
Additive Effect of Releasing Sterile Insects Plus Biocontrol Agents against Fruit Fly Pests (Diptera: Tephritidae) under Confined Conditions
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Areas
2.2. Biological Material
2.3. Founder Population
2.4. Adult Fly Emergence and Survival
2.5. Treatments
2.6. Sterile Insect Releases
2.7. Parameter Determination
2.8. Statistical Analysis
3. Results
3.1. Adult Fly Emergence
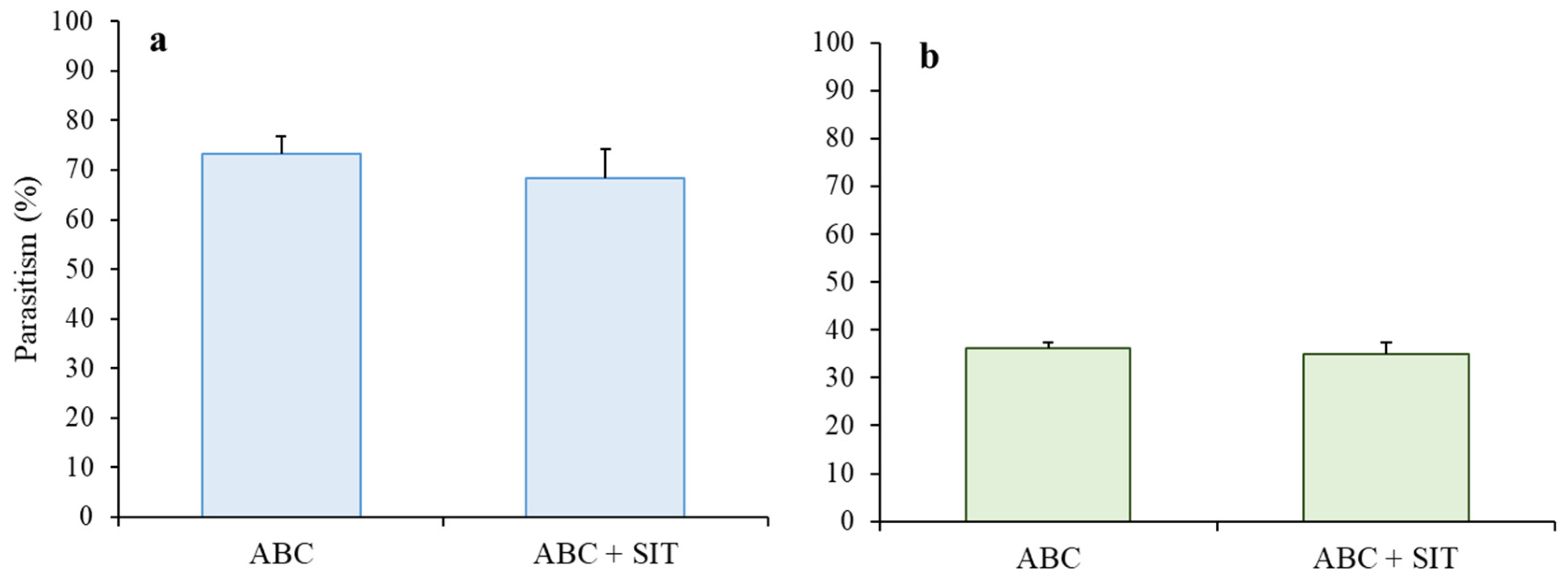
3.2. Parasitism
3.3. Induction of Sterility
3.4. Daily Net Reproduction
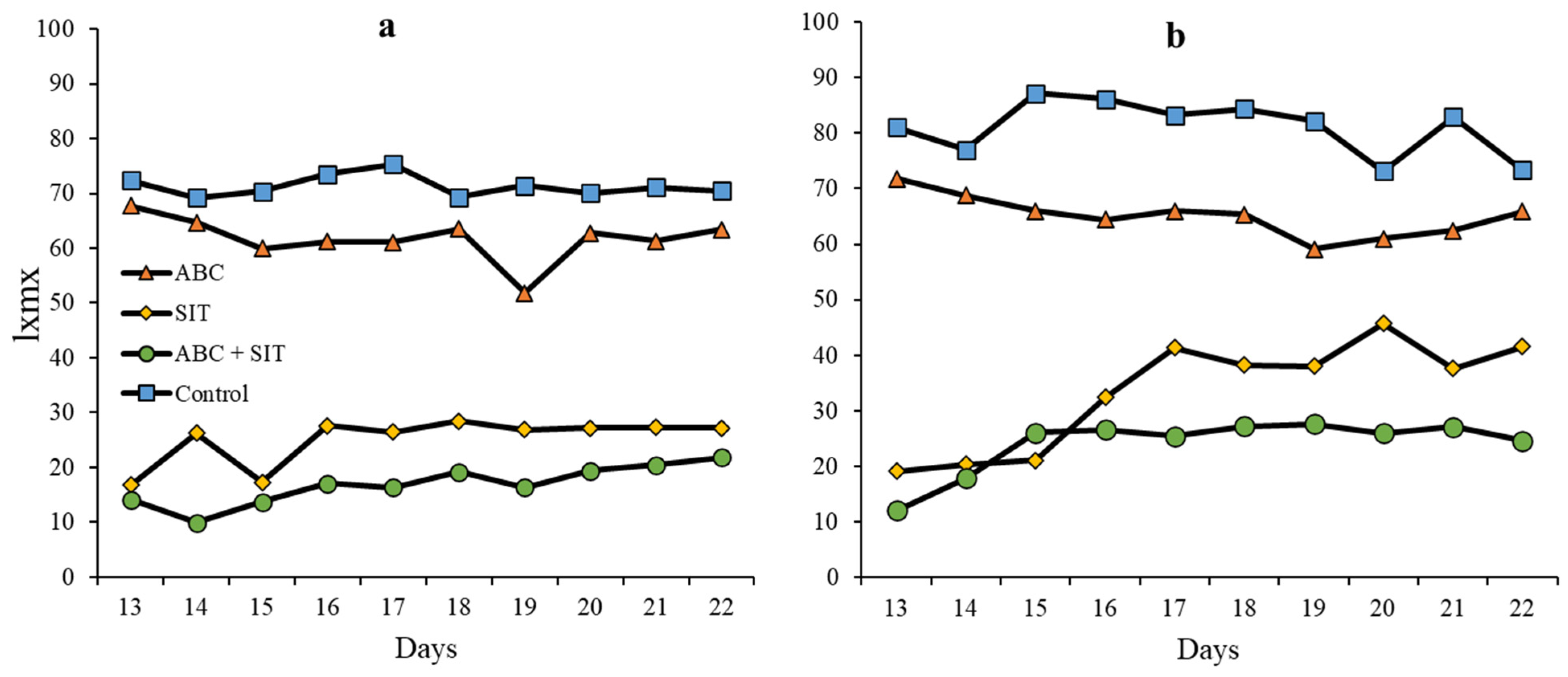
3.5. Survival
Reproductive Parameters
4. Discussions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barclay, H.J. Models for pest control: Complementary effects of periodic releases of sterile pest and parasitoids. Theor. Popul. Biol. 1987, 32, 76–89. [Google Scholar] [CrossRef]
- Knipling, E.F. Sterile insect and parasite augmentation techniques unexploited solutions for many insect pest problems. Fla. Entomol. 1998, 81, 34–160. [Google Scholar]
- Barclay, H.J. Mathematical models for using sterile insects. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dick, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRS Press: Boca Raton, FL, USA, 2021; pp. 201–242. [Google Scholar]
- Enkerlin, W. Impact of fruit fly control programmes using the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dick, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRS Press: Boca Raton, FL, USA, 2021; pp. 979–1005. [Google Scholar]
- Hendrichs, J.; Ortiz, G.; Liedo, P.; Schwarz, A. Six years of successful medfly program in Mexico and Guatemala. In Fruit Flies of Economic Importance; Cavalloro, R., Ed.; A. A. Balkema: Rotterdam, The Netherlands, 1983; pp. 353–365. [Google Scholar]
- Enkerlin, W.R.; Gutiérrez, J.M.; Pantaleon, R.; Soto, C.; Villaseñor, A.; Zavala, J.L.; Orozco, D.; Montoya, P.; Silva, L.; Cotoc, E.; et al. The Moscamed Regional Programme: A Success Story of Area-Wide Sterile Insect Technique Application. Entomol. Exp. Appl. 2017, 164, 188–203. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Staples, D.; Díaz-Fleischer, F.; Montoya, P. The Sterile Insect Technique: Success and Perspectives in the Neotropics. Neotrop. Entomol. 2020, 50, 172–185. [Google Scholar] [CrossRef]
