
Banker Plant Bonuses? The Benefits and Risks of Including Brassicas in Field Margins to Promote Conservation Biocontrol of Specialist Pests in Oilseed Rape

Abstract
:Simple Summary
Abstract
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pest | Parasitoids | Notes and References |
|---|---|---|
| Pollen beetle (Brassicogethes aeneus) | Tersilochus heterocerus (Thomson) 1,a,† Phradis interstitialis (Thomsson) 1,a,† Phradis morionellus (Holmgren) 1,a,† | Diospilus capito (Nees) 2,a,† and Blacus nigricornis Haeselbarth 2,a,† are widespread but less common and therefore of minor importance for biocontrol [37] |
| Cabbage seed weevil (Ceutorhynchus obstrictus) | Trichomalus perfectus Walker 3,b,† Stenomalina gracilis Walker 3,b,† Mesopolobus morys Walker 3,b,† | [37,39] |
| Brassica pod midge (Dasineura brassicae) | Platygaster subuliformis Kieffer 4,a,‡ Omphale clypealis Haliday 5,a,† | [37,40] |
| Cabbage stem weevil (Ceutorhynchus pallidactylus) | Tersilochus obscurator Aubert 1 | [41] |
2. Materials and Methods
2.1. Plant Selection and Experimental Design
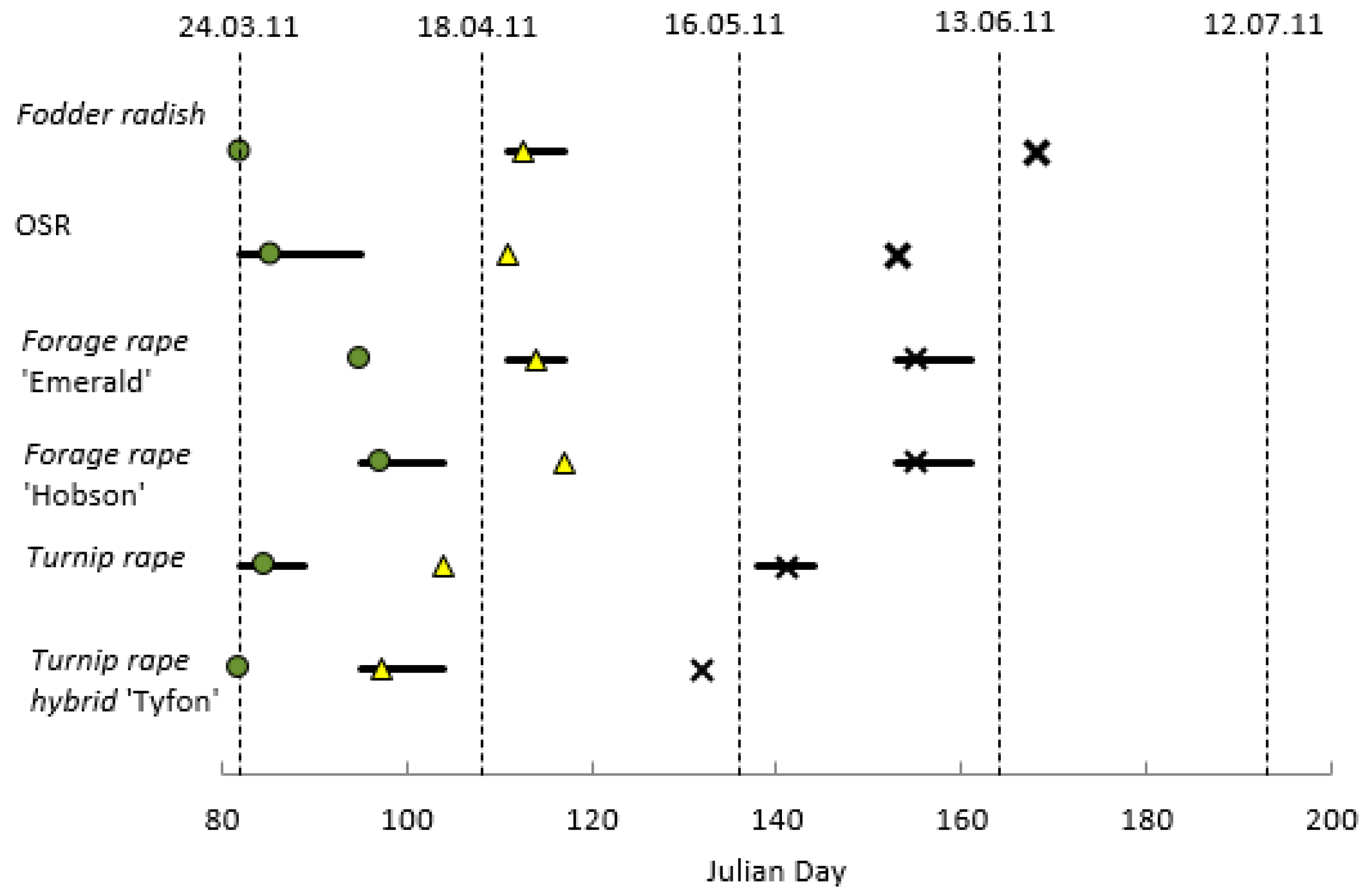
2.2. Plant Growth Stage Assessments
2.3. Suction Sampling of Adult Insects
