
Microplitis manilae Ashmead (Hymenoptera: Braconidae): Biology, Systematics, and Response to Climate Change through Ecological Niche Modelling


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling, Preparation, Terminology, and Photography
2.2. Rearing Thai Specimens of Caterpillars and Parasitoid Wasp
2.3. Species Identifications
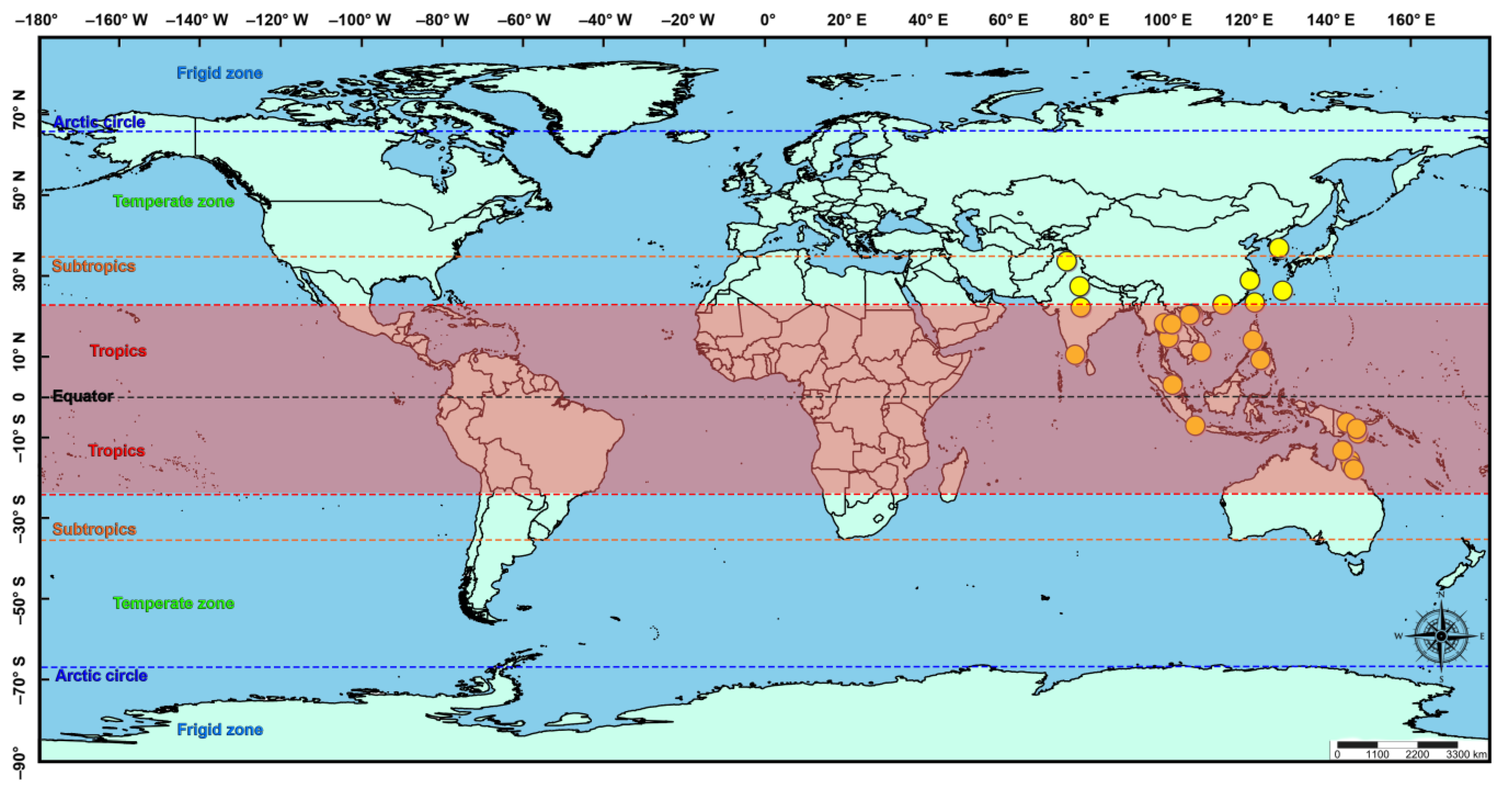
2.4. Ecological Niche Modelling
2.5. Depositories
3. Results
3.1. Taxonomic Account
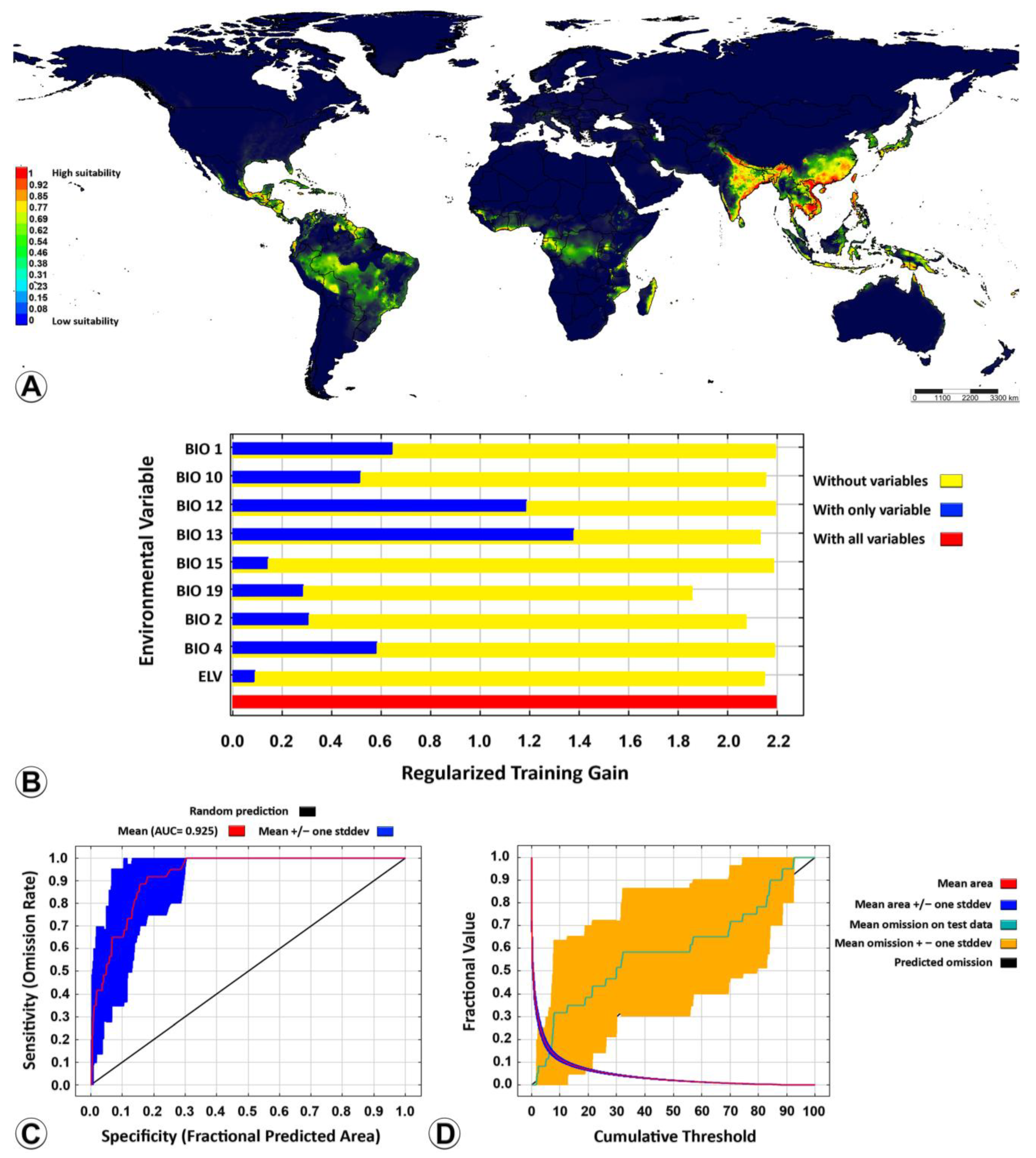
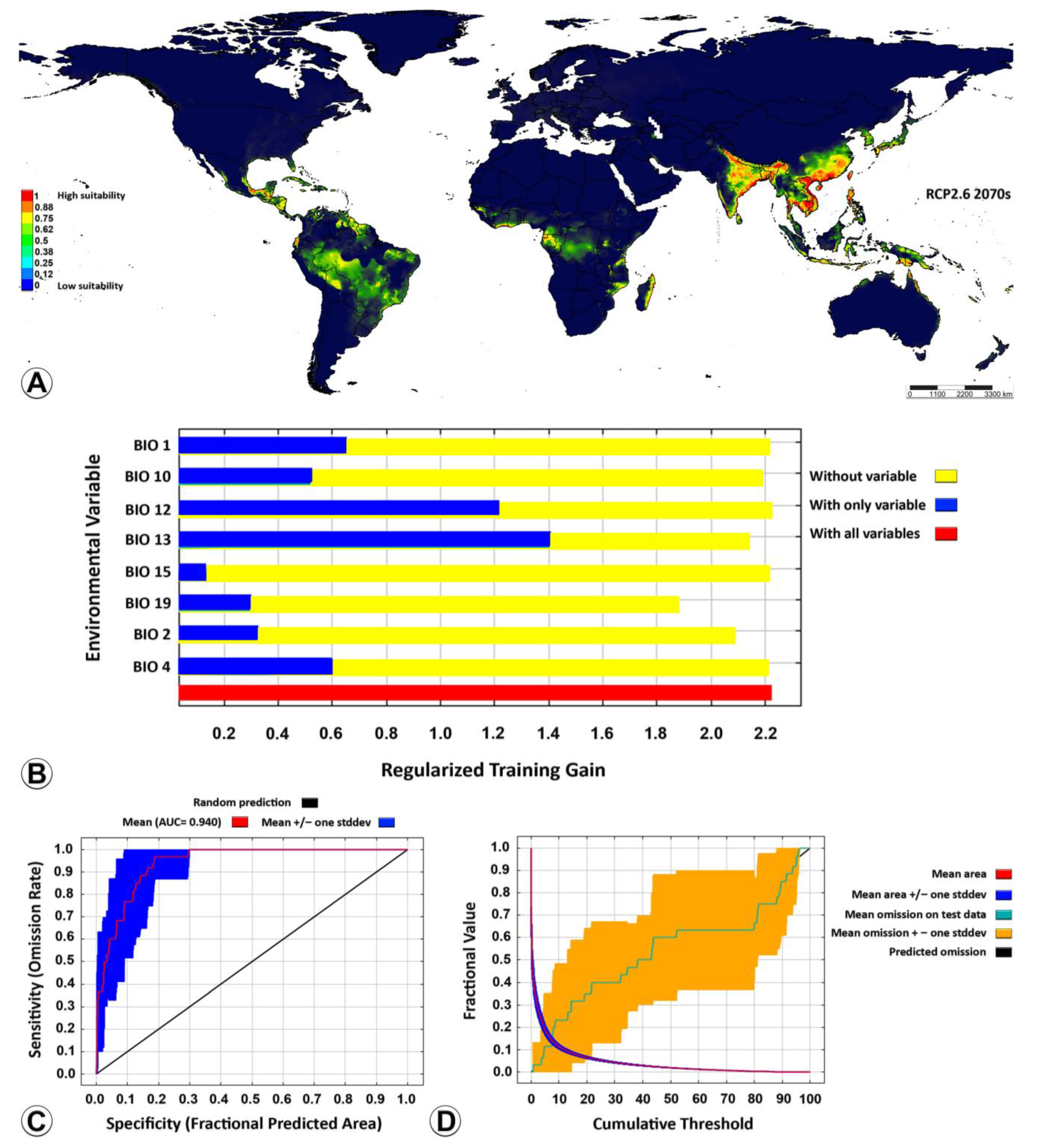
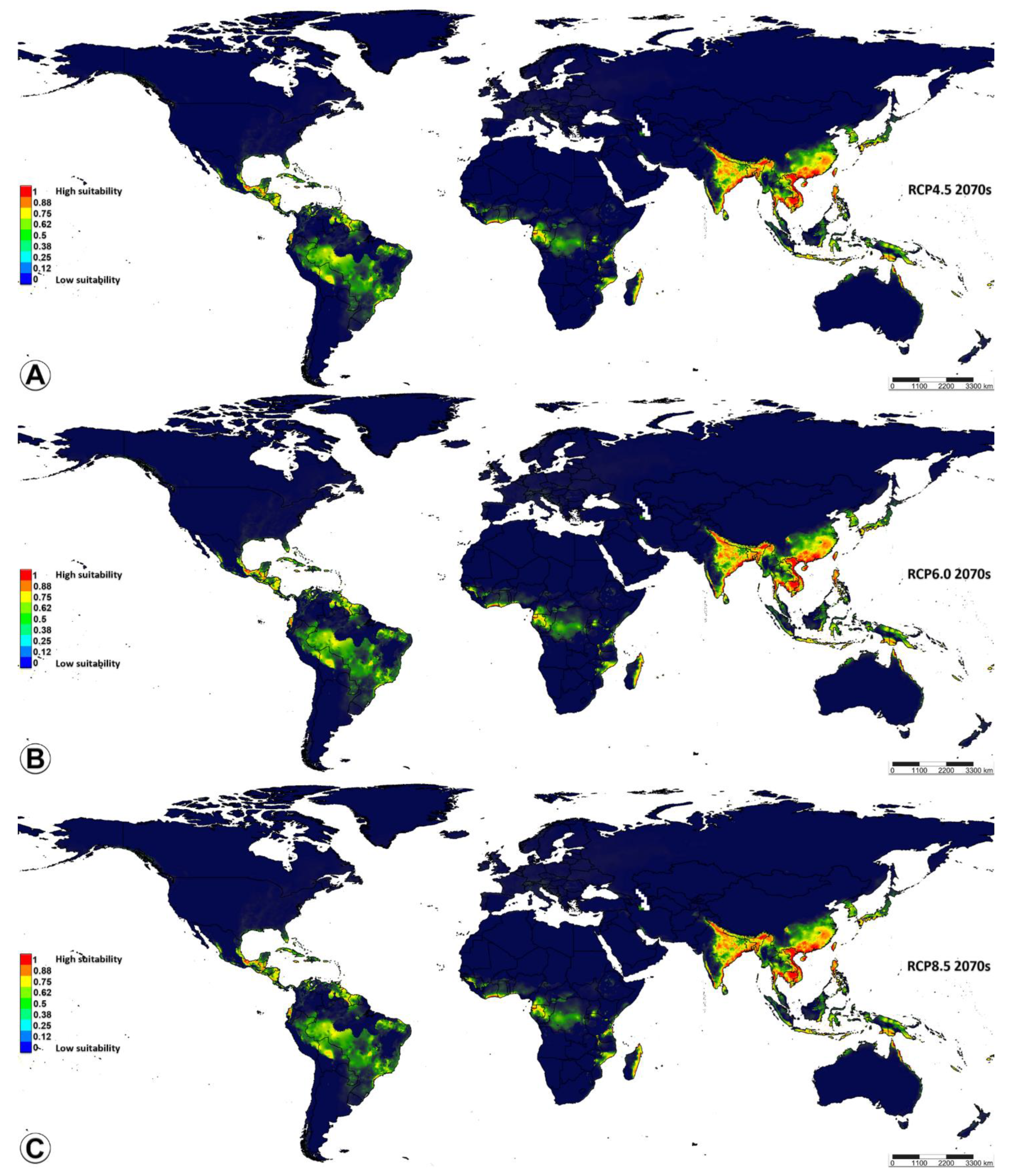
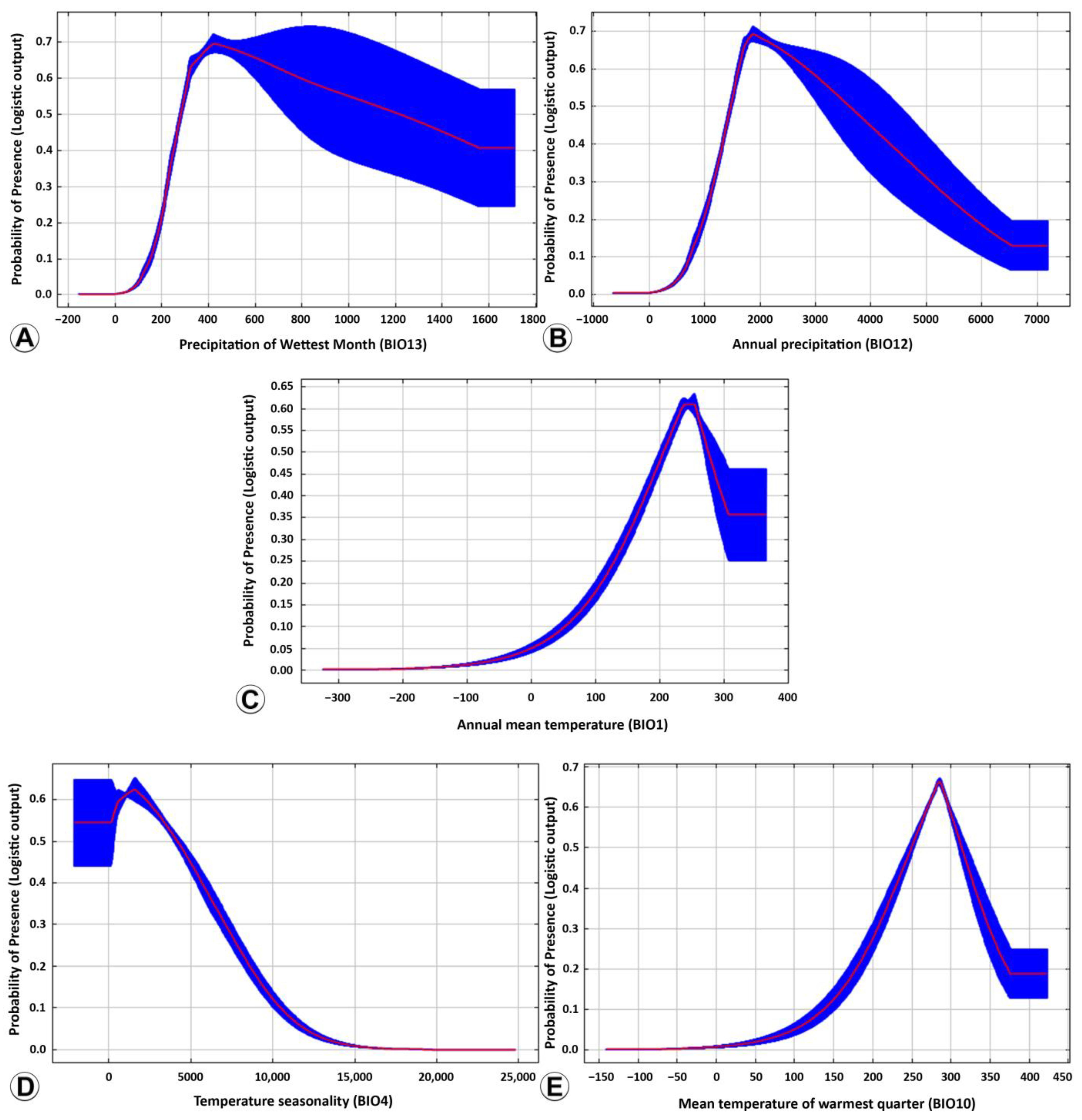
3.2. Ecological Niche Modelling
4. Discussion
4.1. Taxonomic Notes
4.2. Host-Parasitoid-Food Plant Associations
4.3. Ecological Niche Modelling
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Z.; Liu, Y.; Shi, M.; Huang, J.; Chen, X. Parasitoid wasps as effective biological control agents. J. Integr. Agric. 2019, 18, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Quicke, D.L.J. The Braconid and Ichneumonid Parasitic Wasps: Biology, Systematics, Evolution and Ecology; Wiley Blackwell: Chichester, UK, 2015; 688p. [Google Scholar] [CrossRef]