- Dias, N.P.; Montoya, P.; Nava, D.E. Historical invasion of Medfly in the Neotropical region and adoption of management techniques. Curr. Opin. Insect. Sci. 2022, 50, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Liedo, P.; Montoya, P.; Toledo, J. Area-wide management of fruit flies in a tropical mango growing area integrating the sterile insect technique and biological control: From research to an operational programme. In Area-Wide Integrated Pest Management: Development and Field Application; Hendrichs, J., Pereira, R., Vreysen, M.J.B., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 197–214. [Google Scholar]
- Wong, T.T.Y.; Ramadan, M.M.; McInnis, D.O.; Mochizuki, N.; Nishimoto, J.A.; Herr, J.C. Augmentative releases of Diachasmimorpha tryoni (Hymenoptera: Braconidae) to suppress a Mediterranean fruit fly (Diptera: Tephritidae) population in Kula, Maui, Hawaii. Biol. Control 1991, 1, 2–7. [Google Scholar] [CrossRef]
- Sivinski, J.M.; Calkins, C.O.; Baranowski, R.M.; Harris, D.; Brambila, J.; Diaz, J.; Burns, R.E.; Holler, T.; Dodson, G. Suppression of a Caribbean fruit fly (Anastrepha suspensa (Loew) Diptera: Tephritidae) population through augmentative releases of the parasitoid Diachasmimorpha longicaudata (Ashmead) (Hymenoptera: Braconidae). Biol. Control 1996, 6, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Montoya, P.; Liedo, P.; Benrey, B.; Barrera, J.F.; Cancino, J.; Sivinski, J.; Aluja, M. Biological control of Anastrepha spp. (Diptera: Tephritidae) in mango orchards through augmentative releases of Diachasmimorpha longicaudata (Ashmead) (Hymenoptera: Braconidae). Biol. Control 2000, 18, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, G.; Murúa, F.; Suárez, L.; Van Nieuwenhove, G.; Taret, G.; Pantano, V.; Ovruski, S.M. Augmentative releases of Diachasmimorpha longicaudata (Hymenoptera: Braconidae) for Ceratitis capitata (Diptera: Tephritidae) control in a fruit-growing region of Argentina. Biol. Control 2016, 103, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Cancino, J.; Ruiz, L.; López, E.; Aguilar, E.; Gálvez, C.; Montoya, P.; Liedo, P. Suppression of Ceratitis capitata (Wied.) (Diptera: Tephritidae) populations in coffee in the Mexico-Guatemala border region through the augmentative releases of Diachasmimorpha longicaudata (Ashmead) (Hymenoptera: Braconidae). Biocontrol Sci. Technol. 2019, 29, 822–826. [Google Scholar] [CrossRef]
- Nagel, P.; Peveling, R. Environment and the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dick, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 754–780. [Google Scholar]
- Wong, T.T.; Ramadan, M.M.; Herr, J.C.; McInnis, D.O. Suppression of a Mediterranean fruit fly (Diptera: Tephritidae) population with concurrent parasitoid and sterile fly releases in Kula, Maui, Hawaii. J. Econ. Entomol. 1992, 85, 1671–1681. [Google Scholar] [CrossRef]
