
Infection Prevalence of Microsporidia Vairimorpha (Nosema) spp. in Japanese Bumblebees

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling of Bumblebees
2.2. Detection of Vairimorpha spp. Spores
2.3. PCR and Genotyping of Vairimorpha spp. Infections
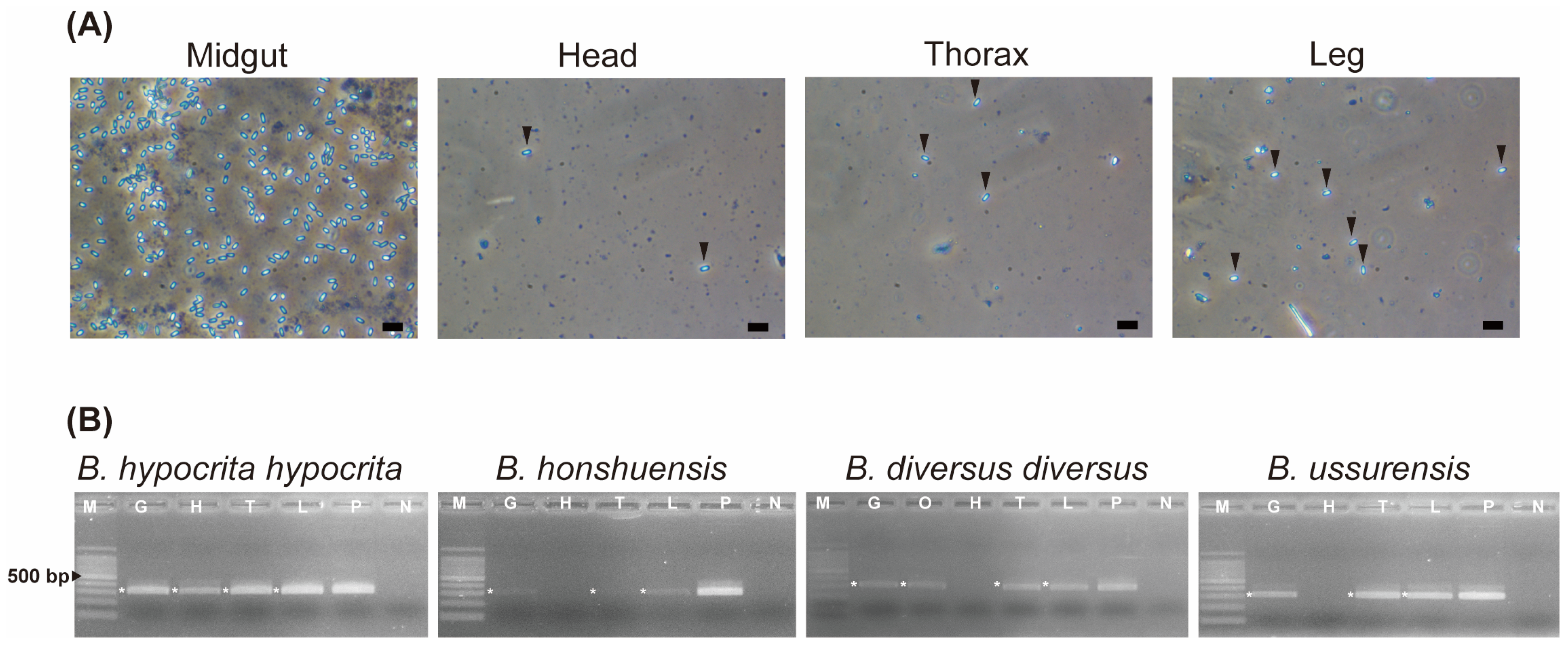
2.4. Organs Infected with Vairimorpha spp.
2.5. Quantitative PCR
3. Results
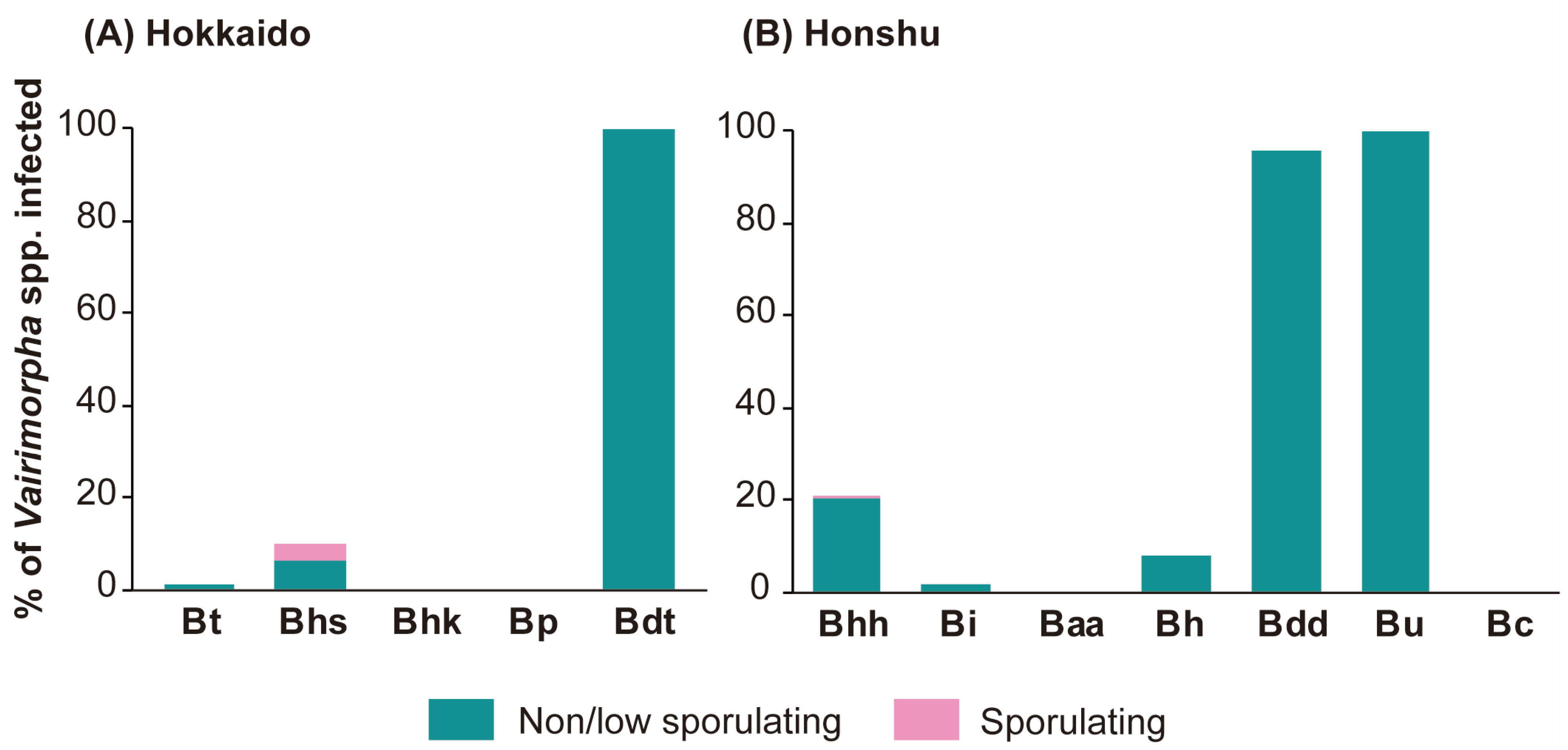
3.1. Prevalence of Vairimorpha spp.
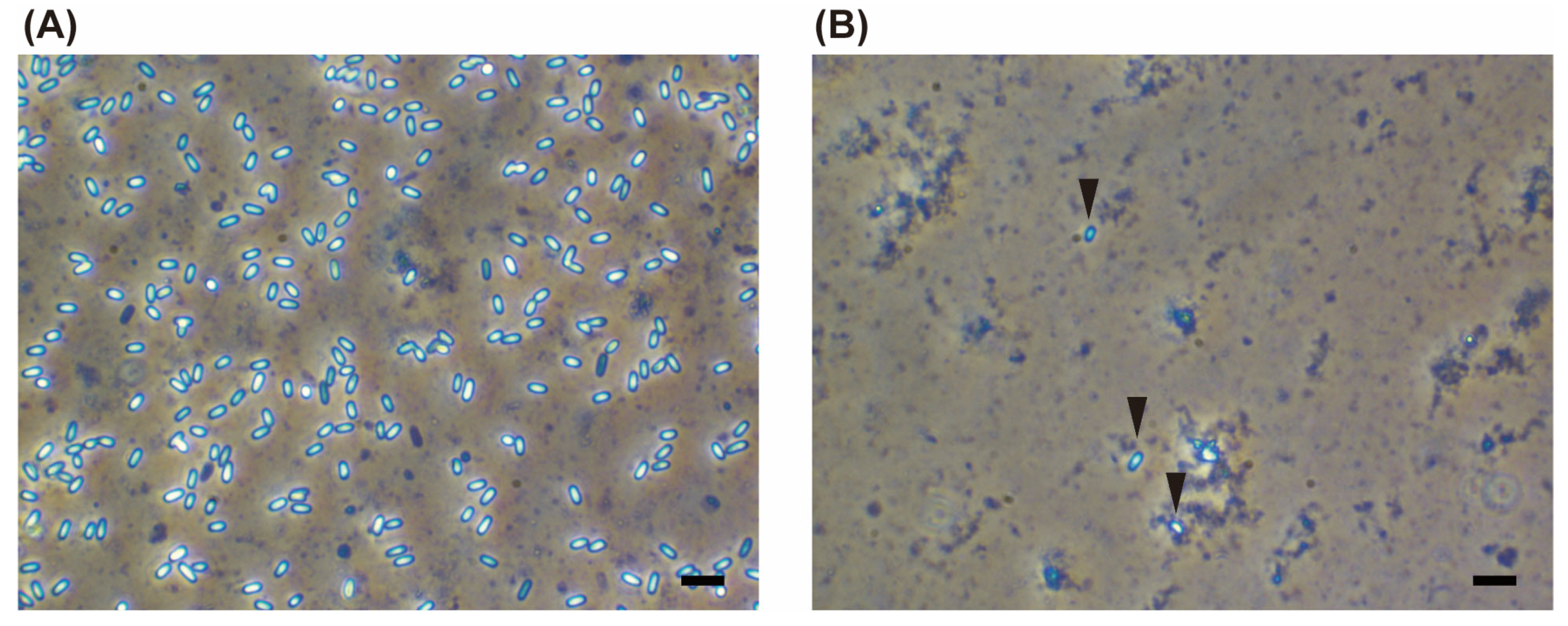
3.2. Organ Specificity of Vairimorpha spp.
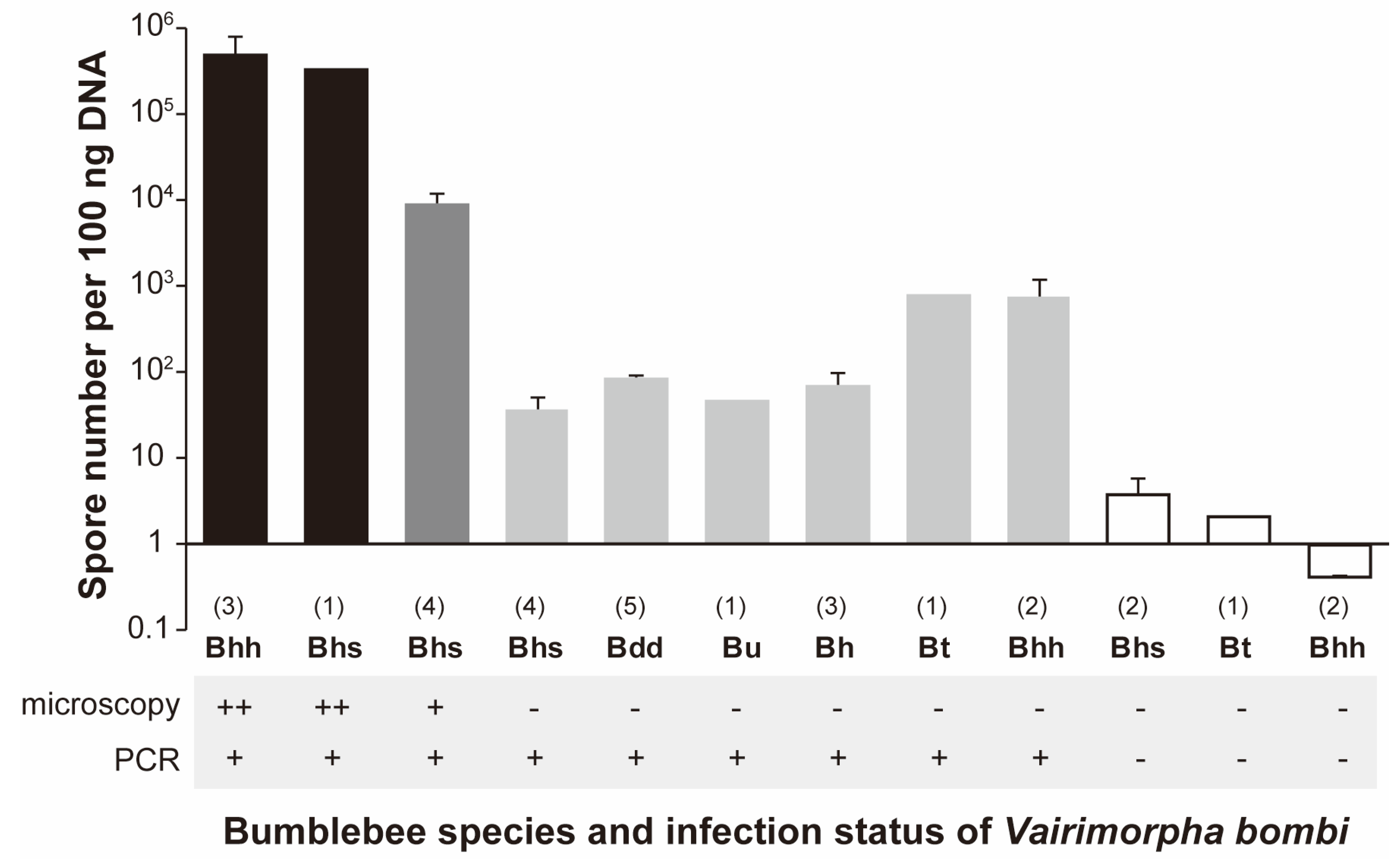
3.3. Spore Numbers of Vairimorpha spp. among Bumblebees
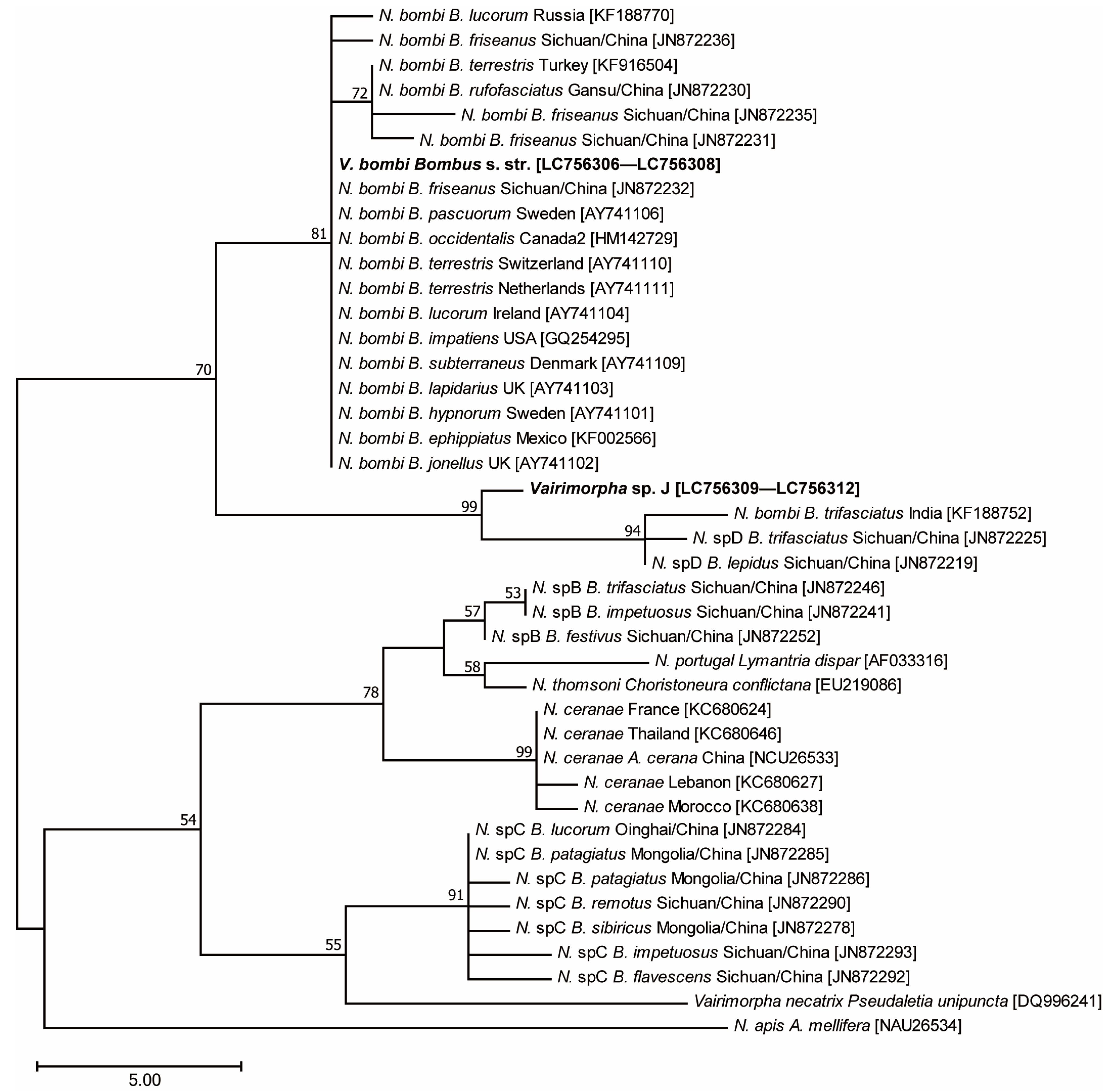
3.4. Phylogenetic Analysis Using Microsporidian SSU rRNA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kosior, A.; Celary, W.; Olejniczak, P.; Fijal, J.; Krol, W.; Solarz, W.; Plonka, P. The decline of the bumble bees and cuckoo bees (Hymenoptera: Apidae: Bombini) of western and central Europe. Oryx 2007, 41, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Cameron, S.A.; Lozier, J.D.; Strange, J.P.; Koch, J.B.; Cordes, N.; Solter, L.F.; Griswold, T.L. Patterns of widespread decline in North American bumble bees. Proc. Natl. Acad. Sci. USA 2011, 108, 662–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, S.A.; Lim, H.C.; Lozier, J.D.; Duennes, M.A.; Thorp, R. Test of the invasive pathogen hypothesis of bumble bee decline in North America. Proc. Natl. Acad. Sci. USA 2016, 113, 4386–4391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, L.M.; Becnel, J.J. Microsporidia: Pathogens of Opportunity; Wiley Blackwell: Ames, IA, USA, 2014. [Google Scholar]
- Fujiwara, T. Infectivity and pathogenicity of Nosema bombysis to larvae of the silkworm. J. Sericult. Sci. Jpn. 1979, 48, 376–380. [Google Scholar]
- Vilcinskas, A.; Stoecker, K.; Schmidtberg, H.; Roehrich, C.R.; Vogel, H. Invasive harlequin ladybird carries biological weapons against native competitors. Science 2013, 340, 862–863. [Google Scholar] [CrossRef]