- Fernandez-Triana, J.L.; Shaw, M.R.; Boudreault, C.; Beaudin, M.; Broad, G.R. Annotated and illustrated world checklist of Microgastrinae parasitoid wasps (Hymenoptera, Braconidae). ZooKeys 2020, 920, 1–1089. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, J.B.; Austin, A.; Fernandez-Triana, J.L. Systematics, Biology, and Evolution of Microgastrine Parasitoid Wasps. Annu. Rev. Entomol. 2018, 63, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.; Mitter, M.; Regier, J.C. Systematics and evolution of the cutworm moths (Lepidoptera: Noctuidae): Evidence from two protein-coding nuclear genes. Syst. Entomol. 2006, 31, 21–46. [Google Scholar] [CrossRef]
- Pogue, M. A World Revision of the Genus Spodoptera Guenée (Lepidoptera: Noctuidae). Mem. Am. Entomol. Soc. 2002, 43, 1–202. [Google Scholar]
- Capinera, J.L. Fall Armyworm, Spodoptera frugiperda (J.E. Smith) (Insecta: Lepidoptera: Noctuidae); University of Florida: Gainesville, FL, USA, 2002; pp. 1–7. [Google Scholar] [CrossRef]
- Gupta, A. Revision of the Indian Microplitis Foerster (Hymenoptera: Braconidae: Microgastrinae), with description of one new species. Zootaxa 2013, 3620, 429–452. [Google Scholar] [CrossRef] [Green Version]
- Xing, B.; Yang, L.; Gulinuer, A.; Li, F.; Wu, S. Effect of Pupal Cold Storage on Reproductive Performance of Microplitis manilae (Hymenoptera: Braconidae), a Larval Parasitoid of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 449. [Google Scholar] [CrossRef]
- Gulinuer, A.; Xing, B.; Yang, L. Host Transcriptome Analysis of Spodoptera frugiperda Larvae Parasitized by Microplitis manilae. Insects 2023, 14, 100. [Google Scholar] [CrossRef]
- Frank, J.H.; McCoy, E.D. The introduction of insects into Florida. Fla. Entomol. 1993, 76, 1–53. [Google Scholar] [CrossRef]
- Qiu, B.; Zhou, Z.-S.; Luo, S.-P.; Xu, Z.-F. Effect of Temperature on Development, Survival, and Fecundity of Microplitis manilae (Hymenoptera: Braconidae). Environ. Entomol. 2012, 41, 657–664. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological Niches and Geographic Distributions (MPB-49); Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar] [CrossRef]