2.4. Pod Infestation by the Pest Larvae of Cabbage Seed Weevil and Brassica Pod Midge and Parasitism of Cabbage Seed Weevil Larvae
2.5. Trapping of Emerging Insects
2.6. Statistical Analyses
3. Results
3.1. Growth Stage Assessments
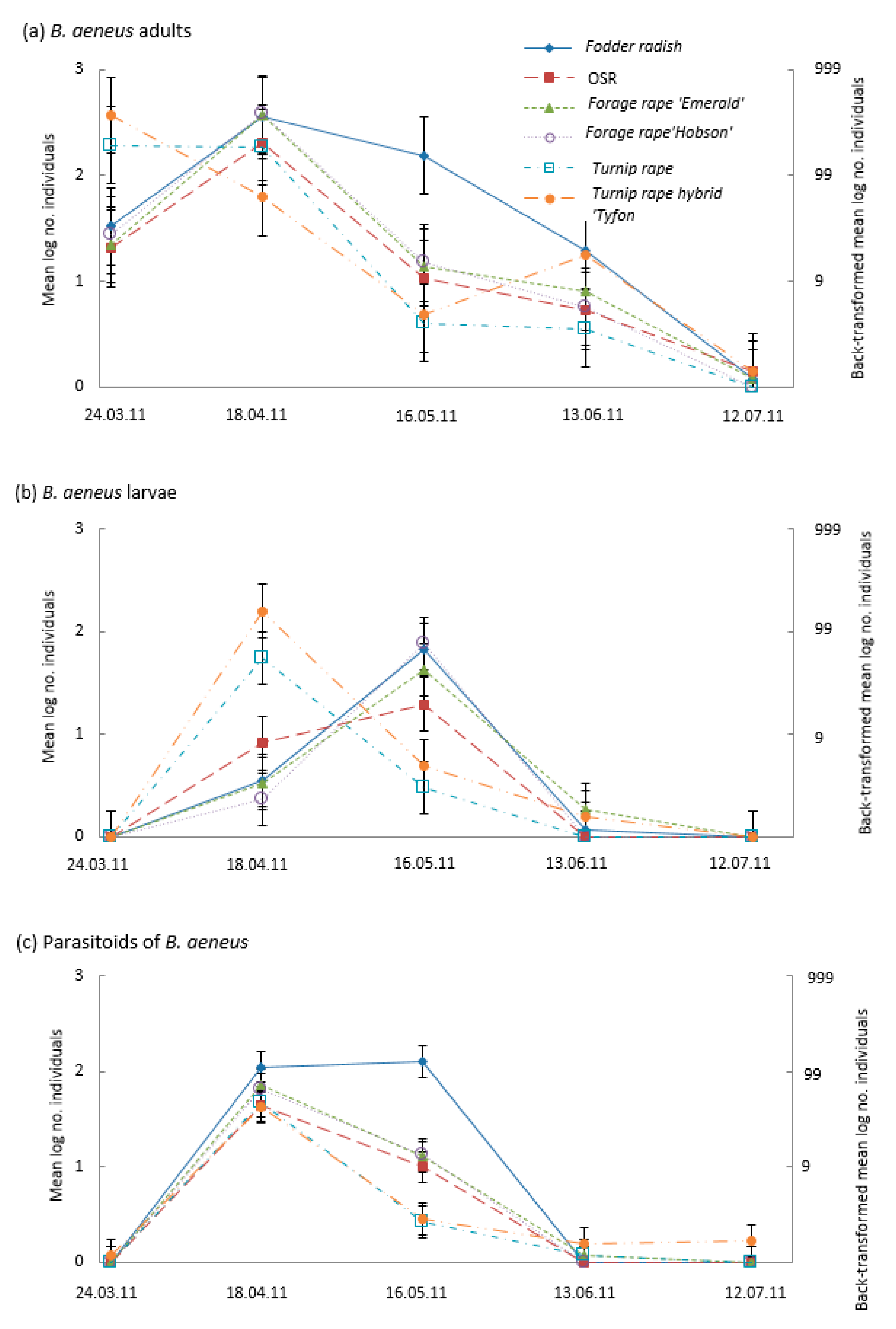
3.2. Suction Sampling of Adult Insects
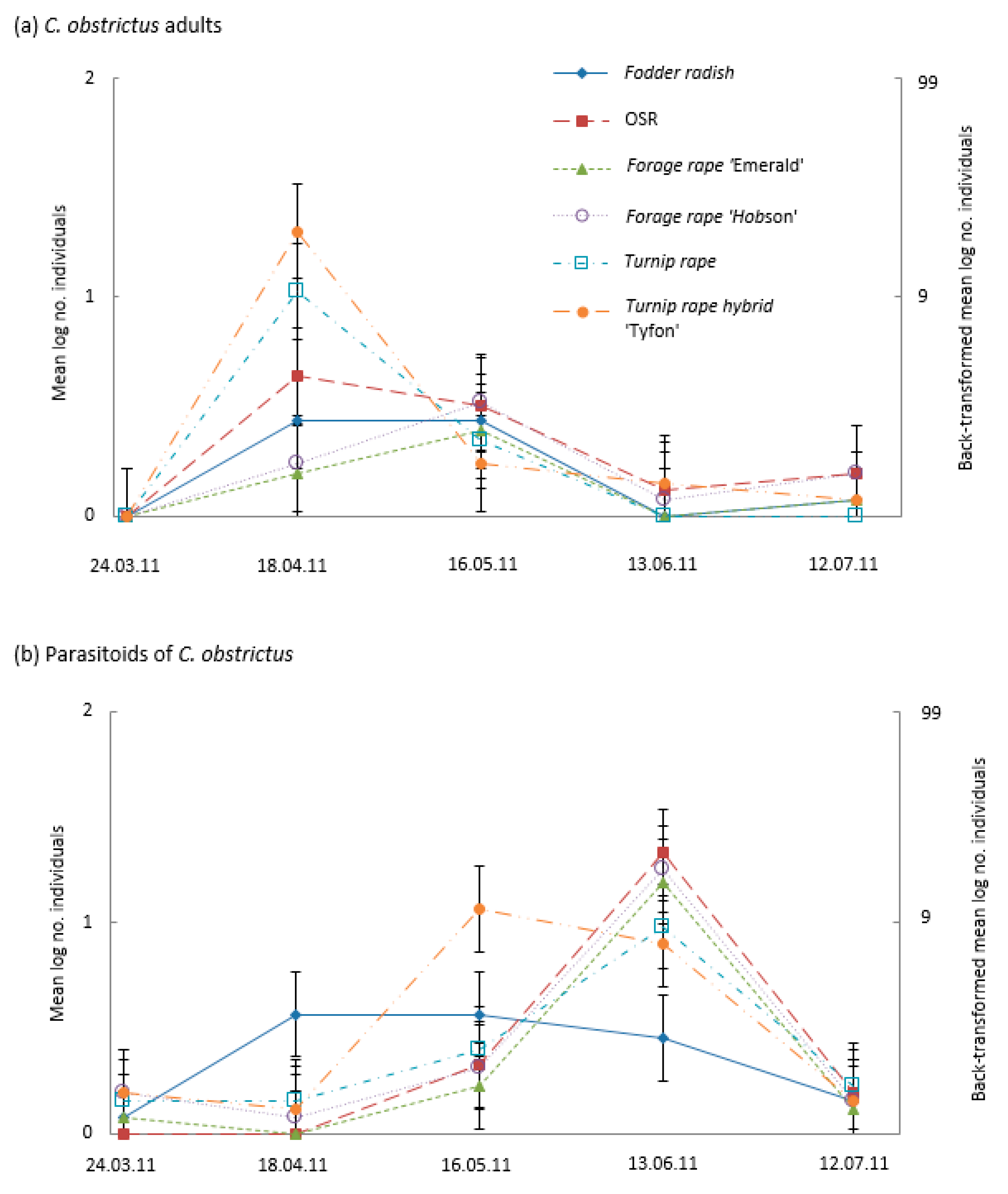
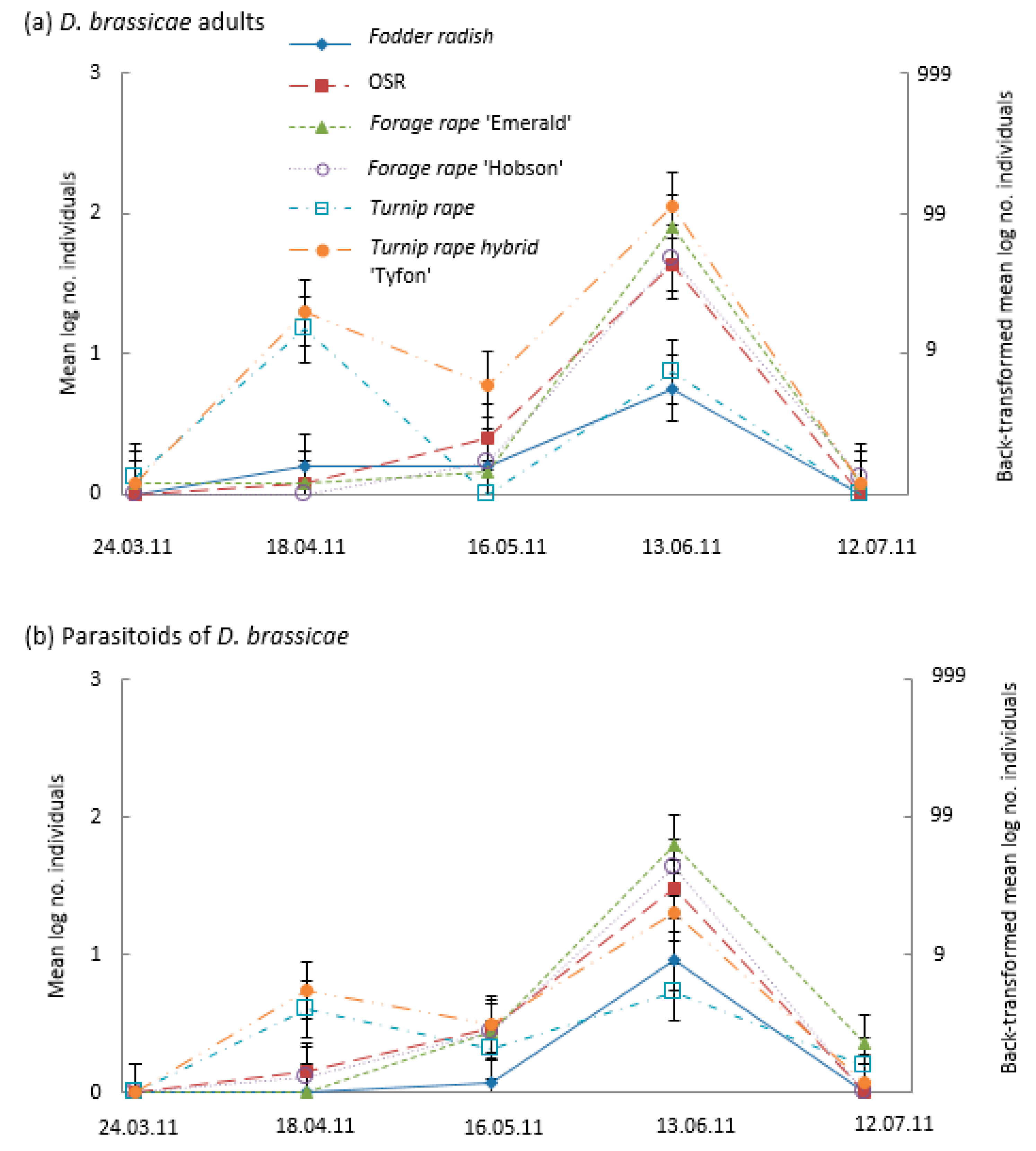
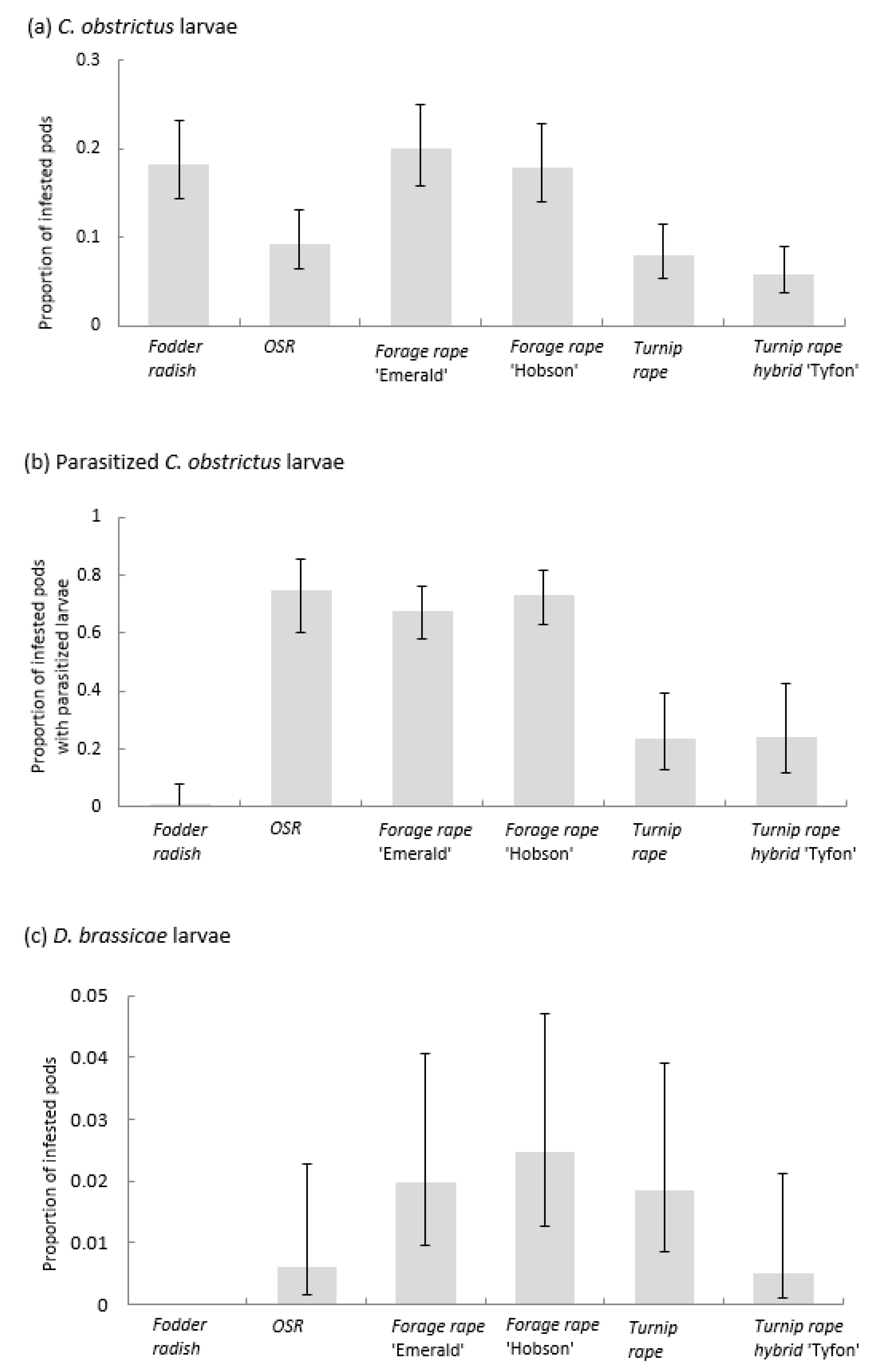
3.3. Pod Infestation by the Pest Larvae of the Cabbage Seed Weevil (Ceutorhynchus obstrictus), the Brassica Pod Midge (Dasineura brassicae), and Parasitism of Cabbage Seed Weevil Larvae
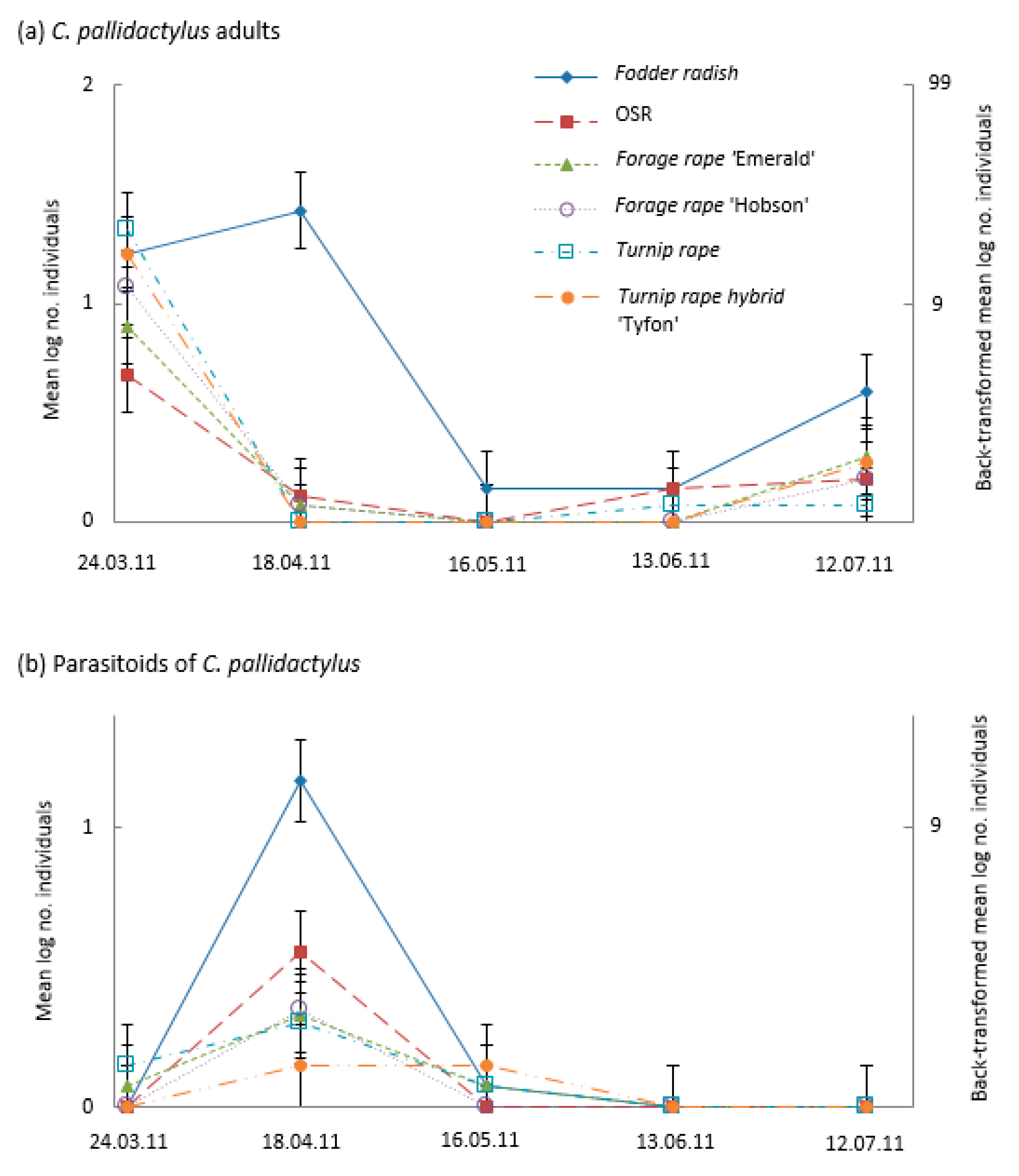
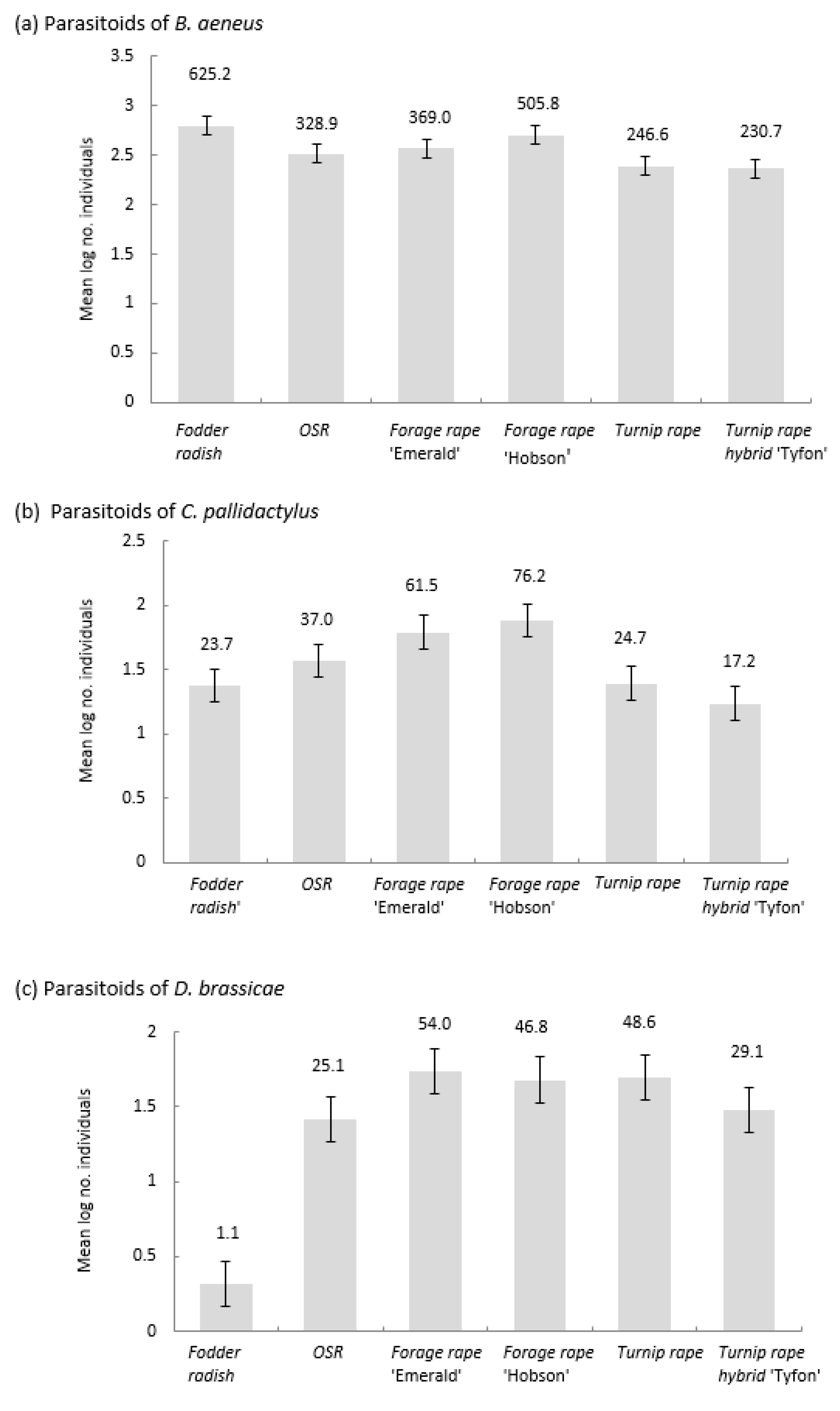
3.4. Parasitoid Emergence from Emergence Traps
4. Discussion
4.1. Banker Plants for Parasitoids of Pollen Beetles
4.2. Banker Plants for Parasitoids of Weevil Pests
4.3. Banker Plants for Parasitoids of Brassica Pod Midge
4.4. Banker Plants as Trap Crops for Pests of Oilseed Rape and Implications for Parasitoids
4.5. Selection of Banker Plants for Pest Management in Oilseed Rape
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape Perspectives on Agricultural Intensification and Biodiversity–Ecosystem Service Management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Robinson, R.A.; Sutherland, W.J. Post-war Changes in Arable Farming and Biodiversity in Great Britain. J. Appl. Ecol. 2002, 39, 157–176. [Google Scholar] [CrossRef] [Green Version]
