, Y.; Itou, I.K.; Miyata, A. Gamma Radiation Effects on Reproductive Potential and Sperm Use Pattern in Pseudaletia separata (Lepidoptera: No Ctuidae ). J. Econ. Entomol. 1995, 88, 1626–1630. [Google Scholar] [CrossRef]
- Arif, M.A.; Guarino, S.; Colazza, S.; Peri, E. The Role of (E)-2-Octenyl Acetate as a Pheromone of Bagrada hilaris (Burmeister): Laboratory and Field Evaluation. Insects 2020, 11, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suckling, D.; Cristofaro, M.; Roselli, G.; Levy, M.; Cemmi, A.; Mazzoni, V.; Stringer, L.; Zeni, V.; Ioriatti, C.; Anfora, G. The Competitive Mating of Irradiated Brown Marmorated Stink Bugs, Halyomorpha halys, for the Sterile Insect Technique. Insects 2019, 10, 411. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | Model Synthesis | AIC | BIC | LogLik | Dev. | df |
|---|---|---|---|---|---|---|
| (mating) | behavior1 ~ treatment + age+ (1|day period) | 115.3 | 127.7 | −52.6 | 105.3 | 84 |
| (m.sig) | behavior2 ~ treatment + age+ Mount signals + (1|day period) | 79.0 | 92.8 | −33.5 | 67.0 | 68 |
| B | Fixed Effects | Estimate | Std. Error | Z Value | Pr (>|z|) | |
| (mating) | (Intercept) treatment 100 Gy | 0.7850 −1.8429 | 0.4512 0.5721 | 1.740 −3.222 | 0.08188 0.00127 ** | |
| (m.sig) | (Intercept) Treatment 100 Gy Age Young | 1.7371 −2.7747 1.6420 | 0.7383 0.8348 2.366 | 2.353 −3.324 2.366 | 0.018627 * 0.000888 *** 0.017961 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peccerillo, C.; Mainardi, C.E.; Nieri, R.; Fouani, J.M.; Cemmi, A.; Cristofaro, M.; Anfora, G.; Mazzoni, V. The Effect of the Sterile Insect Technique on Vibrational Communication: The Case of Bagrada hilaris (Hemiptera: Pentatomidae). Insects 2023, 14, 353. https://doi.org/10.3390/insects14040353
Peccerillo C, Mainardi CE, Nieri R, Fouani JM, Cemmi A, Cristofaro M, Anfora G, Mazzoni V. The Effect of the Sterile Insect Technique on Vibrational Communication: The Case of Bagrada hilaris (Hemiptera: Pentatomidae). Insects. 2023; 14(4):353. https://doi.org/10.3390/insects14040353
Chicago/Turabian StylePeccerillo, Chiara, Chiara Elvira Mainardi, Rachele Nieri, Jalal Melhem Fouani, Alessia Cemmi, Massimo Cristofaro, Gianfranco Anfora, and Valerio Mazzoni. 2023. "The Effect of the Sterile Insect Technique on Vibrational Communication: The Case of Bagrada hilaris (Hemiptera: Pentatomidae)" Insects 14, no. 4: 353. https://doi.org/10.3390/insects14040353