Can Beauveria bassiana (Bals.-Criv.) Vuill. Control the Key Fruit Pests of the European Chestnut Tree, under Field Conditions?


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Treatments
2.2. Insects
2.3. Assessment of Mortality and Contamination of Larvae
2.4. Fungal Isolate
2.5. Weather Data
2.6. DNA Extraction, PCR Amplification and Sequencing
2.7. Data Analysis
- %Mcor = % of corrected mortality;
- %Mobs = % of mortality observed in treatment;
- %Mcont = % of mortality in the control group.
3. Results
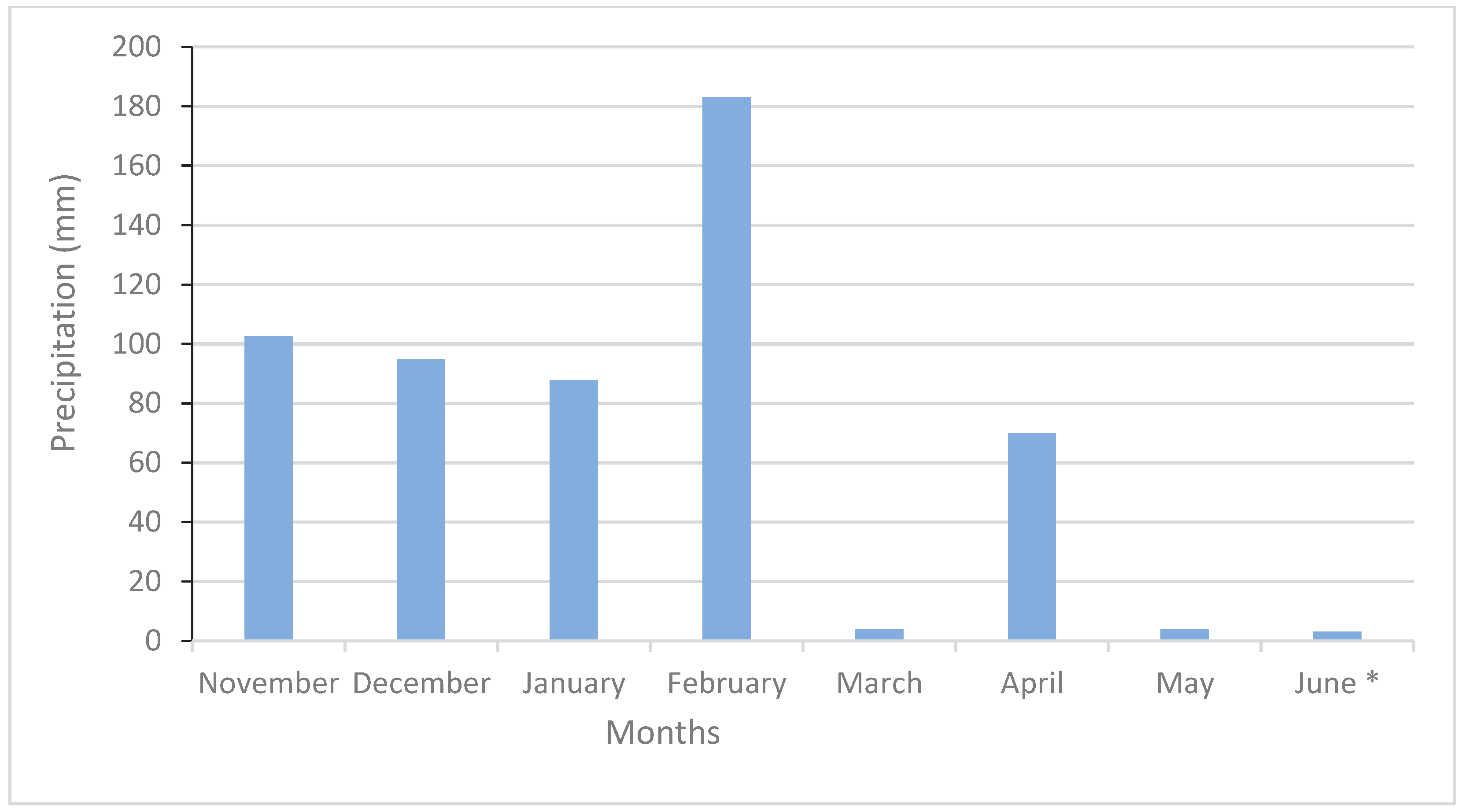
3.1. Weather Data
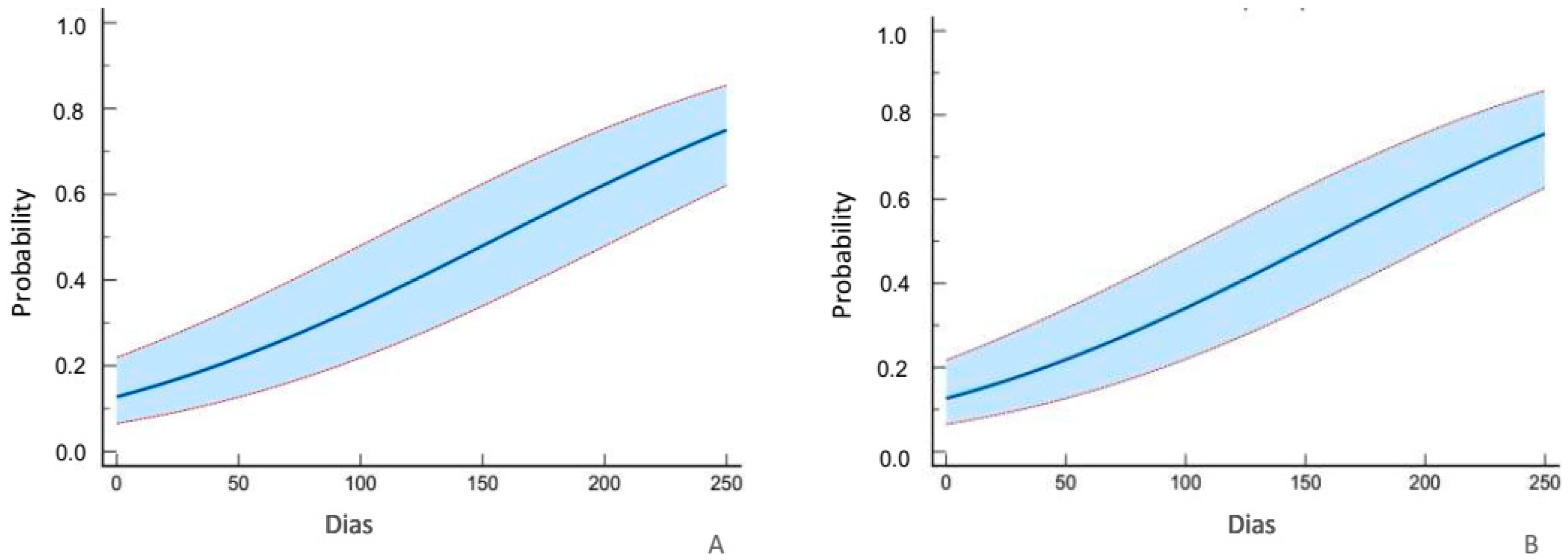
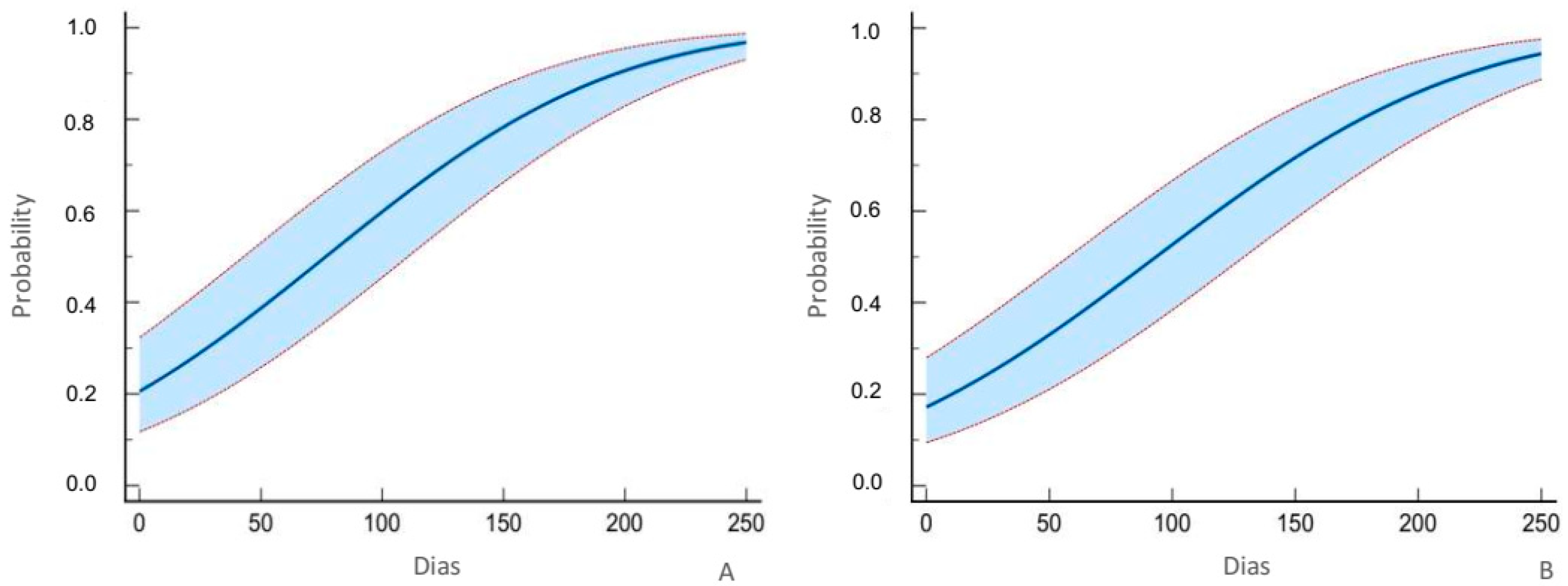
3.2. Efficacy of B. bassiana Strain ATCC74040 (Mortality)
3.3. Efficacy of B. bassiana Strain ATCC74040 (Total Mortality)
3.4. Mean Lethal Time LT50
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 12 January 2023).
- Instituto Nacional de Estatística, I.P (INE). Available online: https://www.ine.pt/xportal/xmain?xpid=INE&xpgid=ine_main (accessed on 15 January 2023).