- Raven, P.H.; Wagner, D.L. Agricultural Intensification and Climate Change Are Rapidly Decreasing Insect Biodiversity. Proc. Natl. Acad. Sci. USA 2021, 118, e2002548117. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide Decline of the Entomofauna: A Review of Its Drivers. Biol. Conservl. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Bianchi, F.J.; Booij, C.; Tscharntke, T. Sustainable Pest Regulation in Agricultural Landscapes: A Review on Landscape Composition, Biodiversity and Natural Pest Control. Proc. R Soc. B 2006, 273, 1715–1727. [Google Scholar] [CrossRef] [Green Version]
- Rusch, A.; Valantin-Morison, M.; Sarthou, J.-P.; Roger-Estrade, J. Biological Control of Insect Pests in Agroecosystems: Effects of Crop Management, Farming Systems, and Seminatural Habitats at the Landscape Scale: A Review. Adv. Agron. 2010, 109, 219–259. [Google Scholar]
- Sarthou, J.-P.; Badoz, A.; Vaissière, B.; Chevallier, A.; Rusch, A. Local More than Landscape Parameters Structure Natural Enemy Communities during Their Overwintering in Semi-Natural Habitats. Agric. Ecosyst. Environ. 2014, 194, 17–28. [Google Scholar] [CrossRef]
