
Host-Plant Selection Behavior of Ophraella communa, a Biocontrol Agent of the Invasive Common Ragweed Ambrosia artemisiifolia
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Host Plants
2.2. Insect Culture
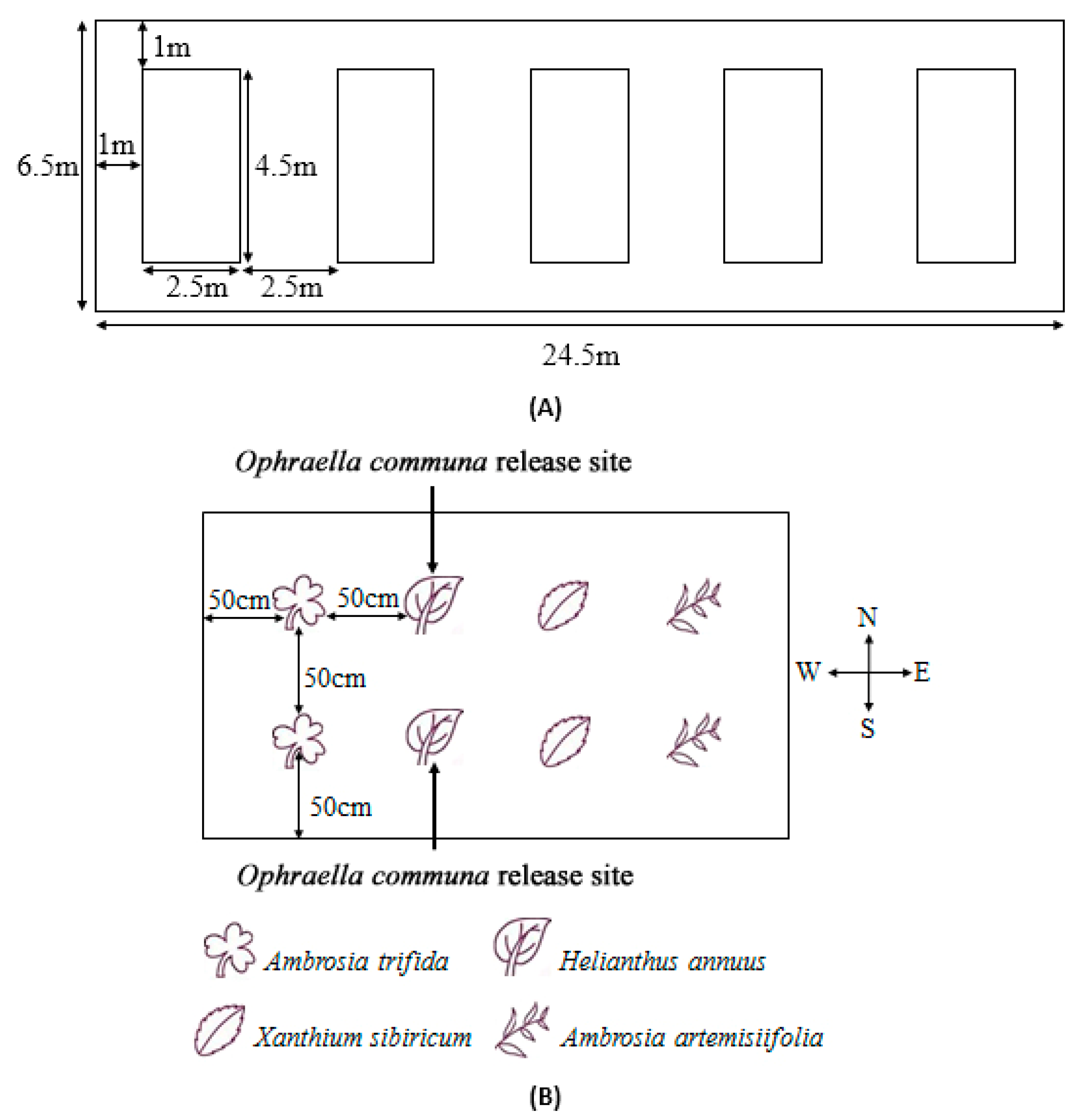
2.3. Distribution and Oviposition Preference Behavior of O. communa Adults on Four Different Coexisting Plant Species in Outdoor Cages
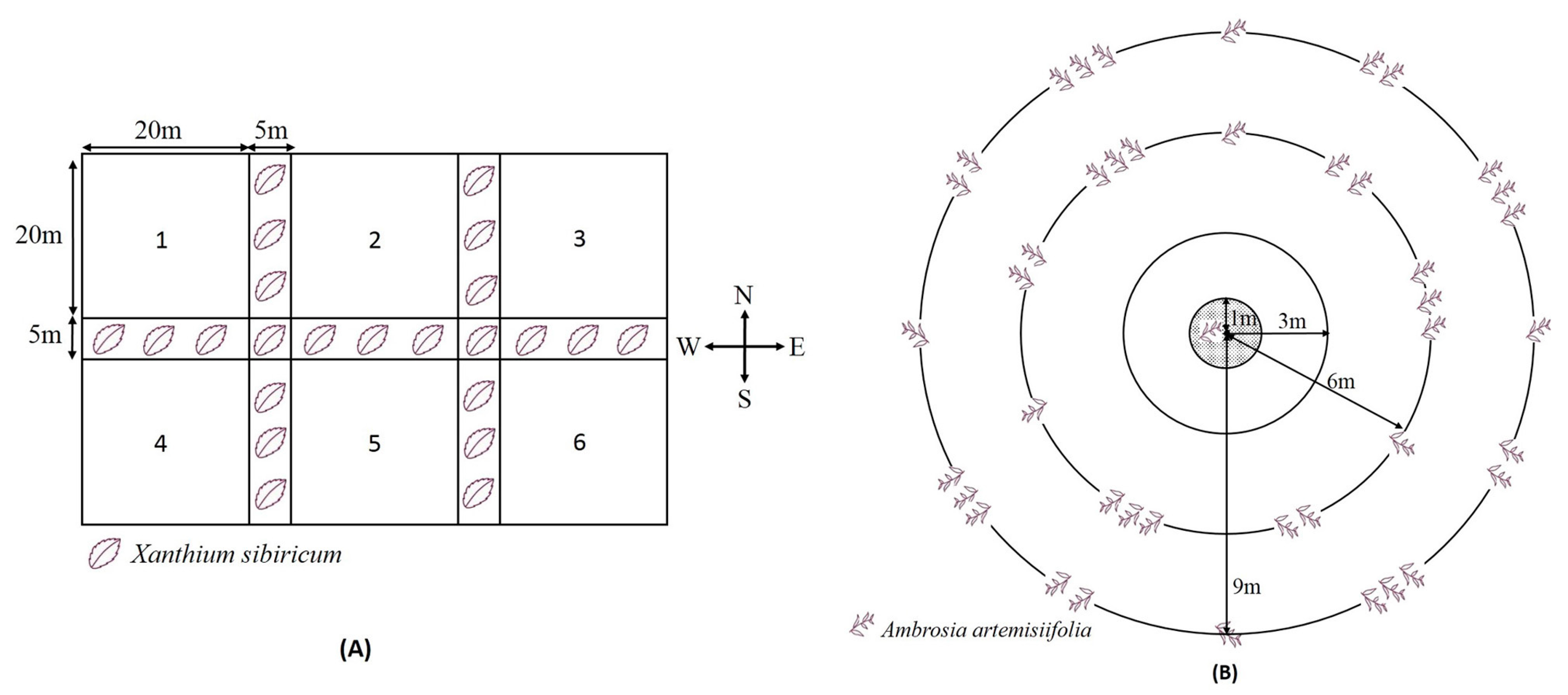
2.4. Host-Plant Selection Behavior of O. communa on Regularly Distributed Ragweed Patches in Sunflower Plots
2.5. Data Analysis
3. Results
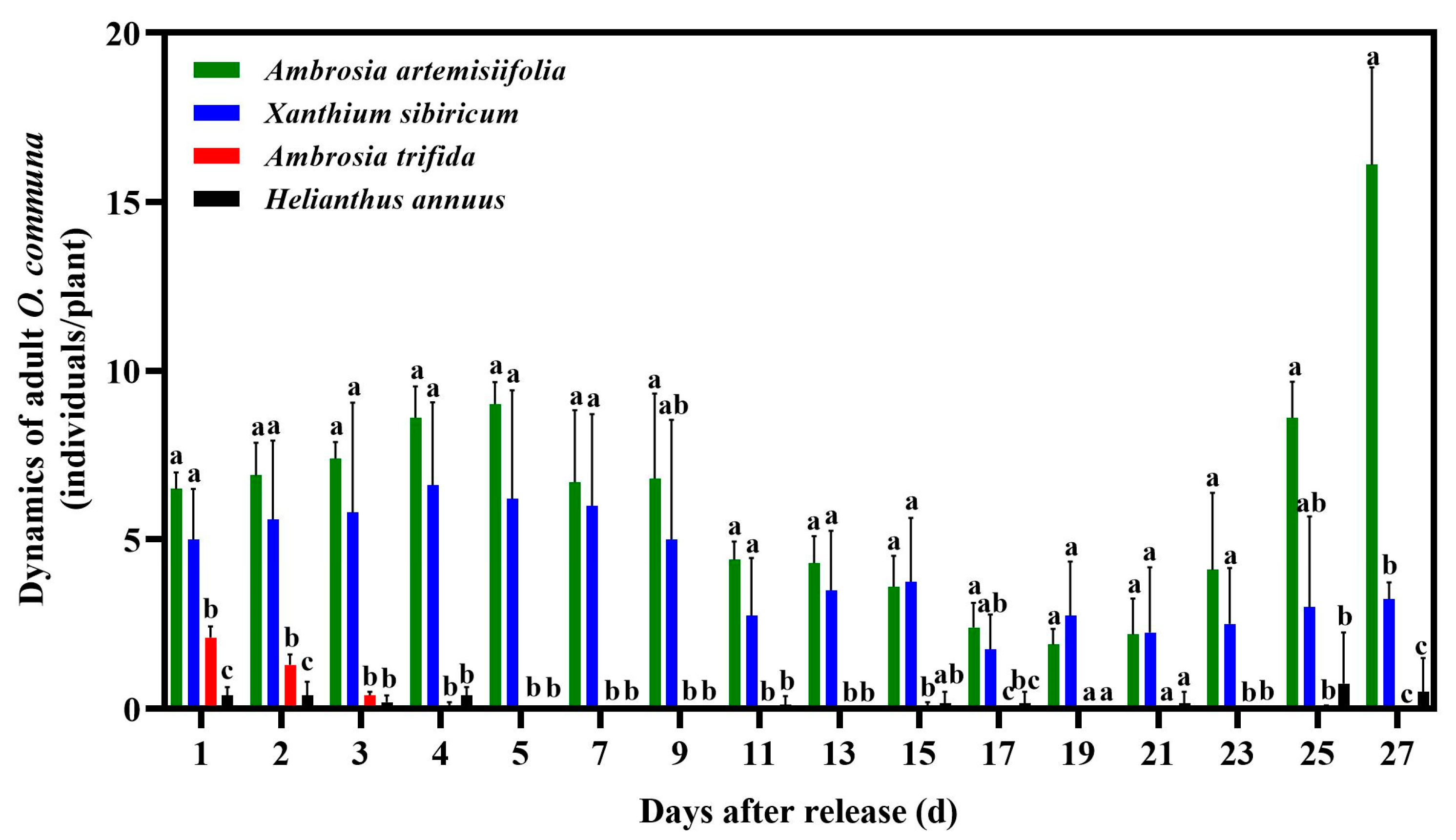
3.1. Distribution of O. communa Adults on Four Different Coexisting Plant Species in Outdoor Cages
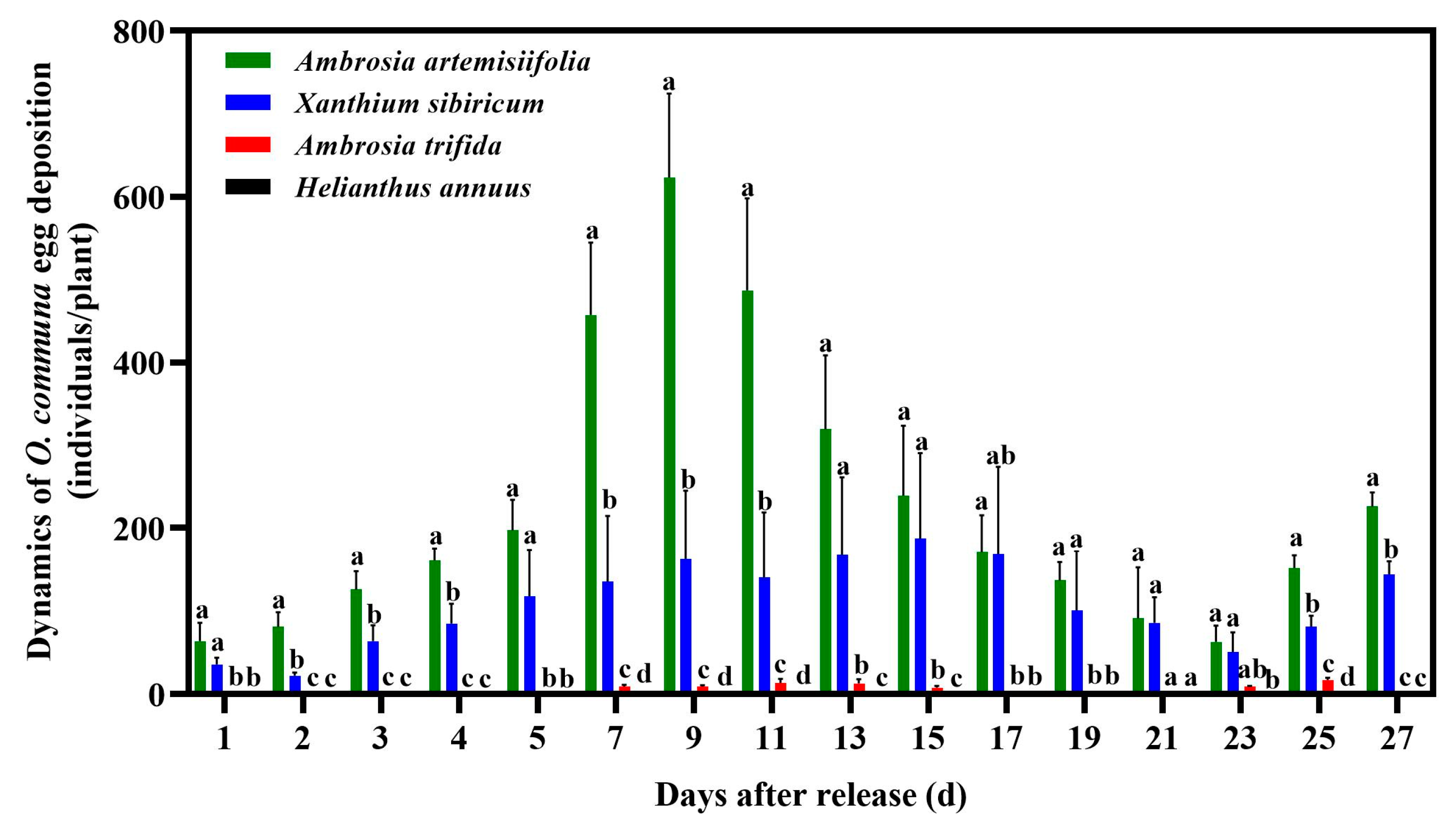
3.2. Oviposition Preference Behavior of O. communa Adults on Four Different Coexisting Plant Species in Outdoor Cages
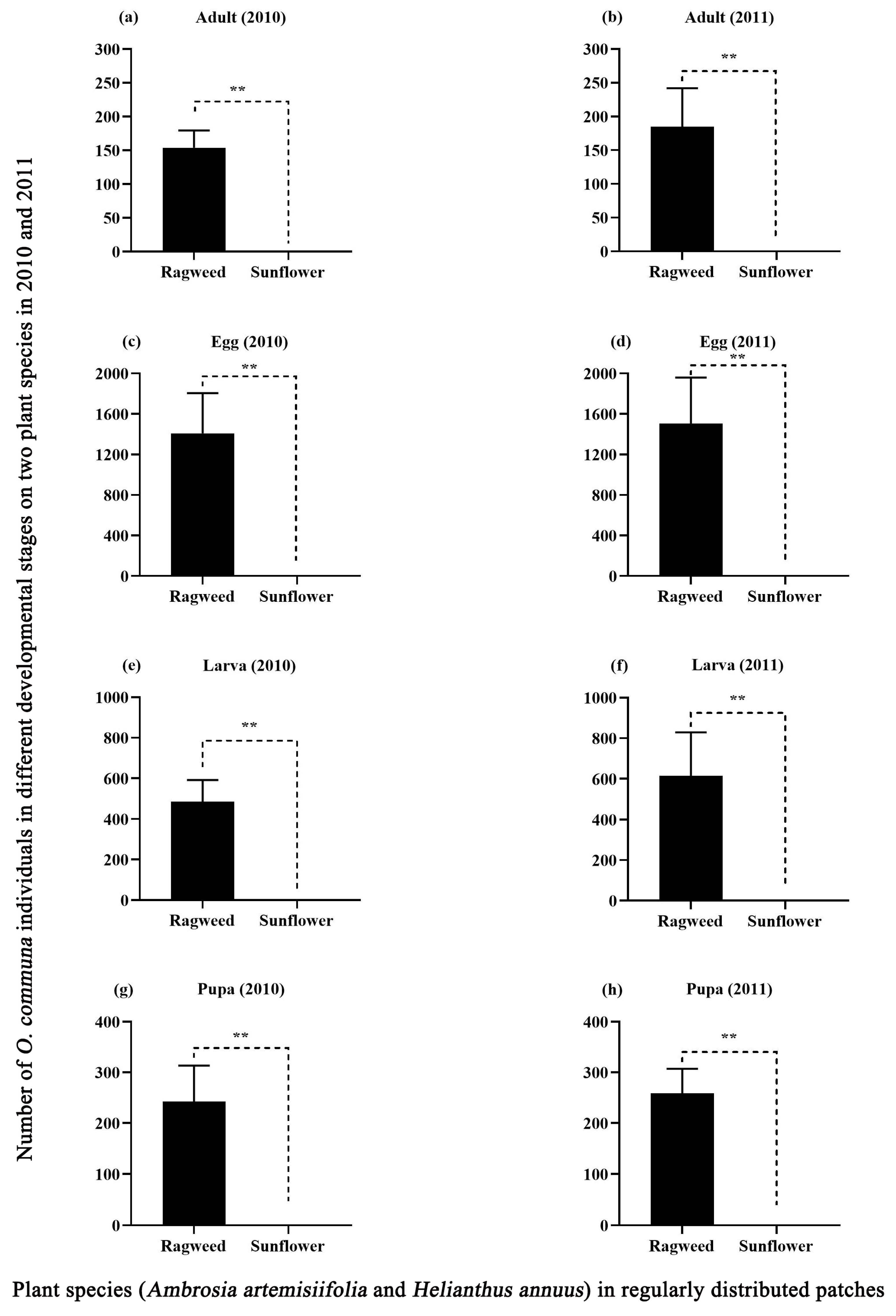
3.3. Host-Plant Selection Behavior of O. communa on Regularly Distributed Ragweed Patches in Sunflower Plots