- Booth, T.H. Species distribution modelling tools and databases to assist managing forests under climate change. For. Ecol. Manag. 2018, 430, 196–203. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Pérez-De la O, N.B.; Espinosa-Zaragoza, S.; López-Martínez, V.D.; Hight, S.; Varone, L. Ecological Niche Modeling to Calculate Ideal Sites to Introduce a Natural Enemy: The Case of Apanteles opuntiarum (Hymenoptera: Braconidae) to Control Cactoblastis cactorum (Lepidoptera: Pyralidae) in North America. Insects 2020, 11, 454. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Triana, J.L.; Whitfield, J.; Rodriguez, J.; Smith, M.; Janzen, D.; Hajibabaei, M.; Burns, J.; Solis, A.; Brown, J.; Cardinal, S.; et al. Review of Apanteles sensu stricto (Hymenoptera, Braconidae, Microgastrinae) from Area de Conservación Guanacaste, northwestern Costa Rica, with keys to all described species from Mesoamerica. ZooKeys 2014, 383, 1–565. [Google Scholar] [CrossRef]
- Ghafouri Moghaddam, M.; Rakhshani, E.; van Achterberg, C.; Mokhtari, A. A study of the Iranian species of Choeras Mason (Hymenoptera: Braconidae: Microgastrinae), with the description of a new species. Zootaxa 2018, 4446, 455–476. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri Moghaddam, M.H.; Ghafouri Moghaddam, M.; Rakhshani, E.; Mokhtari, A. An upgrade pinning block: A mechanical practical aid for fast labelling of the insect specimens. Biodivers. Data J. 2017, 5, e20648. [Google Scholar] [CrossRef] [Green Version]
- Ghafouri Moghaddam, M.; Rakhshani, E.; van Achterberg, C.; Mokhtari, A. A taxonomic review of the genus Diolcogaster Ashmead (Hymenoptera, Braconidae, Microgastrinae) in Iran, distribution and morphological variability. Zootaxa 2019, 4590, 95–124. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.S.K.; van Achterberg, C.; Horstmann, K. Taxapad 2016, Ichneumonoidea 2015. Database on Flash-Drive. 2016, Nepean, Ontario. Available online: http://www.taxapad.com (accessed on 1 February 2023).
- Shorthouse, D.P. SimpleMappr, an Online Tool to Produce Publication-Quality Point Maps. 2010. Available online: https://www.simplemappr.net (accessed on 14 January 2023).
- Austin, A.D.; Dangerfield, P.C. Systematics of Australian and New Guinean Microplitis Foerster and Snellenius Westwood (Hymenoptera: Braconidae: Microgastrinae), with a review of their biology and host relationships. Invertebr. Taxon. 1993, 7, 1097–1166. [Google Scholar] [CrossRef]
- Ashmead, W.H. A list of Hymenoptera of the Philippine Islands with descriptions of new species. J. N. Y. Entomol. Soc. 1904, 12, 1–22. Available online: https://digitalcommons.usu.edu/bee_lab_an/95 (accessed on 5 February 2023).
- Guo, M.; Han, C.; Guan, Q.; Huang, Y. A universal parallel scheduling approach to polyline and polygon vector data buffer analysis on conventional GIS platforms. Trans. GIS 2020, 24, 1630–1654. [Google Scholar] [CrossRef]
- Petersen, M.J. Evidence of a climatic niche shift following North American introductions of two crane flies (Diptera; genus Tipula). Biol. Invasions 2013, 15, 885–897. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2007: Synthesis Report. Contribution of Working Groups I, II and III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2007; p. 104.