- Tokarev, Y.S.; Huang, W.; Solter, L.F.; Malysh, J.M.; Becnel, J.J.; Vossbrinck, C.R. A formal redefinition of the genera Nosema and Vairimorpha (Microsporidia: Nosematidae) and reassignment of species based on molecular phylogenetics. J. Invertebr. Pathol. 2020, 169, 107279. [Google Scholar] [CrossRef]
- Fantham, H.B.; Porter, A. The morphology, biology and economic importance of Nosema bombi, n. sp., parasitic in various humble bees (Bombus Spp.). Ann. Trop. Med. Parasitol. 1914, 8, 623–638. [Google Scholar] [CrossRef]
- Otti, O.; Schmid-Hempel, P. Nosema bombi: A pollinator parasite with detrimental fitness effects. J. Invertebr. Pathol. 2007, 96, 118–124. [Google Scholar] [CrossRef]
- Rutrecht, S.T.; Brown, M.J.F. Differential virulence in a multiple-host parasite of bumble bees: Resolving the paradox of parasite survival? Oikos 2009, 118, 941–949. [Google Scholar] [CrossRef]
- Blaker, E.A.; Strange, J.P.; James, R.R.; Monroy, F.P.; Cobb, N.S. PCR reveals high prevalence of non/low sporulating Nosema bombi (Microsporidia) infections in bumble bees (Bombus) in Northern Arizona. J. Invertebr. Pathol. 2014, 123, 25–33. [Google Scholar] [CrossRef]
- Kissinger, C.N.; Cameron, S.A.; Thorp, R.W.; White, B.; Solter, L.F. Survey of bumble bee (Bombus) pathogens and parasites in Illinois and selected areas of northern California and southern Oregon. J. Invertebr. Pathol. 2011, 107, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Shykoff, J.A.; Schmidhempel, P. Incidence and effects of four parasites in populations of bumble bees in Switzerland. Apidologie 1991, 22, 117–125. [Google Scholar] [CrossRef]
- Huth-Schwarz, A.; Settele, J.; Moritz, R.F.A.; Kraus, F.B. Factors Influencing Nosema bombi infections in natural populations of Bombus terrestris (Hymenoptera: Apidae). J. Invertebr. Pathol. 2012, 110, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Cordes, N.; Huang, W.F.; Strange, J.P.; Cameron, S.A.; Griswold, T.L.; Lozier, J.D.; Solter, L.F. Interspecific geographic distribution and variation of the pathogens Nosema bombi and Crithidia species in United States bumble bee populations. J. Invertebr. Pathol. 2012, 109, 209–216. [Google Scholar] [CrossRef]