- Conedera, M.; Tinner, W.; Krebs, P.; de Rigo, D.; Caudullo, G. Castanea sativa in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publ. Off. EU: Luxembourg, 2016; p. e0125e0+. [Google Scholar]
- Balsa, C.; Bento, A.; Paparella, F. Biological control of the Asian chestnut gall wasp in Portugal: Insights from a mathematical model. PLoS ONE 2021, 16, e0254193. [Google Scholar] [CrossRef]
- Bento, A.; Pereira, J.A.; Pereira, S. Pragas associadas à castanha em Trás-os-Montes: Biologia e estragos. In Proceedings of the II Congresso Ibérico do Castanheiro, Vila Real, Portugal, 20–22 June 2007. [Google Scholar]
- Ruocco, M.; Lanzuise, S.; Lombardi, N.; Varlese, R.; Aliberti, A.; Carpenito, S.; Woo, S.L.; Scala, F.; Lorito, M. New tools to improve the shelf life of chestnut fruit during storage. Acta Hortic. 2016, 1144, 309–315. [Google Scholar] [CrossRef]
- Marrão, R.; Lopes, E.; Bento, A. The chestnut moth, Cydia splendana (Hübner) (Lepidoptera: Torticidae) in Portugal. Millenium 2021, 2, 97–107. [Google Scholar] [CrossRef]
- Navarro, C.M.I. Diversidad Genética y Resistencia del Castaño a Factores Bióticos y Abióticos. Ph.D. Thesis, Universidad Córdoba, Córdoba, Spain, April 2019. Available online: http://hdl.handle.net/10396/18717 (accessed on 6 January 2023).
- Serrano, J.F.; Fernández, P.J.S.; Rodríguez, J.A.S.; Gutiérrez, F.J.D.P.; Martínez, P.M. El Castanõ. Manual y Guía Didáctica, 1st ed.; Instituto de Restauración y Medio Ambiente León, Espanha: León, Spain, 2001; pp. 323–327. [Google Scholar]
- Souza, M.E.C. Bioecologia de Pragas Carpófagas da Castanha e Controlo Microbiológico de Cydia splendana (Lepidoptera, Tortricidae) e Curculio elephas (Coleoptera, Curculionidae). Master’s Thesis, Instituto Politécnico de Bragança, Braganca, Portugal, 2022. Available online: http://hdl.handle.net/10198/25848 (accessed on 5 January 2023).