- Shcheglovitova, M.; Anderson, R.P. Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Modell. 2013, 269, 9–17. [Google Scholar] [CrossRef]
- Na, X.; Zhou, H.; Zang, S.; Wu, C.; Li, W.; Li, M. Maximum Entropy modeling for habitat suitability assessment of Red-crowned crane. Ecol. Indic. 2018, 91, 439–446. [Google Scholar] [CrossRef]
- Walden-Schreiner, C.; Leung, Y.F.; Kuhn, T.; Newburger, T.; Tsai, W.L. Environmental and managerial factors associated with pack stock distribution in high elevation meadows: Case study from Yosemite National Park. J. Environ. Manag. 2017, 193, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Barrion, A.T.; Bandong, J.P.; De La Cruz, C.G.; Apo Stol, R.F.; Litsinger, J.A. Natural enemies of the bean pod-borer, Maruca testulalis in the Philippines. Trop. Grain Leg. Bull. 1987, 34, 21–22. [Google Scholar]
- Ghafouri Moghaddam, M.; Tomlinson, S.; Jaffe, S.; Arias-Penna, D.C.; Whitfield, J.B.; Janzen, D.H.; Hallwachs, W.; Heidari Latibari, M. Review of the New World species of Microplitis Foerster (Hymenoptera, Braconidae, Microgastrinae) attacking Sphingidae (Lepidoptera, Bombycoidea). Insect Syst. Evol. 2022, 53, 221–241. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Ghafouri Moghaddam, M.; Arias-Penna, D.C.; Heidari Latibari, M. Notes on the genus Choeras Mason, 1981 (Hymenoptera: Ichneumonoidea, Braconidae, Microgastrinae) from Iran. Tijdschr. Voor Entomol. 2022, 165, 37–48. [Google Scholar] [CrossRef]
- Pante, E.; Schoelinck, C.; Puillandre, N. From integrative taxonomy to species description: One step beyond. Syst. Biol. 2015, 64, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Lyal, C.; Kirk, P.; Smith, D.; Smith, R. The value of taxonomy to biodiversity and agriculture. Biodiversity 2008, 9, 8–13. [Google Scholar] [CrossRef]
- Arias-Penna, D.C.; Whitfield, J.B.; Janzen, D.H.; Hallwachs, W.; Dyer, L.A.; Smith, M.A.; Hebert, P.D.N.; Fernández-Triana, J.L. A species-level taxonomic review and host associations of Glyptapanteles (Hymenoptera, Braconidae, Microgastrinae) with an emphasis on 136 new reared species from Costa Rica and Ecuador. ZooKeys 2019, 890, 1–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Bioclimatic Variables | Units | Being Used for Modelling |
|---|---|---|---|
| BIO1 | Annual mean temperature | °C | Yes |
| BIO2 | Mean diurnal range (mean of monthly temp (max temp–min temp)) | °C | Yes |
| BIO3 | Isothermality (BIO2/BIO7) × 100 | °C | No |
| BIO4 | Temperature seasonality | SD × 100 | Yes |
| BIO5 | Max temperature during the warmest month | °C | No |
| BIO6 | Min temperature during the coldest month | °C | No |
| BIO7 | Annual temperature range (BIO5–BIO6) | °C | No |
| BIO8 | Mean temperature during wettest quarter | °C | No |
| BIO9 | Mean temperature during driest quarter | °C | No |
| BIO10 | Mean temperature during warmest quarter | °C | Yes |
| BIO11 | Mean temperature during coldest quarter | °C | No |
| BIO12 | Annual precipitation | mm | Yes |
| BIO13 | Precipitation during the wettest month | mm | Yes |
| BIO14 | Precipitation during the driest month | mm | No |
| BIO15 | Precipitation seasonality | CV | Yes |
| BIO16 | Precipitation during the wettest quarter | mm | No |
| BIO17 | Precipitation during the driest quarter | mm | No |
| BIO18 | Precipitation during the warmest quarter | mm | No |
| BIO19 | Precipitation during the coldest quarter | mm | Yes |
| ELV | Elevation | m | Yes |
| Species | Author | Host | Host Plant | Cocoon | Larvae | MOR | MOL | BIO |
|---|---|---|---|---|---|---|---|---|
| Microplitis sp. | – | S. depravata (Butler) | ? | ? | ? | – | – | P |
| M. abrs | Austin and Dangerfield, 1993 | S. litura (Fabricius) | ? | ? | S | + | – | + |
| S. exigua (Hübner)? | ||||||||
| M. ajmerensis | Rao and Kurian, 1950 | S. exigua (Hübner) | ? | ? | S? | + | – | P |
| M. albotibialis | Telenga, 1955 | S. exigua (Hübner) | ? | ? | S? | + | – | P |