- Li, J.; Chen, W.; Wu, J.; Peng, W.; An, J.; Schmid-Hempel, P.; Schmid-Hempel, R. Diversity of Nosema associated with bumblebees (Bombus spp.) from China. Int. J. Parasitol. 2012, 42, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Vavilova, V.; Sormacheva, I.; Woyciechowski, M.; Eremeeva, N.; Fet, V.; Strachecka, A.; Bayborodin, S.I.; Blinov, A. Distribution and diversity of Nosema bombi (Microsporidia: Nosematidae) in the natural populations of bumblebees (Bombus spp.) from west Siberia. Parasitol. Res. 2015, 114, 3373–3383. [Google Scholar] [CrossRef]
- Szentgyorgyi, H.; Blinow, A.; Eremeeva, N.; Luzyanin, S.; Grzes, I.M.; Woyciechowski, M. Bumblebees (Bombidae) along pollution gradient—Heavy metal accumulation, species diversity, and Nosema bombi infection level. Pol. J. Ecol. 2011, 59, 599–610. [Google Scholar]
- Aytekin, A.M.; Cagatay, N.; Hazir, S. Floral choices, parasites and micro-organisms in natural populations of bumblebees (Apidae: Hymenoptera) in Ankara Province. Turk Zool. Derg. 2002, 26, 149–155. [Google Scholar]
- Evison, S.E.F.; Roberts, K.E.; Laurenson, L.; Pietravalle, S.; Hui, J.; Biesmeijer, J.C.; Smith, J.E.; Budge, G.; Hughes, W.O.H. Pervasiveness of parasites in pollinators. PLoS ONE 2012, 7, e30641. [Google Scholar] [CrossRef] [Green Version]
- Jabal-Uriel, C.; Martin-Hernandez, R.; Ornosa, C.; Higes, M.; Berriatua, E.; De la Rua, P. First data on the prevalence and distribution of pathogens in bumblebees (Bombus terrestris and Bombus pascuorum) from Spain. Span. J. Agric. Res. 2017, 15. [Google Scholar] [CrossRef] [Green Version]
- Vavilova, V.Y.; Konopatskaia, I.; Luzyanin, S.L.; Woyciechowski, M.; Blinov, A.G. Parasites of the genus Nosema, Crithidia and Lotmaria in the honeybee and bumblebee populations: A case study in India. Vavilovskij Žurnal Genet. I Sel. 2017, 21, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Sinpoo, C.; Disayathanoowat, T.; Williams, P.H.; Chantawannakul, P. Prevalence of Infection by the microsporidian Nosema spp. in native bumblebees (Bombus spp.) in northern Thailand. PLoS ONE 2019, 14, e0213171. [Google Scholar] [CrossRef] [Green Version]