- Luís, M.; Martins, J.P.; Moura, J.V.; Cardoso, M.d.R.A.; Paula, O.; Paula, S. Arnaldo e Teresa Pinto AlertCast: Sistema de alertas para doenças e pragas do castanheiro. In Proceedings of the 10º Encontro Nacional de Proteção Integrada, Beja, Portugal, 2–3 May 2014; pp. 230–237. [Google Scholar]
- Butt, T.M.; Jackson, C.; Magan, N. Fungi as Biocontrol Agents: Progress Problems and Potential; CABI: Wallingford UK, 2001. [Google Scholar] [CrossRef]
- Jamal, Z. Application de Beauveria Bassiana Contre la Punaise terne Lygus lineolaris (Palisot de Beauvois) (Hémiptères: Miridés) dans les Vignobles; Université du Québec à Montréal: Montréal, QC, Canada, 2008. [Google Scholar]
- Lambert, N. Lutte Biologique aux Ravageurs: Applicabilité au Québec. Ph.D. Thesis, Université de Sherbrooke, Sherbrooke, QC, Canada, 2010. [Google Scholar]
- Inglis, G.D.; Goettel, M.S.; Butt, T.M.; Strasser, H. Use of hyphomycetous fungi for managing insect pests. In Fungi as Biocontrol Agents: Progress, Problems and Potential; CABI Publishing: Wallingford UK, 2001; pp. 23–69. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Sci. Technol. 2007, 17, 553–596. [Google Scholar] [CrossRef]
- Meyling, N.V.; Eilenberg, J. Ecology of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in temperate agroecosystems: Potential for conservation biological control. Biol. Control 2007, 43, 145–155. [Google Scholar] [CrossRef]
- Mwamburi, L.A.; Laing, M.D.; Miller, R.M. Effect of surfactants and temperature on germination and vegetative growth of Beauveria bassiana. Braz. J. Microbiol. 2015, 46, 67–74. [Google Scholar] [CrossRef]
- Kaiser, D.; Bacher, S.; Mène-Saffrané, L.; Grabenweger, G. Efficiency of natural substances to protect Beauveria bassiana conidia from UV radiation. Pest Manag. Sci. 2019, 75, 556–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacey, L.A. Entomopathogens used as microbial control agents. In Microbial Control of Insect and Mite Pests; Lacey, L.A., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 3–12. [Google Scholar] [CrossRef]
- Shelton, A.L.; Henning, J.A.; Schultz, P.; Clay, K. Effects of abundant white-tailed deer on vegetation, animals, mycorrhizal fungi, and soils. For. Ecol. Manag. 2014, 320, 39–49. [Google Scholar] [CrossRef]
- Goettel, M.S.; Eilenberg, J.; Glare, T. Entomopathogenic fungi and their role in regulation of insect populations. In Insect Control; Gilbert, L., Iatrou, K., Gill, S., Eds.; Elsevier: Boston, MA, USA, 2010; Volume 6361–6406. [Google Scholar]
- Wan, J.; Dai, W.; Ren, Z.; Chen, R.; Shi, Y. Study on isolation of pathogenic dematialaceyceous fungi from wild toads. Chin. J. Dermatol. 2003, 36, 461–463. [Google Scholar]
- Paparatti, B.; Speranza, S. Biological Control of Chestnut Weevil (Curculio Elephas Gyll.; Coleoptera, Curculionidae) with the Entomopathogen Fungus Beauveria bassiana (Balsamo) Vuill.(Deuteromycotina, Hyphomycetes). In Proceedings of the II International Symposium on Chestnut, Bordeaux, France, 9–23 October 1998. [Google Scholar] [CrossRef]
- Cerboneschi, A. Infection of Curculio elephas Gyll. and Cydia splendana Hb. larvae with the entomopathogenic fungus Beauveria bassiana (Balsamo). CABI 2012, 76, 257–261. [Google Scholar]
- Oliveira, I.V. Entomopathogenic Fungi Associated to Prays Oleae: Isolation, Characterization and Selection for Biological Control. Ph.D. Thesis, Universidade do Minho, Minho, Portugal, 2013. Available online: http://repositorium.sdum.uminho.pt/handle/1822/25728 (accessed on 3 February 2023).