- Holland, J.M.; Bianchi, F.J.; Entling, M.H.; Moonen, A.; Smith, B.M.; Jeanneret, P. Structure, Function and Management of Semi-natural Habitats for Conservation Biological Control: A Review of European Studies. Pest. Manag. Sci. 2016, 72, 1638–1651. [Google Scholar] [CrossRef]
- Sutter, L.; Amato, M.; Jeanneret, P.; Albrecht, M. Overwintering of Pollen Beetles and Their Predators in Oilseed Rape and Semi-Natural Habitats. Agric. Ecosyst. Environ. 2018, 265, 275–281. [Google Scholar] [CrossRef]
- Wäckers, F.L.; van Rijn, P.C.J.; Bruin, J. Plant-Provided Food for Carnivorous Insects: A Protective Mutualism and Its Applications; Cambridge University Press: Cambridge, UK, 2005; ISBN 1-139-44354-2. [Google Scholar]
- Timberlake, T.P.; Vaughan, I.P.; Memmott, J. Phenology of Farmland Floral Resources Reveals Seasonal Gaps in Nectar Availability for Bumblebees. J. Appl. Ecol. 2019, 56, 1585–1596. [Google Scholar] [CrossRef]
- Denys, C.; Tscharntke, T. Plant-Insect Communities and Predator-Prey Ratios in Field Margin Strips, Adjacent Crop Fields, and Fallows. Oecologia 2002, 130, 315–324. [Google Scholar] [CrossRef]