- Votavová, A.; Trněný, O.; Staveníková, J.; Dybová, M.; Brus, J.; Komzáková, O. Prevalence and distribution of three bumblebee pathogens from the Czech Republic. Insects 2022, 13, 1121. [Google Scholar] [CrossRef]
- Rutrecht, S.T.; Klee, J.; Brown, M.J.F. Horizontal transmission success of Nosema bombi to its adult bumble bee hosts: Effects of dosage, spore source and host age. Parasitology 2007, 134, 1719–1726. [Google Scholar] [CrossRef] [PubMed]
- Donovan, B.J.; Wier, S.S. Development of hives for field population increase, and studies on the life cycles of the four species of introduced bumble bees in New Zealand. N. Z. J. Agric. Res. 1978, 21, 733–756. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D.; Hanley, M.E. Distribution and forage use of exotic bumblebees in South Island, New Zealand. N. Z. J. Ecol. 2004, 28, 225–232. [Google Scholar]
- MacFarlane, R.P.; Gurr, L. Distribution of bumble bees in New Zealand. N. Z. Entomol. 1995, 18, 29–36. [Google Scholar] [CrossRef]
- Dafni, A. The Threat of Bombus terrestris spread. Bee World 1998, 79, 113–114. [Google Scholar] [CrossRef]
- Montalva, J.; Sepulveda, V.; Vivallo, F.; Silva, D.P. New records of an invasive bumble bee in northern Chile: Expansion of its range or new introduction events? J. Insect Conserv. 2017, 21, 657–666. [Google Scholar] [CrossRef]
- Ruz, L.; Herrera, R. Preliminary observations on foraging activities of Bombus dahlbomii and Bombus terrestris (Hym: Apidae) on native and non-native vegetation in Chile. Acta Hortic. 2001, 561, 165–169. [Google Scholar] [CrossRef]
- Morales, C.L.; Arbetman, M.P.; Cameron, S.A.; Aizen, M.A. Rapid ecological replacement of a native bumble bee by invasive species. Front. Ecol. Environ. 2013, 11, 529–534. [Google Scholar] [CrossRef]
- Torretta, J.P.; Medan, D.; Abrahamovich, A.H. First record of the invasive bumblebee Bombus terrestris (L.) (Hymenoptera, Apidae) in Argentina. Trans. Am. Entomol. Soc. 2006, 132, 285–289. [Google Scholar]