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heard, T.A. Concepts in insect host-plant selection behaviour and their application to host specificity testing. In Proceedings of Session: Host Specificity Testing of Exotic Arthropod Biological Control Agents: The Biological Basis for Improvement in Safety, Proceedings of the X International Symposium on Biological Control of Weeds, Bozeman, MT, USA, 4–14 July 1999; Van Driesche, R.G., Heard, T.A., McClay, A., Reardon, R., Eds.; USDA Forest Service, Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2000; pp. 1–10. [Google Scholar]
- Augustinus, B.A.; Gentili, R.; Horvath, D.; Naderi, R.; Sun, Y.; Tournet, A.M.T.E.; Müller-Schärer, H. Assessing the risks of non-target feeding by the accidentally introduced ragweed leaf beetle, Ophraella communa, to native European plant species. Biol. Control 2020, 150, 104356. [Google Scholar] [CrossRef]
- Müller-Schärer, H.; Bouchemousse, S.; Litto, M.; McEvoy, P.B.; Roderick, G.K.; Sun, Y. How to better predict long-term benefits and risks in weed biocontrol: An evolutionary perspective. Curr. Opin. Insect Sci. 2020, 38, 84–91. [Google Scholar] [CrossRef]
- Cullen, J.M. Current problems in host-specificity screening. In Proceedings of the VII International Symposium on Biological Control of Weeds, Rome, Italy, 6–11 March 1988; Delfosse, E.S., Ed.; Istituto Sperimentale per la Patologia Vegetale: Rome, Italy, 1990; pp. 27–36. [Google Scholar]
- Marohasy, J. The design and interpretation of host specificity tests for weed biological control with particular reference to insect behavior. Biocontrol. News Inf. 1998, 19, 13–20. [Google Scholar]
- Urs, S. Host range testing of insects for biological weed control: How can it be better interpreted? Bioscience 2001, 51, 951–959. [Google Scholar]
- Kuhlmann, U.; Mason, P.G.; Hinz, H.L.; Blossey, B.; De Clerck-Floate, R.A.; Dosdall, L.M.; McCaffrey, J.P.; Schwarzlaender, M.; Olfert, O.; Brodeur, J.; et al. Avoiding conflicts between insect and weed biological control: Selection of non-target species to assess host specificity of cabbage seedpod weevil parasitoids. J. Appl. Entomol. 2006, 130, 129–141. [Google Scholar] [CrossRef]