- Vargas, R.I.; Long, J.; Miller, N.W.; Delate, K.; Jackson, C.G.; Uchida, G.K.; Bautista, R.C.; Harris, E.J. Releases of Psyttalia fletcheri (Hymenoptera: Braconidae) and Sterile Flies to Suppress Melon Fly (Diptera: Tephritidae) in Hawaii. J. Econ. Entomol. 2004, 97, 1531–1539. [Google Scholar] [CrossRef] [PubMed]
- Rendón, P.; Sivinski, J.; Holler, T.; Bloem, K.; López, M.; Martínez, A.; Aluja, M. The effects of sterile males and two braconid parasitoids, Fopius arisanus (Sonan) and Diachasmimorpha krausii (Fullaway) (Hymenoptera), on caged populations of Mediterranean fruit flies, Ceratitis capitata (Wiedemann) (Diptera: Tephritidae) at various sites in Guatemala. Biol. Control 2006, 36, 224–231. [Google Scholar] [CrossRef]
- Aluja, M. Bionomics and management of Anastrepha. Annu. Rev. Entomol. 1994, 39, 155–178. [Google Scholar] [CrossRef]
- Montoya, P.; Cancino, J.; Zenil, M.; Santiago, G.; Gutierrez, J.M. The augmentative biological control component in the Mexican Campaign against Anastrepha spp. fruit flies. In Area-Wide Control of Insect Pests: From Research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 661–670. [Google Scholar]
- Orozco-Dávila, D.; Quintero, L.; Hernández, E.; Solís, E.; Artiaga, T.; Hernández, R.; Montoya, P. Mass rearing and sterile insect releases for the control of Anastrepha spp. (Diptera: Tephritidae) pests in Mexico—A review. Entomol. Exp. Appl. 2017, 164, 176–187. [Google Scholar] [CrossRef] [Green Version]
- Zepeda-Cisneros, C.S.; Meza-Hernández, J.S.; Palacios, J.I.; Martínez, V.G.; de León Crisóstomo, A.H.; Flores–García, H. Development and evaluation of genetic sexing strain of Anastrepha ludens for sterile insect technique. In Proceedings of the International Congress of Entomology, Daegu, Republic of Korea, 20–24 August 2012. [Google Scholar]
- Orozco, D.; Meza, J.S.; Zepeda, S.; Solís, E.; Quintero, J.L. Tapachula-7, a new genetic sexing strain of the Mexican fruit fly (Diptera: Tephritidae): Sexual compatibility and competitiveness. J. Econ. Entomol. 2013, 106, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Cancino, J.; Ruíz, L.; López, P.; Moreno, F.M. Cría masiva de parasitoides. In Moscas de la Fruta: Fundamentos y Procedimientos para Su Manejo; Montoya, P., Toledo, J., Hernández, E., Eds.; S y G Editores: Ciudad de México, Mexico, 2020; pp. 463–481. [Google Scholar]
- Cancino, J.; Montoya, P. Advances and Perspectives in the mass rearing of fruit fly parasitoids in Mexico. In Fruit Flies of Economic Importance: From Basic to Applied knowledge; Sugayama, R., Zucchi, R.A., Ovruski, S.M., Sivinski, J., Eds.; Press Color: Bahia, Brazil, 2008; pp. 133–142. [Google Scholar]