- Hingston, A.B.; McQuillan, P.B. Does the recently introduced bumblebee Bombus terrestris (Apidae) threaten Australian ecosystems? Aust. J. Ecol. 1998, 23, 539–549. [Google Scholar] [CrossRef]
- Semmens, T.D.; Turner, E.; Buttermore, R. Bombus terrestris (L.) (Hymenoptera: Apidae) now established in Tasmania. Aust. J. Entomol. 1993, 32, 346. [Google Scholar] [CrossRef]
- Inoue, M.N.; Yokoyama, J.; Washitani, I. Displacement of Japanese native bumblebees by the recently introduced Bombus terrestris (L.) (Hymenoptera: Apidae). J. Insect Conserv. 2008, 12, 135–146. [Google Scholar] [CrossRef]
- Matsumura, C.; Yokoyama, J.; Washitani, I. Invasion status and potential ecological impacts of an invasive alien bumblebee, Bombus terrestris L. (Hymenoptera: Apidae) naturalized in southern Hokkaido, Japan. Glob. Environ. Res. 2004, 8, 51–66. [Google Scholar]
- Inari, N.; Nagamitsu, T.; Kenta, T.; Goka, K.; Hiura, T. Spatial and temporal pattern of introduced Bombus terrestris abundance in Hokkaido, Japan, and its potential impact on native bumblebees. Popul. Ecol. 2005, 47, 77–82. [Google Scholar] [CrossRef]
- Nukatsuka, Y.; Yokoyama, J. Environmental factors and land uses related to the naturalization of Bombus terrestris in Hokkaido, northern Japan. Biol. Invasions 2010, 12, 795–804. [Google Scholar] [CrossRef]
- Ono, M. Why Are Now the Bumblebees? Nat. Insects 1998, 33, 2–3. (In Japanese) [Google Scholar]
- Inoue, M.N.; Yokoyama, J.; Tsuchida, K. Colony growth and reproductive ability of feral nests of the introduced bumblebee Bombus terrestris in northern Japan. Insectes Soc. 2010, 57, 29–38. [Google Scholar] [CrossRef]
- Inoue, M.N.; Makino, T.T.; Yokoyama, J.; Sakai, S. Is Bombus terrestris (Hymenoptera: Apidae) a stronger competitor against native Japanese species? A comparison of foraging efficiency. Appl. Entomol. Zool. 2010, 45, 71–75. [Google Scholar] [CrossRef] [Green Version]
- Niwa, S.; Iwano, H.; Asada, S.; Matsuura, M.; Goka, K. Microsporidian pathogen isolated from a Colony of the European Bumblebees, Bombus Terrestris, and Infectivity on Japanese Bumblebee. Jap. J. Appl. Entomol. Zool. 2004, 48, 60–64. (In Japanese) [Google Scholar] [CrossRef] [Green Version]