- Wissinger, S.A. Cyclic Colonization in Predictably Ephemeral Habitats: A Template for Biological Control in Annual Crop Systems. Biol. Control. 1997, 10, 4–15. [Google Scholar] [CrossRef]
- Gladbach, D.J.; Holzschuh, A.; Scherber, C.; Thies, C.; Dormann, C.F.; Tscharntke, T. Crop–Noncrop Spillover: Arable Fields Affect Trophic Interactions on Wild Plants in Surrounding Habitats. Oecologia 2011, 166, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Cole, L.J.; Kleijn, D.; Dicks, L.V.; Stout, J.C.; Potts, S.G.; Albrecht, M.; Balzan, M.V.; Bartomeus, I.; Bebeli, P.J.; Bevk, D.; et al. A Critical Analysis of the Potential for EU Common Agricultural Policy Measures to Support Wild Pollinators on Farmland. J. Appl. Ecol. 2020, 57, 681–694. [Google Scholar] [CrossRef] [PubMed]
- European Commission Report from the Commission to the European Parliament and the Council on the Implementation of the Ecological Focus Area Obligation under the Direct Payment Scheme. 2017. Available online: https://ec.europa.eu/smart-regulation/roadmaps/docs/2017_agri_002_evaluation_greening_en.pdf#:~:text=There%20is%20also%20a%20legal%20obligation%20in%20Article,relevant%20information%20for%20the%20ecological%20focus%20area%20report (accessed on 30 March 2023).