- Cancino, J.; Liedo, P.; Ruiz, L.; López, G.; Montoya, P.; Barrera, J.F.; Sivinski, J.; Aluja, M. Discrimination by Coptera haywardi (Hymenoptera: Diapriidae) of hosts previously attacked by conspecifics or by the larval parasitoid Diachasmimorpha longicaudata (Hymenoptera: Braconidae). Biocontrol Sci. Technol. 2012, 22, 899–914. [Google Scholar] [CrossRef]
- Sivinski, J.M.; Vulinec, K.; Menezes, E.; Aluja, M. The bionomics of Coptera haywardi (Oglobin) (Hymenoptera: Diapridae) and the other pupal parasitoids of tephritid fruit flies (Diptera). Biol. Control 1998, 11, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Guillén, L.; Aluja, M.; Equihua, M.; Sivinski, J. Performance of two fruit fly (Diptera: Tephritidae) pupal parasitoids Coptera haywardi (Hymenoptera: Diapriidae) and Pachycrepoideus vindemiae (Hymenoptera: Pteromalidae) under different environmental soil conditions. Biol. Control 2001, 23, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Baeza-Larios, G.; Sivinski, J.; Holler, T.; Aluja, M. The ability of Coptera haywardi (Oglobin) (Hymenoptera: Diapriidae) to locate and attack the pupae of the Mediterranean fruit fly, Ceratitis capitata (Wiedemann) (Diptera: Tephritidae), under seminatural conditions. Biol. Control 2002, 23, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Montoya, P.; Gálvez, C.; Díaz-Fleischer, F. Host availability affects the interaction between pupal parasitoid Coptera haywardi (Hymenoptera: Diapriidae) and larval–pupal parasitoid Diachasmimorpha longicaudata (Hymenoptera: Braconidae). Bull. Entomol. Res. 2019, 109, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Flores, S.; Montoya, P.; Toledo, J.; Enkerlin, W.; Liedo, P. Estimation of Populations and Sterility Induction in Anastrepha ludens (Diptera: Tephritidae) Fruit Flies. J. Econ. Entomol. 2014, 107, 1502–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, S.; Gómez-Escobar, E.; Liedo, P.; Toledo, J.; Montoya, P. Density estimation and optimal sterile: Wild ratio to induce sterility in Anastrepha obliqua (Diptera: Tephritidae) populations. Entomol. Exp. Appl. 2017, 164, 284–290. [Google Scholar] [CrossRef] [Green Version]
- Carey, J.R. Applied Demography for Biologists: With Special Emphasis on Insects; Oxford University Press: New York, NY, USA, 1993; 206p. [Google Scholar]
- Knipling, F.E. Principles of Insect Parasitism Analyzed from New Perspectives; Agriculture Handbook; USDA, ARS: Washington DC, USA, 1992; pp. 1–159.