- Takahashi, J.; Takeuchi, M.; Matsumoto, K.; Nomura, T. Prevalence of microsporidia isolated from honeybee and bumblebees in Japan. Bull. Res. Inst. Adv. Technol. Kyoto Sangyo Univ. 2013, 12, 59–68. (In Japanese) [Google Scholar]
- Schmid-Hempel, E.; Eckhardt, M.; Goulson, D.; Heinzmann, D.; Lange, C.; Plischuk, S.; Escudero, L.R.; Salathe, R.; Scriven, J.J.; Schmid-Hempel, P. The invasion of southern South America by imported bumblebees and associated parasites. J. Anim. Ecol. 2014, 83, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Arismendi, N.; Riveros, G.; Zapata, N.; Smagghe, G.; González, C.; Vargas, M. Occurrence of bee viruses and pathogens associated with emerging infectious diseases in native and non-native bumble bees in southern Chile. Biol. Invasions 2021, 23, 1175–1189. [Google Scholar] [CrossRef]
- Klee, J.; Tay, W.T.; Paxton, R.J. Specific and sensitive detection of Nosema bombi (Microsporidia: Nosematidae) in bumble bees (Bombus spp.; Hymenoptera: Apidae) by PCR of partial rRNA gene sequences. J. Invertebr. Pathol. 2006, 91, 98–104. [Google Scholar] [CrossRef]
- Tsugeno, Y.; Koyama, H.; Takamatsu, T.; Nakai, M.; Kunimi, Y.; Inoue, M.N. Identification of an early male-killing agent in the oriental tea tortrix, Homona magnanima. J. Hered. 2017, 108, 553–560. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Plischuk, S.; Martin-Hernandez, R.; Prieto, L.; Lucia, M.; Botias, C.; Meana, A.; Abrahamovich, A.H.; Lange, C.; Higes, M. South American native bumblebees (Hymenoptera: Apidae) infected by Nosema ceranae (Microsporidia), an emerging pathogen of honeybees (Apis Mellifera). Environ. Microbiol. Rep. 2009, 1, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Goka, K.; Okabe, K.; Yoneda, M.; Niwa, S. Bumblebee commercialization will cause worldwide migration of parasitic mites. Mol. Ecol. 2001, 10, 2095–2099. [Google Scholar] [CrossRef] [PubMed]