- Aldebis, H.K.; Romero, Á.; Hatem, A.; Vargas-Osuna, E. Carpophagous insects of holm oak in southern Spain and their susceptibility to Beauveria bassiana native isolates. Integr. Prot. Oak For. IOBC-WPRS Bull. 2017, 127, 1–5. [Google Scholar]
- Cerboneschi, A.; Sechi, C. Indirect infection of Curculio elephas Gyll. larvae with the fungus Beauveria bassiana (Balsamo) Vuill. CABI 2017, 127, 68–71. [Google Scholar]
- Torrini, G.; Benvenuti, C.; Binazzi, F.; Marianelli, L.; Paoli, F.; Sabbatini Peverieri, G.; Roversi, P.F. Entomopathogenic fungi and nematodes against larvae of the chestnut weevil, Curculio elephas (Coleoptera: Curculionidae): A laboratory evaluation. Int. J. Pest Manag. 2018, 64, 287–293. [Google Scholar] [CrossRef]
- Sabbahi, R. Use of the Entomopathogenic Fungus Beauveria bassiana in a Strategy of Phytosanitary Management of the Main Insect Pests in Strawberry Plantations; University of Quebec: Quebec, QC, Canada, 2008. [Google Scholar]
- Zimmermann, G. The ‘Galleria bait method’for detection of entomopathogenic fungi in soil. J. Appl. Entomol. 1986, 102, 213–215. [Google Scholar] [CrossRef]
- Llácer, E.; Santiago-Álvarez, C.; Jacas, J.A. Could sterile males be used to vector a microbiological control agent? The case of Rhynchophorus ferrugineus and Beauveria bassiana. Bull. Entomol. Res. 2013, 103, 241–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In Pcr Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Püntener, W. Manual for Field Trials in Plant Protection; Ciba-Geigy Limited: Basle, Switzerland, 1981. [Google Scholar]
- Finney, D.J. Probit Analysis; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]
- Mkiga, A.M.; Mohamed, S.A.; du Plessis, H.; Khamis, F.M.; Akutse, K.S.; Ekesi, S. Metarhizium anisopliae and Beauveria bassiana: Pathogenicity, horizontal transmission, and their effects on reproductive potential of Thaumatotibia leucotreta (Lepidoptera: Tortricidae). J. Econ. Entomol. 2020, 113, 660–668. [Google Scholar] [CrossRef] [PubMed]
- Sarker, S.; Woo, Y.H.; Lim, U.T. Laboratory Evaluation of Beauveria bassiana ARP14 Against Grapholita molesta (Lepidoptera: Tortricidae). Curr. Microbiol. 2020, 77, 2365–2373. [Google Scholar] [CrossRef] [PubMed]
- Sahayaraj, K.; Namachivayam, S. Field evaluation of three entomopathogenic fungi on groundnut pests. Tropicultura 2011, 29, 143–147. [Google Scholar]
- Paparatti, B.; Speranza, S. Biological control of hazelnut weevil (Curculio nucum L., Coleoptera, Curculionidae) using the entomopathogenic fungus Beauveria bassiana (Balsamo) Vuill. (Deuteromycotina, Hyphomycetes). In Proceedings of the VI International Congress on Hazelnut, Tarragona-Reus, Spain, 30 July 2005. [Google Scholar] [CrossRef] [Green Version]