- Sivinski, J.; Jeronimo, F.; Holler, T. Development of aerial releases of Diachasmimorpha tryoni (Cameron) (Hymenoptera: Braconidae), a parasitoid that attacks the Mediterranean fruit fly, Ceratitis capitata (Weidemann) (Diptera: Tephritidae), in the Guatemalan highlands. Biocontrol Sci. Techol. 2000, 10, 15–25. [Google Scholar] [CrossRef]
- Harbi, A.; Beitia, F.J.; Tur, C.; Chermiti, B.; Verdú, M.J.; Sabater-Muñoz, B. Field releases of the larval parasitoid Diachasmimorpha longicaudata in Spain: First results on dispersal pattern. Acta Hortic. 2015, 1065, 1057–1062. [Google Scholar] [CrossRef]
- Müller, F.A.; Dias, N.P.; Gottschalk, M.S.; Garcia, F.R.M.; Nava, D.E. Potential distribution of Bactrocera oleae and the parasitoids Fopius arisanus and Psyttalia concolor, aiming at classical biological control. Biol. Control 2019, 132, 144–151. [Google Scholar] [CrossRef]
- Hendrichs, J.; Enkerlin, W.; Pereira, R. Invasive Insect pests: Challenges and the role of the Sterile Insect Technique in their prevention, containment and eradication. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dick, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 885–921. [Google Scholar]
- Liebhold, A.M.; Work, T.T.; McCullough, D.G.; Carey, J.F. Airline baggage as a pathway for alien insect species invading the United States. Am. Entomol. 2006, 52, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Zavala-Lopez, J.L.; Marte-Diaz, G.; Martínez-Pujols, F. Successful area-wide eradication of the invading Mediterranean fruit fly in the Dominican Republic. In Area-Wide Integrated Pest Management; Hendrichs, J., Pereira, R., Vreysen, M.J.B., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 519–537. [Google Scholar]
- Argov, Y.; Gazit, Y. Biological control of the Mediterranean fruit fly in Israel: Introduction and establishment of natural enemies. Biol. Control 2008, 46, 502–507. [Google Scholar] [CrossRef]
- Vinson, S.B. The general host selection behavior of parasitoid Hymenoptera and a comparison of initial strategies utilized by larvaphagous and oophagous species. Biol. Control 1998, 11, 79–96. [Google Scholar] [CrossRef] [Green Version]