- Colla, S.R.; Otterstatter, M.C.; Gegear, R.J.; Thomson, J.D. Plight of the bumble bee: Pathogen spillover from commercial to wild populations. Biol. Conserv. 2006, 129, 461–467. [Google Scholar] [CrossRef]
- Meeus, I.; Brown, M.J.F.; De Graaf, D.C.; Smagghe, G. Effects of invasive parasites on bumble bee declines. Conserv. Biol. 2011, 25, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Szabo, N.D.; Colla, S.R.; Wagner, D.L.; Gall, L.F.; Kerr, J.T. Do Pathogen spillover, pesticide use, or habitat loss explain recent North American bumblebee declines? Conserv. Lett. 2012, 5, 232–239. [Google Scholar] [CrossRef]
- Graystock, P.; Yates, K.; Evison, S.E.F.; Darvill, B.; Goulson, D.; Hughes, W.O.H. The trojan hives: Pollinator pathogens, imported and distributed in bumblebee colonies. J. Appl. Ecol. 2013, 50, 1207–1215. [Google Scholar] [CrossRef]
- Tay, W.T.; O’Mahony, E.M.; Paxton, R.J. Complete rRNA gene sequences reveal that the microsporidium Nosema bombi infects diverse bumblebee (Bombus spp.) hosts and contains multiple polymorphic sites. J. Eukaryot. Microbiol. 2005, 52, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.H. A preliminary cladistic investigation of relationships among the bumble bees (Hymenoptera, Apidae). Syst. Entomol. 1985, 10, 239–255. [Google Scholar] [CrossRef]
- Woodard, S.H.; Lozier, J.D.; Goulson, D.; Williams, P.H.; Strange, J.P.; Jha, S. Molecular tools and bumble bees: Revealing hidden details of ecology and evolution in a model system. Mol. Ecol. 2015, 24, 2916–2936. [Google Scholar] [CrossRef] [PubMed]
- Katayama, E.; Ochiai, H.; Takamizawa, K. Supplementary notes on the nests of some japanese bumblebees: II. Bombus ussurensis. Jap. J. Entomol. 1990, 58, 335–346. [Google Scholar]
- Sokolova, Y.Y.; Lange, C.E. An ultrastructural study of Nosema locustae canning (Microsporidia) from three species of Acrididae (Orthoptera). Acta Protozool. 2002, 41, 229–236. [Google Scholar]
- Van der Steen, J.J.M. Infection and transmission of Nosema bombi in Bombus terrestris colonies and its effect on hibernation, mating and colony founding. Apidologie 2008, 39, 273–282. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Species * | |||||