| M. bicoloratus | Chen, 2004 | S. litura (Fabricius) | Arachis hypogaea L. | ? | S | + | – | + |
| M. demolitor | Wilkinson, 1934 | S. frugiperda (J. E. Smith) | ? | ? | S | + | + | + |
| S. littoralis (Boisduval) | ? | |||||||
| S. litura (Fabricius) | ? | |||||||
| M. fulvicornis | (Wesmael, 1837) | S. exigua (Hübner) | Beta vulgaris L. | White grayish | S | + | – | + |
| M. leucaniae | Xu and He, 2002 | S. litura (Fabricius) | ? | ? | S? | + | – | P |
| M. manilae | Ashmead, 1904 | S. exempta (Walker) | ? | Light brown | S | + | P | + |
| S. exigua (Hübner) | ? | |||||||
| S. littoralis (Boisduval) | ? | |||||||
| S. litura (Fabricius) | Nicotiana tabacum L. Glycine max (L.) Brassica oleracea L. (var. italica, capitata, botrytis) | |||||||
| S. frugiperda (J. E. Smith) | Zea mays L. | |||||||
| M. pallidipes | Szépligeti, 1902 | S. litura (Fabricius) | ? | ? | S | + | – | + |
| S. exigua (Hübner) | ? | |||||||
| M. prodeniae | Rao and Kurian, 1950 | S. litura (Fabricius) | Amaranthus sp. Nicotiana tabacum L. | Light brown | S | + | + | + |
| M. rufiventris | Kokujev, 1914 | S. cilium Guenée | ? | Tan | S, G | + | + | + |
| S. exigua (Hübner) | Medicago sativa L. Zea mays L. | |||||||
| S. littoralis (Boisduval) | Gossypium hirsutum L. | |||||||
| S. frugiperda (J. E. Smith) | Glycine max (L.) | |||||||
| M. similis | Lyle, 1921 | S. litura (Fabricius) | Glycine max (L.) | ? | S, G | + | + | + |
| M. spectabilis | (Haliday, 1834) | S. exigua (Hübner) | ? | ? | S? | + | – | P |
| M. spodopterae | Rao and Kurian, 1950 | S. mauritia (Boisduval) | Trigonella foenum-graecum L. | Brown | S | + | – | + |
| M. tuberculifer | (Wesmael, 1837) | S. exigua (Hübner) | Zea mays L. | ? | S | + | P | + |
| S. litura (Fabricius) | ? |
| Code | Percent Contribution/% | Permutation Importance/% |
|---|---|---|
| At present time and under the climate change scenario RCP2.6 2070s | ||
| BIO13 | 67.4 | 43.5 |
| BIO19 | 18.6 | 21.5 |
| BIO2 | 5.3 | 5.6 |
| BIO10 | 2.5 | 20.9 |
| ALT | 2.4 | 5.1 |
| BIO4 | 1.9 | 0.4 |
| BIO1 | 1 | 1.5 |
| BIO15 | 0.5 | 1.2 |
| BIO12 | 0.4 | 0.3 |
| Under climate change scenarios RCP4.5, RCP6.0, and RCP8.5 in the period of the 2070s | ||
| BIO13 | 66.9 | 60.5 |
| BIO19 | 20.1 | 22.2 |
| BIO2 | 6.3 | 4.3 |
| BIO10 | 2.9 | 4.7 |
| BIO4 | 1.9 | 5.3 |
| BIO1 | 0.9 | 2.4 |
| BIO12 | 0.9 | 0.1 |
| BIO15 | 0.3 | 0.6 |
| Niche Overlap | Current | RCP8.5 2070s | RCP6.0 2070s | RCP4.5 2070s | RCP2.6 2070s |
|---|---|---|---|---|---|
| Current | 1 | ||||
| RCP8.5 2070s | 0.776919293 | 1 | |||
| RCP6.0 2070s | 0.812357129 | 0.881662813 | 1 | ||
| RCP4.5 2070s | 0.812198045 | 0.885771393 | 0.910782646 | 1 | |
| RCP2.6 2070s | 0.842805348 | 0.85744628 | 0.8886515 | 0.895545884 | 1 |
| Bioclimatic Variables | Suitable Range | Optimum Value |
|---|---|---|
| BIO13 (mm) | 269.2–763.4 | 435.8 |
| BIO19 (mm) | 175.1–704.8 | 342.3 |
| BIO2 (°C) | 64.3–107.4 | 80.6 |
| BIO10 (°C) | 198.4–326.2 | 292.4 |
| ALT (m) | −57.0–350.3 | −57.1 |
| BIO4 (SD × 100) | 132–4855.6 | 2240.7 |
| BIO1 (°C) | 197.3–286.4 | 246.7 |
| BIO15 (CV) | 59.4–160.1 | 83.5 |
| BIO12 (mm) | 1428.1–3982.5 | 1876.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghafouri Moghaddam, M.; Butcher, B.A. Microplitis manilae Ashmead (Hymenoptera: Braconidae): Biology, Systematics, and Response to Climate Change through Ecological Niche Modelling. Insects 2023, 14, 338. https://doi.org/10.3390/insects14040338
Ghafouri Moghaddam M, Butcher BA. Microplitis manilae Ashmead (Hymenoptera: Braconidae): Biology, Systematics, and Response to Climate Change through Ecological Niche Modelling. Insects. 2023; 14(4):338. https://doi.org/10.3390/insects14040338
Chicago/Turabian StyleGhafouri Moghaddam, Mostafa, and Buntika A. Butcher. 2023. "Microplitis manilae Ashmead (Hymenoptera: Braconidae): Biology, Systematics, and Response to Climate Change through Ecological Niche Modelling" Insects 14, no. 4: 338. https://doi.org/10.3390/insects14040338