- Carvell, C.; Westrich, P.; Meek, W.R.; Pywell, R.F.; Nowakowski, M. Assessing the Value of Annual and Perennial Forage Mixtures for Bumblebees by Direct Observation and Pollen Analysis. Apidologie 2006, 37, 326–340. [Google Scholar] [CrossRef] [Green Version]
- Haaland, C.; Naisbit, R.E.; Bersier, L.-F. Sown Wildflower Strips for Insect Conservation: A Review. Insect. Conserv. Diver. 2011, 4, 60–80. [Google Scholar] [CrossRef]
- Vickery, J.; Feber, R.; Fuller, R. Arable Field Margins Managed for Biodiversity Conservation: A Review of Food Resource Provision for Farmland Birds. Agric. Ecosyst. Environ. 2009, 133, 1–13. [Google Scholar] [CrossRef]
- Tougeron, K.; Couthouis, E.; Marrec, R.; Barascou, L.; Baudry, J.; Boussard, H.; Burel, F.; Couty, A.; Doury, G.; Francis, C. Multi-Scale Approach to Biodiversity Proxies of Biological Control Service in European Farmlands. Sci. Total. Environ. 2022, 822, 153569. [Google Scholar] [CrossRef]
- Holland, J.; Oaten, H.; Moreby, S.; Birkett, T.; Simper, J.; Southway, S.; Smith, B. Agri-Environment Scheme Enhancing Ecosystem Services: A Demonstration of Improved Biological Control in Cereal Crops. Agric. Ecosyst. Environ. 2012, 155, 147–152. [Google Scholar] [CrossRef]
- Olson, D.M.; Wäckers, F.L. Management of Field Margins to Maximize Multiple Ecological Services. J. Appl. Ecol. 2007, 44, 13–22. [Google Scholar] [CrossRef]
- Winkler, K.; Wackers, F.; Termorshuizen, A.; van Lenteren, J. Assessing Risks and Benefits of Floral Supplements in Conservation Biological Control. BioControl 2010, 55, 719–727. [Google Scholar] [CrossRef] [Green Version]
- Frank, S.D. Biological Control of Arthropod Pests Using Banker Plant Systems: Past Progress and Future Directions. Biol. Control. 2010, 52, 8–16. [Google Scholar] [CrossRef]
- Olson, D.M.; Webster, T.M.; Scully, B.T.; Strickland, T.C.; Davis, R.F.; Knoll, J.E.; Anderson, W.F. Use of Winter Legumes as Banker Plants for Beneficial Insect Species in a Sorghum and Cotton Rotation System. J. Entomol. Sci. 2012, 47, 350–359. [Google Scholar] [CrossRef]
- George, D.R.; Croft, P.; Wakefield, M.; Wäckers, F.L. Perennial Field Margins for Functional Biodiversity in UK Vegetable Rotation Schemes: Establishment and Resource Provision. In Proceedings of the IOBC/WPRS Working Group Landscape Management for Functional Biodiversity, Llaida, Spain, 7–10 May 2012; IOBC-WPRS Bulletin, 2012. Volume 75, pp. 77–81. [Google Scholar]
- Rand, T.A.; Louda, S.M. Spillover of Agriculturally Subsidised Predators as a Potential Threat to Native Herbivores in Fragmented Landscapes. Conserv. Biol. 2006, 20, 1720–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelton, A.; Badenes-Perez, F. Concepts and Applications of Trap Cropping in Pest Management. Annu. Rev. Entomol. 2006, 51, 285–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, S.M.; Rasmussen, H.; Birkett, M.; Murray, D.; Pye, B.; Watts, N.; Williams, I. Behavioural and Chemical Ecology Underlying the Success of Turnip Rape (Brassica Rapa) Trap Crops in Protecting Oilseed Rape (Brassica Napus) from the Pollen Beetle (Meligethes Aeneus). Arthropod-Plant Interact. 2007, 1, 57–67. [Google Scholar] [CrossRef]
- Veromann, E.; Metspalu, L.; Williams, I.; Hiiesaar, K.; Mand, M.; Kaasik, R.; Kovacs, G.; Jogar, K.; Svilponis, E.; Kivimagi, I.; et al. Relative Attractiveness of Brassica napus, Brassica nigra, Eruca sativa and Raphanus sativus for Pollen Beetle (Meligethes aeneus) and Their Potential for Use in Trap Cropping. Arthropod-Plant Interact. 2012, 6, 385–394. [Google Scholar] [CrossRef]
- Kovács, G.; Kaasik, R.; Kaart, T.; Metspalu, L.; Luik, A.; Veromann, E. In Search of Secondary Plants to Enhance the Efficiency of Cabbage Seed Weevil Management. BioControl 2017, 62, 29–38. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#home (accessed on 30 March 2023).