- Melanie, L.H.; Rowan, M.E.; Andy, S.; Pauline, S.; Toni, M.W.; Susan, P.W. Impliations of individual variation in insect behavior for host specificity testing in weed. Biocontrol 2013, 58, 703–713. [Google Scholar]
- Lu, Y.H.; Zhang, Y.J.; Wu, K.M. Host-plant selection mechanisms and behavioural manipulation strategies of phytophagous insects. Acta Ecol. Sin. 2008, 10, 5113–5122. [Google Scholar]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [Green Version]
- Finch, S.; Collier, R.H. Host-plant selection by insects-a theory based on ‘appropriate/inappropriate landings’ by pest insects of cruciferous plants. Entomol. Exp. Appl. 2000, 96, 91–102. [Google Scholar] [CrossRef]
- Ma, C.; Cui, S.W.; Bai, Q.; Tian, Z.Q.; Zhang, Y.; Chen, G.M.; Gao, X.Y.; Tian, Z.Y.; Chen, H.S.; Guo, J.Y.; et al. Olfactory co-receptor is involved in host recognition and oviposition in Ophraella communa (Coleoptera: Chrysomelidae). Insect Mol. Biol. 2020, 29, 381–390. [Google Scholar] [CrossRef]
- Bonebrake, T.C.; Boggs, C.L.; McNally, I.M.; Ranganathan, J.; Ehrlich, P.R. Oviposition behavior and offspring performance in herbivorous insects: Consequences of climatic and habitat heterogeneity. Oikos 2010, 119, 927–934. [Google Scholar] [CrossRef]
- Gripenberg, S.; Mayhew, P.J.; Parnell, M.; Roslin, T.A. Meta-analysis of preference-performance relationships in phytophagous insects. Ecol. Lett. 2010, 13, 383–393. [Google Scholar] [CrossRef]
- Scheirs, J.; De Bruyn, L. Integrating optimal foraging and optimal oviposition theory in plant-insect research. Oikos 2002, 96, 187–191. [Google Scholar] [CrossRef]
- Poykko, H. Females and larvae of a geometrid moth, Cleorodes lichenaria, prefer a lichen host that assures shortest larval period. Environ. Entomol. 2006, 35, 1669–1676. [Google Scholar] [CrossRef] [Green Version]
- Loewy, K.J.; Murphy, S.M. Trade-offs in host choice by an herbivorous insect based on parasitism and host plant quality. Oecologia 2015, 179, 741–751. [Google Scholar]