- Dias, N.P.; Nava, D.E.; Zotti, M.J.; Montoya, P.; Carvalho, I.R. Fruit fly management research: A systematic review of monitoring and control tactics. Crop. Prot. 2018, 112, 187–200. [Google Scholar] [CrossRef]
- Montoya, P.; Ayala, A.; López, P.; Cancino, J.; Cabrera, H.; Cruz, J.; Martínez, A.M.; Figueroa, I.; Liedo, P. Natural parasitism in fruit fly populations in disturbed areas adjacent to commercial mango orchards in Chiapas and Veracruz, Mexico. Environ. Entomol. 2016, 42, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Montoya, P.; López, P.; Cruz, J.; López, F.; Cadena, C.; Cancino, J.; Liedo, P. Effect of Diachasmimorpha longicaudata releases on the native parasitoid guild attacking Anastrepha spp. larvae in disturbed zones of Chiapas, Mexico. BioControl. 2017, 62, 581–593. [Google Scholar] [CrossRef]
- Sivinski, J.; Aluja, M.; López, M. Spatial and temporal distributions of parasitoids of Mexican Anastrepha species (Diptera: Tephritidae) within the canopies of fruit trees. Ann. Entomol. Soc. Am. 1997, 90, 604–618. [Google Scholar] [CrossRef]
- Shelly, T.E.; Whittier, T.S. Mating competitiveness of sterile male Mediterranean fruit flies (Diptera: Tephritidae) in male-only releases. Ann. Entomol. Soc. Am. 1996, 89, 754–758. [Google Scholar] [CrossRef]
- Shelly, T.E.; Edu, J.; Pahio, E. Age-dependent variation in mating success of sterile male Mediterranean fruit flies (Diptera: Tephritidae): Implications for sterile insect technique. J. Econ. Entomol. 2007, 100, 1180–1187. [Google Scholar] [CrossRef] [PubMed]
- Klassen, W.; Vreysen, M.J.B. Area-wide integrated pest management and the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dick, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 75–111. [Google Scholar]




{kind=link}
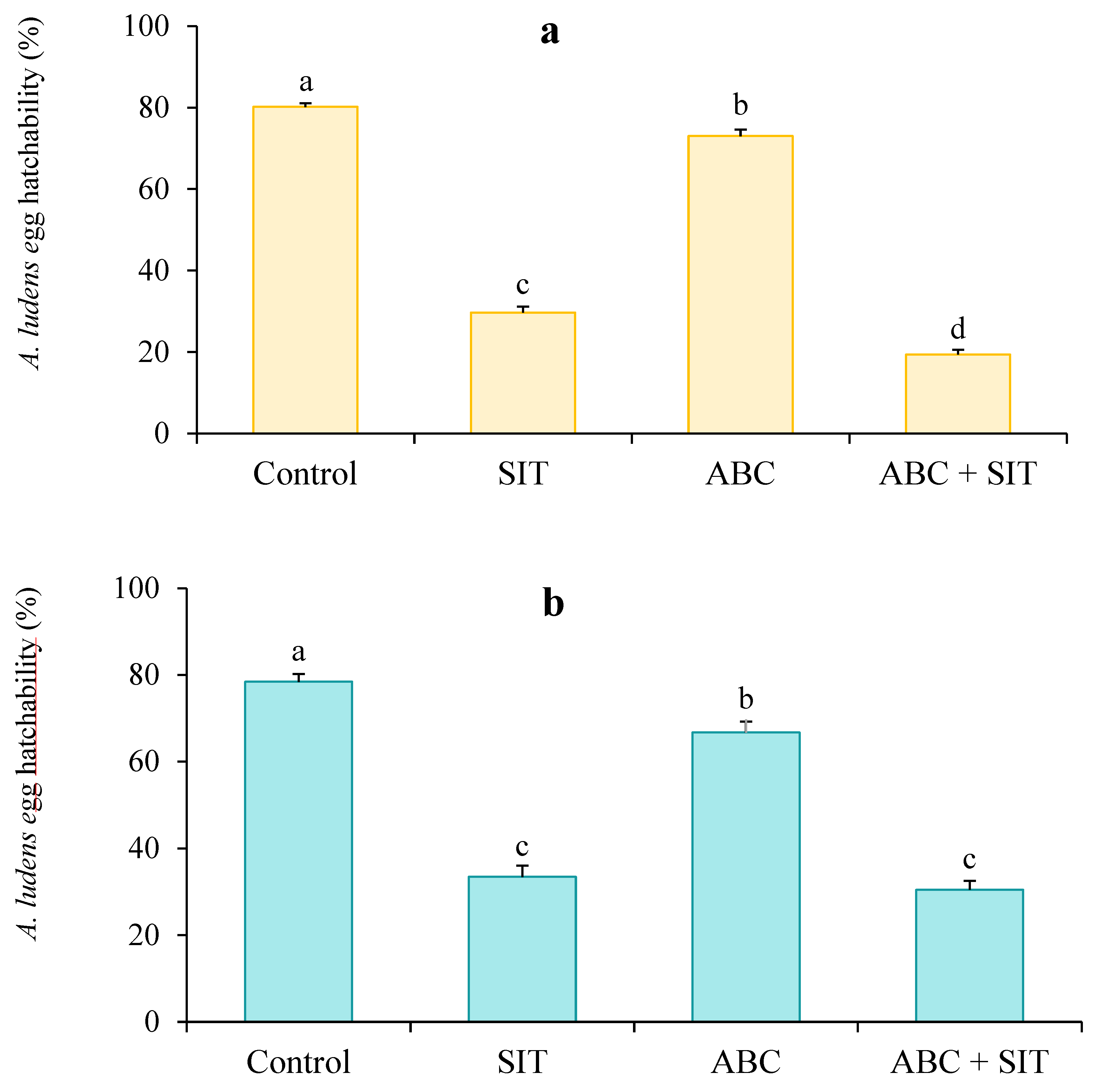{kind=link}
{kind=link}
| Treatments | Emergence (%) in Sentinel Cages | Males Emerged in Field Cages | Sterile Males Released | Rate Sterile: Fertile |
|---|---|---|---|---|
| Assay with D. longicaudata | ||||