- Padmaja, V. Role of Entomopathogenic Fungi in Insect Pest Management. In Green Pesticides for Integrated Management; Ignachimuthu, S.J., Jayaraj, S., Eds.; Narosa Publishing House: New Delhi, India, 2005; p. 324. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cydia splendana (A) | p-value | ||||||
| D8 c | D15 bc | D62 bc | D100 ab | D220 a | <0.001 | ||
| T0 B | 0 | 0 A | 4.17 ± 0.54 A | 10.71 ± 0.47 A | 12 ± 0 B | 0.3639 | |
| T1 A | 0 c | 25 ± 1.25 Abc | 19.73 ± 0.47 Abc | 38 ± 1.41 Aab | 54.55 ± 0.47 Aa | 0.0178 | |
| T2 A | 0 c | 29.17 ± 0.7 Ab | 25.46 ± 1.19 Ab | 36.53 ± 1.44 Ab | 58.33 ± 1.44 Aa | 0.05 | |
| p-value | 0.012 | - | 0.6249 | 0.3984 | 0.1991 | 0.0216 | |
| Curculio elephas (B) | p-value | ||||||
| D8 b | D15 b | D62 b | D100 a | D220 a | <0.001 | ||
| T0 B | 0 A | 12.5 ± 0.27 A | 12.5 ± 0.47 B | 11.11 ± 0.47 B | 11.11 ± 0.47 B | 0.5251 | |
| T1 A | 6.67 ± 0.27 Ad | 7.64 ± 0.72 Ac | 22.29 ± 0.72 Ab | 83.93 ± 0 Aa | 85 ± 0.27 Aa | <0.001 | |
| T2 A | 3.33 ± 0.27 Ad | 26.11 ± 0.82 Ac | 33.33 ± 0.27 Ac | 61.21 ± 0.72 Ab | 88.75 ± 0 Aa | <0.001 | |
| p-value | <0.001 | 0.2963 | 0.182 | 0.0493 | <0.001 | <0.001 | |
| Cydia splendana (A) | p-value | ||||||
| D8 b | D15 b | D62 b | D100 b | D220 a | 0.0064 | ||
| T0 C | 4.17 ± 0.27 A | 6.67 ± 0.27 B | 8.33 ± 0.27 B | 10.71 ± 0.47 B | 10.71 ± 0 C | 0.5339 | |
| T1 B | 21.74 ± 0.47 Ab | 25.11 ± 0.27 Ab | 26.63 ± 0.47 Bb | 46.53 ± 0.72 Ab | 54.55 ± 0.47 Ba | 0.0017 | |
| T2 A | 21.74 ± 0.94 Ab | 37.5 ± 0.27 Ab | 45.45 ± 0.72 Ab | 40 ± 0.47 Ab | 88.64 ± 0.47 Aa | 0.0014 | |
| p-value | <0.001 | 0.2843 | 0.0091 | 0.0085 | 0.039 | <0.001 | |
| Curculio elephas (B) | p-value | ||||||
| D8 b | D15 b | D62 b | D100 a | D220 a | <0.001 | ||
| T0 B | 0 B | 3.33 ± 0.27 A | 8.33 ± 0.27 A | 7.4 ± 0 B | 11.11 ± 0.47 B | 0.5251 | |
| T1 A | 10 ± 0.82 Ab | 17.95 ± 0.82 Ab | 25.82 ± 0.54 Ab | 88.42 ± 0.27 Aa | 88.75 ± 0 Aa | <0.001 | |
| T2 A | 3.33 ± 0.27 Bc | 28.66 ± 0.47 Ab | 36.36 ± 0.98 Ab | 73.93 ± 0.27 Aa | 88.75 ± 0.72 Aa | <0.001 | |
| p-value | <0.001 | 0.0247 | 0.087 | 0.1322 | <0.001 | <0.001 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza, M.E.C.e.; Nóbrega, F.; Bento, A.A. Can Beauveria bassiana (Bals.-Criv.) Vuill. Control the Key Fruit Pests of the European Chestnut Tree, under Field Conditions? Insects 2023, 14, 342. https://doi.org/10.3390/insects14040342
Souza MECe, Nóbrega F, Bento AA. Can Beauveria bassiana (Bals.-Criv.) Vuill. Control the Key Fruit Pests of the European Chestnut Tree, under Field Conditions? Insects. 2023; 14(4):342. https://doi.org/10.3390/insects14040342
Chicago/Turabian StyleSouza, Maria Eliza Cota e, Filomena Nóbrega, and Albino António Bento. 2023. "Can Beauveria bassiana (Bals.-Criv.) Vuill. Control the Key Fruit Pests of the European Chestnut Tree, under Field Conditions?" Insects 14, no. 4: 342. https://doi.org/10.3390/insects14040342