- Ortega-Ramos, P.; Cook, S.; Mauchline, A.L. How Contradictory EU Policies Led to the Development of a Pest: The Story of Oilseed Rape and the Cabbage Stem Flea Beetle. GCB Bioenergy 2022, 14, 258–266. [Google Scholar] [CrossRef]
- Williams, I.H. The Major Insect Pests of Oilseed Rape in Europe and Their Management: An Overview. In Biocontrol-Based Integrated Management of Oilseed Rape Pests; Williams, I.H., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 1–43. ISBN 978-90-481-3982-8. e-ISBN 978-90-481-3983-5. [Google Scholar]
- Nilsson, C. Impact of Soil Tillage on Parasitoids of Oilseed Rape Pests. In Biocontrol-Based Integrated Management of Oilseed Rape Pests; Williams, I.H., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 305–311. ISBN 978-90-481-3982-8. e-ISBN 978-90-481-3983-5. [Google Scholar]
- Ferguson, A.; Holdgate, R.; Mason, N.; Clark, S.; Williams, I. Non Inversion Tillage to Conserve Functional Biodiversity for Biocontrol of Oilseed Rape Pests. In Proc. XVI Int Plant Protection Congress; British Crop Protection Council: Cambridge, UK, 2007; Volume 18, pp. 818–819. [Google Scholar]
- Ulber, B.; Williams, I.H.; Klukowski, Z.; Luik, A.; Nilsson, C. Parasitoids of Oilseed Rape Pests in Europe: Key Species for Conservation Biocontrol. In Biocontrol-Based Integrated Management of Oilseed Rape Pests; Williams, I.H., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 45–76. ISBN 978-90-481-3982-8. e-ISBN 978-90-481-3983-5. [Google Scholar]
- Thies, C.; Tscharntke, T. Biological Rape Pest Control in Spatio-Temporally Changing Landscapes. In Biocontrol-Based Integrated Management of Oilseed Rape Pests; Williams, I.H., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 273–284. ISBN 978-90-481-3982-8. eISBN 978-90-481-3983-5. [Google Scholar]
- Williams, I.H. Parasitoids of Cabbage Seed Weevil. In Biocontrol of Oilseed Rape Pests; Blackwell Science Ltd.: Oxford, UK, 2003; pp. 97–112. ISBN 10: 0632054271. eISBN 13: 9780632054275. [Google Scholar]
- Williams, I.H. Parasitoids of Brassica Pod Midge. In Biocontrol of Oilseed Rape Pests; Blackwell Science Ltd.: Oxford, UK, 2003; p. 113. ISBN 10: 0632054271. eISBN 13: 9780632054275. [Google Scholar]
- Ulber, B. Parasitoids of Ceutorhynchid Stem Weevils. In Biocontrol of Oilseed Rape Pests; Blackwell Science Ltd.: Oxford, UK, 2003; pp. 87–95. ISBN 10: 0632054271. eISBN 13: 9780632054275. [Google Scholar]
- Lancashire, P.D.; Bleiholder, H.; Boom, T.V.D.; Langelüddeke, P.; Stauss, R.; Weber, E.; Witzenberger, A. A Uniform Decimal Code for Growth Stages of Crops and Weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Arnold, A. Insect Suction Sampling without Nets, Bags or Filters. Crop. Prot. 1994, 13, 73–76. [Google Scholar] [CrossRef]
- Ferguson, A.W.; Williams, I.H.; Castle, L.M.; Skellern, M. Key Parasitoids of the Pests of Oilseed Rape in Europe: A Guide to Their Identification. In Biocontrol-Based Integrated Management of Oilseed Rape Pests; Williams, I.H., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 77–114. ISBN 978-90-481-3982-8. e-ISBN 978-90-481-3983-5. [Google Scholar]
- Ferguson, A.W.; Klukowski, Z.; Walczak, B.; Clark, S.J.; Mugglestone, M.A.; Perry, J.N.; Williams, I.H. Spatial Distribution of Pest Insects in Oilseed Rape: Implications for Integrated Pest Management. Agric. Ecosyst. Environ. 2003, 95, 509–521. [Google Scholar] [CrossRef]
- Tschumi, M.; Albrecht, M.; Bärtschi, C.; Collatz, J.; Entling, M.H.; Jacot, K. Perennial, Species-Rich Wildflower Strips Enhance Pest Control and Crop Yield. Agric. Ecosyst. Environ. 2016, 220, 97–103. [Google Scholar] [CrossRef]
- Hatt, S.; Lopes, T.; Boeraeve, F.; Chen, J.; Francis, F. Pest Regulation and Support of Natural Enemies in Agriculture: Experimental Evidence of within Field Wildflower Strips. Ecol. Eng. 2017, 98, 240–245. [Google Scholar] [CrossRef]
- Sutter, L.; Albrecht, M.; Jeanneret, P. Landscape Greening and Local Creation of Wildflower Strips and Hedgerows Promote Multiple Ecosystem Services. J. Appl. Ecol. 2018, 55, 612–620. [Google Scholar] [CrossRef]
- Hatt, S.; Francis, F.; Xu, Q.; Wang, S.; Osawa, N. Perennial Flowering Strips for Conservation Biological Control of Insect Pests: From Picking and Mixing Flowers to Tailored Functional Diversity. In Integrative Biological Control: Ecostacking for Enhanced Ecosystem Services; Gao, Y., Hokkanen, H., Menzler-Hokkanen, I., Eds.; Integrative Biological Control. Progress in Biological Control; Springer: Dordrecht, The Netherlands, 2020; Volume 20, pp. 57–71. [Google Scholar] [CrossRef]
- Lavandero, B.; Wratten, S.D.; Didham, R.K.; Gurr, G. Increasing Floral Diversity for Selective Enhancement of Biological Control Agents: A Double-Edged Sward? Basic. Appl. Ecol. 2006, 7, 236–243. [Google Scholar] [CrossRef]
- Frearson, D.J.; Ferguson, A.W.; Campbell, J.M.; Williams, I.H. The Spatial Dynamics of Pollen Beetles in Relation to Inflorescence Growth Stage of Oilseed Rape: Implications for Trap Crop Strategies. Entomol. Exp. Et Appl. 2005, 116, 21–29. [Google Scholar] [CrossRef]
- Jönsson, M.; Lindkvist, A.; Anderson, P. Behavioural Responses in Three Ichneumonid Pollen Beetle Parasitoids to Volatiles Emitted from Different Phenological Stages of Oilseed Rape. Entomol. Exp. Et Appl. 2005, 115, 363–369. [Google Scholar] [CrossRef]
- Hopkins, R.J.; Ekbom, B. The Pollen Beetle, Meligethes aeneus, Changes Egg Production Rate to Match Host Quality. Oecologia 1999, 120, 274–278. [Google Scholar] [CrossRef]
- Ekbom, B.; Popov, S.Y. Host Plant Affects Pollen Beetle (Meligethes aeneus) Egg Size. Physiol. Entomol. 2004, 29, 118–122. [Google Scholar] [CrossRef]
- Billqvist, A.; Ekbom, B. Effects of Host Plant Species on the Interaction between the Parasitic Wasp Diospilus capito and Pollen Beetles (Meligethes spp.). Agric. For. Entomol. 2001, 3, 147–152. [Google Scholar] [CrossRef]
- Åhman, I. Wild and Cultivated Crucifers as Hosts for Dasineura brassicae Winn.(Dipt., Cecidomyiidae). J. Appl. Entomol. 1988, 105, 420–424. [Google Scholar] [CrossRef]
- Åhman, I. Oviposition in Dasineura Brassicae Winn. (Diptera, Cecidomyiidae). Adaptive, Mechanistic and Applied Aspects. Växtskyddsrapporter. Avhandlingar, vol 9. 1986 Sveriges Lantbruksuniversitet, Uppsala. 1986. Available online: https://agris.fao.org/agris-search/search.do?recordID=FR880480488 (accessed on 30 March 2023).