| ABC | 85.40 ± 3.18 a | 63.43 ± 10.61 b | -- | -- |
| SIT | 80.40 ± 2.73 a | 238.00 ± 3.39 a | 2250.00 ± 7.90 a | 10:1 |
| ABC + SIT | 80.20 ± 2.47 a | 73.76 ± 16.36 b | 2136.00 ± 57.73 a | 29:1 |
| Control | 82.40 ± 2.73 a | 239.00 ± 7.48 a | -- | -- |
| Assay with C. haywardi | ||||
| ABC | 74.80 ± 6.39 a | 117.74 ± 5.05 b | -- | -- |
| SIT | 75.80 ± 4.27 a | 193.00 ± 11.68 a | 1896.0 ± 106.8 a | 10:1 |
| ABC + SIT | 85.80 ± 3.49 a | 116.31± 18.89 b | 1716.0 ± 69.97 a | 15:1 |
| Control | 82.20 ± 3.48 a | 214.00 ± 15.60 a | -- | -- |
| Treatments | Gross Fertility Rate | Gross Hatch Rate | Eggs per Female per Day | Mean Age Net Fertility (Days) | Intrinsic Rate of Increase (r) |
|---|---|---|---|---|---|
| Assay with D. longicaudata | |||||
| ABC | 80.59 | 73.24 | 0.04 | 46.47 | 0.094 |
| SIT | 145.46 | 30.59 | 1.06 | 47.07 | 0.105 |
| ABC + SIT | 27.34 | 20.37 | −0.29 | 47.53 | 0.069 |
| Control | 424.14 | 80.29 | 3.64 | 46.56 | 0.129 |
| Assay with C. haywardi | |||||
| ABC | 144.30 | 71.41 | 1.01 | 45.63 | 0.108 |
| SIT | 117.00 | 39.84 | 1.09 | 47.18 | 0.100 |
| ABC + SIT | 63.65 | 26.85 | 0.42 | 46.72 | 0.088 |
| Control | 433.05 | 84.21 | 3.67 | 46.17 | 0.131 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montoya, P.; Flores-Sarmiento, E.; López, P.; Ayala, A.; Cancino, J. Additive Effect of Releasing Sterile Insects Plus Biocontrol Agents against Fruit Fly Pests (Diptera: Tephritidae) under Confined Conditions. Insects 2023, 14, 337. https://doi.org/10.3390/insects14040337
Montoya P, Flores-Sarmiento E, López P, Ayala A, Cancino J. Additive Effect of Releasing Sterile Insects Plus Biocontrol Agents against Fruit Fly Pests (Diptera: Tephritidae) under Confined Conditions. Insects. 2023; 14(4):337. https://doi.org/10.3390/insects14040337
Chicago/Turabian StyleMontoya, Pablo, Erick Flores-Sarmiento, Patricia López, Amanda Ayala, and Jorge Cancino. 2023. "Additive Effect of Releasing Sterile Insects Plus Biocontrol Agents against Fruit Fly Pests (Diptera: Tephritidae) under Confined Conditions" Insects 14, no. 4: 337. https://doi.org/10.3390/insects14040337