|---|---|---|---|---|---|---|
| Bt | Bha | Bhk | Bp | Bdt | Total Specimen Number | |
| 2014 | Q79 (1/0) W30 (0/0) M19 (0/0) | Q106 (11/9) W56 (7/0) M33 (2/1) | Q1 (0/0) | W11 (0/0) M1 (0/0) | W1 (1/0) | 337 (22/10) |
| 2016 | Q61 (1/0) W9 (0/0) | Q65 (8/0) W2 (0/0) | Q1 (0/0) W10 (0/0) | 148 (9/0) | ||
| 2018 | W42 (1/0) | W45 (3/2) | 87 (4/2) | |||
| Total number | 240 (3/0) | 307 (31/12) | 12 (0/0) | 12 (0/0) | 1 (1/0) | 572 (35/12) |
| Year | Species * | Total Number | ||||||
|---|---|---|---|---|---|---|---|---|
| Bhh | Bi | Baa | Bh | Bdd | Bu | Bc | ||
| Hinohara, Tokyo | ||||||||
| 2016 | Q1 (0/0) W1 (0/0) | Q1 (0/0) W23 (0/0) | Q7 (7/0) | 33 (7/0) | ||||
| Oshino, Yamanashi | ||||||||
| 2016 | Q10 (0/0) | Q10 (0/0) | Q1 (1/0) | 21 (1/0) | ||||
| east OMA | ||||||||
| 2016 | Q4 (4/4) W11 (4/1) M5 (4/2) | W27 (4/0) M13 (0/0) | W14 (14/0) | W2 (0/0) | 62 (30/7) | |||
| 2019 | Q18 W27 M4 (-/15) ** | 49 (-/15) | ||||||
| 2020 | Q2 (0/0) W18 (1/0) M3 (0/0) | 23 (0/1) | ||||||
| west OMA | ||||||||
| 2014 | W8 (1/0) M7 (0/0) | M1(0/0) | W17(17/0) M5(5/0) | 38 (23/0) | ||||
| 2016 | W5 (0/0) M6 (1/0) | W46 (0/0) | W5(0/0) | W68(65/0) | W6(6/0) | 136 (72/0) | ||
| Hara, Nagano | ||||||||
| 2016 | M2 (0/0) | W3 (0/0) | W3 (0/0) M1 (0/0) | W9 (7/0) | 18 (7/0) | |||
| Total number | 117 (14+/24) | 64 (1/0) | 34 (0/0) | 50 (4/0) | 121 (116/0) | 6 (6/0) | 2 (0/0) | 380 (140+/23) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yanagisawa, T.; Kato, Y.; Inoue, M.N. Infection Prevalence of Microsporidia Vairimorpha (Nosema) spp. in Japanese Bumblebees. Insects 2023, 14, 340. https://doi.org/10.3390/insects14040340
Yanagisawa T, Kato Y, Inoue MN. Infection Prevalence of Microsporidia Vairimorpha (Nosema) spp. in Japanese Bumblebees. Insects. 2023; 14(4):340. https://doi.org/10.3390/insects14040340
Chicago/Turabian StyleYanagisawa, Takahiro, Yuto Kato, and Maki N. Inoue. 2023. "Infection Prevalence of Microsporidia Vairimorpha (Nosema) spp. in Japanese Bumblebees" Insects 14, no. 4: 340. https://doi.org/10.3390/insects14040340