- Cook, S.; Smart, L.; Martin, J.; Murray, D.; Watts, N.; Williams, I. Exploitation of Host Plant Preferences in Pest Management Strategies for Oilseed Rape (Brassica napus). Entomol. Exp. Appl. 2006, 119, 221–229. [Google Scholar] [CrossRef]
- Vinatier, F.; Gosme, M.; Valantin-Morison, M. A Tool for Testing Integrated Pest Management Strategies on a Tritrophic System Involving Pollen Beetle, Its Parasitoid and Oilseed Rape at the Landscape Scale. Landscape Ecol. 2012, 27, 1421–1433. [Google Scholar] [CrossRef]
- Nilsson, C. Parasitoids of Pollen Beetles. In Biocontrol-Based Integrated Management of Oilseed Rape Pests; Blackwell Science Ltd.: Oxford, UK, 2003; pp. 73–86. ISBN 10: 0632054271. eISBN 13: 9780632054275. [Google Scholar]
- Osbourne, P. Observations on the Natural Enemies of Meligethes aeneus (F.) and viridescens (F.) [Coleoptera: Nitidulae]. Parasitology 1960, 50, 91–100. [Google Scholar] [CrossRef]
- Nitzsche, O.; Ulber, B. Influence of Different Tillage Treatments Following the Harvest of Oilseed Rape on the Mortality of Pollen Beetles (Meligethes spp.) Parasitoids. Z. Für Pflanzenkrankh. Und Pflanzenschutz 1998, 105, 417–421. [Google Scholar]
- Barari, H.; Cook, S.M.; Clark, S.J.; Williams, I.H. Effect of a Turnip Rape (Brassica Rapa) Trap Crop on Stem-Mining Pests and Their Parasitoids in Winter Oilseed Rape (Brassica Napus). Biocontrol 2005, 50, 69–86. [Google Scholar] [CrossRef]
- Kovács, G.; Kaasik, R.; Metspalu, L.; Williams, I.H.; Luik, A.; Veromann, E. Could Brassica Rapa, Brassica iuncea and Sinapis alba Facilitate the Control of the Cabbage Seed Weevil in Oilseed Rape Crops? Biol. Control. 2013, 65, 124–129. [Google Scholar] [CrossRef]
- Williams, I.; Martin, A.; Kelm, M. The Phenology of the Emergence of Brassica Pod Midge (Dasineura brassicae Winn) and Its Infestation of Winter Oilseed Rape (Brassica napus L.). J. Agr Sci 1987, 108, 579–589. [Google Scholar] [CrossRef]
- Buhl, C. Beobachtungen Uber Vermehrtes Schadauftreten Der Kohlschotenmucke (Dasineura brassicae Winn.) an Raps Und Rubsen in Schleswig-Holstein. Nachr Dtsch. Pflanzenschutz Stuttg. 1960, 12, 1–6. [Google Scholar]
- Czajkowska, M. Badania Nad Pryszczarkiem Kapustnikeiem (Dasyneura brassicae Winn.) I. Biologia i Ekologia. Rocz. Nauk. Rol. Ser. E 1987, 8, 115–131. [Google Scholar]
- OP2: Wild Bird Seed Mixture. Available online: https://www.gov.uk/countryside-stewardship-grants/wild-bird-seed-mixture-op2 (accessed on 30 March 2023).
- AB1: Nectar Flower Mix. Available online: https://www.gov.uk/countryside-stewardship-grants/nectar-flower-mix-ab1 (accessed on 30 March 2023).







| Fodder Radish | OSR | Forage Rape ‘Emerald’ | Forage Rape ‘Hobson’ | Turnip Rape ‘Jupiter’ | Tyfon | F5,15 | P | LSD | |
|---|---|---|---|---|---|---|---|---|---|
| Pollen beetle (Brassicogethes aeneus) (adults) | 2.790 (616.6) | 2.458 (287.1) | 2.622 (418.8) | 2.635 (431.5) | 2.628 (424.6) | 2.725 (530.9) | 1.74 | 0.186 | 0.257 |
| Pollen beetle (larvae) | 1.865 (73.3) | 1.578 (37.8) | 1.655 (45.2) | 1.890 (77.6) | 1.753 (56.6) | 2.216 (164.4) | 9.96 | <0.001 | 0.215 |
| Parasitoids of pollen beetle | 2.388 (244.3) | 1.716 (52.0) | 1.924 (83.9) | 1.909 (81.1) | 1.686 (48.5) | 1.647 (44.4) | 6.38 | 0.002 | 0.329 |
| Cabbage seed weevil (Ceutorhynchus obstrictus) adults | 0.527 (3.4) | 0.931 (8.5) | 0.389 (2.4) | 0.615 (4.1) | 1.033 (10.8) | 1.330 (21.4) | 14.04 | <0.001 | 0.286 |
| Parasitoids of cabbage seed weevil | 0.940 (8.7) | 1.353 (22.5) | 1.216 (16.4) | 1.299 (19.9) | 1.065 (11.6) | 1.315 (20.7) | 5.13 | 0.006 | 0.217 |
| Brassica pod midge (Dasineura brassicae) adults * | 0.815 (5.5) | 1.657 (44.4) | 1.906 (79.5) | 1.691 (48.1) | 1.418 (25.2) | 2.157 (142.5) | 11.08 | 0.006 | 0.418 |
| Parasitoids of brassica pod midge | 0.913 (8.2) | 1.500 (31.6) | 1.827 (67.1) | 1.647 (44.4) | 0.993 (9.8) | 1.457 (28.6) | 14.85 | <0.001 | 0.284 |
| Cabbage stem weevil (Ceutorhynchus pallidactylus) adults | 1.692 (49.2) | 0.714 (5.2) | 0.935 (8.6) | 1.089 (12.3) | 1.330 (21.4) | 1.221 (16.6) | 19.64 | <0.001 | 0.230 |
| Parasitoids of cabbage stem weevil * | 1.174 (13.9) | 0.556 (2.6) | 0.336 (1.2) | 0.345 (1.2) | 0.389 (1.4) | 0.301 (1.0) | 7.65 | <0.001 | 0.364 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skellern, M.P.; Clark, S.J.; Ferguson, A.W.; Watts, N.P.; Cook, S.M. Banker Plant Bonuses? The Benefits and Risks of Including Brassicas in Field Margins to Promote Conservation Biocontrol of Specialist Pests in Oilseed Rape. Insects 2023, 14, 349. https://doi.org/10.3390/insects14040349
Skellern MP, Clark SJ, Ferguson AW, Watts NP, Cook SM. Banker Plant Bonuses? The Benefits and Risks of Including Brassicas in Field Margins to Promote Conservation Biocontrol of Specialist Pests in Oilseed Rape. Insects. 2023; 14(4):349. https://doi.org/10.3390/insects14040349
Chicago/Turabian StyleSkellern, Matthew. P., Suzanne J. Clark, Andrew W. Ferguson, Nigel P. Watts, and Samantha M. Cook. 2023. "Banker Plant Bonuses? The Benefits and Risks of Including Brassicas in Field Margins to Promote Conservation Biocontrol of Specialist Pests in Oilseed Rape" Insects 14, no. 4: 349. https://doi.org/10.3390/insects14040349