- Withers, T.M.; Mansfield, S. Choice or no-choice tests? Effects of experimental design on the expression of host ranges. In Proceedings of the Second International Symposium on Biological Control of Arthropods, Davos, Switzerland, 12–16 September 2005; pp. 620–633. [Google Scholar]
- Litto, M.; Bouchemousse, S.; Schaffner, U.; Müller-Schärer, H. Population differentiation in response to temperature in Ophraella communa: Implication for the biological control of Ambrosia artemisiifolia. Biol. Control 2021, 164, 104777. [Google Scholar] [CrossRef]
- Murray, T.J.; Withers, T.M.; Mansfield, S. Choice versus no-choice test interpretation and the role of biology and behavior in parasitoid host specificity tests. Biol. Control 2010, 52, 153–159. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Wu, K.; Ding, J.Q. Host specificity of Euops chinensis, a potential biological control agent of Fallopia japonica, an invasive plant in Europe and North America. BioControl 2010, 55, 551–559. [Google Scholar] [CrossRef]
- Shepherd, R.C.H. Problems which arise with host-specificity testing of insects. In Proceedings of the Seventh International Symposium on the Biological Control of Weeds, Rome, Italy, 6–11 March 1988; Delfosse, E.S., Ed.; Insituto Sperimentale per la Patologia Vegetale: Rome, Italy, 1990; pp. 85–92. [Google Scholar]
- Harley, K.L.S.; Kassulke, R.C.; Julien, M.H. Biology and host specificity of Microthrix inconspicuella Ragonat (Lepidoptera: Pyralidae), a natural enemy of Emex australis in South Africa. J. Entomol. Soc. South. Afr. 1979, 42, 343–348. [Google Scholar]
- Bernays, E.A.; Chapman, R.F. Host Plant Selection by Phytophagous Insect; Chapman and Hall: London, UK, 1994. [Google Scholar]
- Ukeh, D.A.; Birkett, M.A.; Bruce, T.; Allan, E.J.; Luntz, A.J.M. Behavioural responses of the maize weevil, Sitophilus zeamais, to host (stored-grain) and non-host plant volatiles. Pest Manag. Sci. 2010, 66, 44–50. [Google Scholar] [CrossRef]
- Mandal, P.; Mondal, F.; Hossain, M.S. Factors influences selection and adaptation of aphid to their Host Plant. J. Plant Sci. Crop Protec. 2020, 3, 102. [Google Scholar]
- Szentesi, A. The role of experience in host plant choice by phytophagous insects. In Insect-Plant Interactions (1990); CRC Press: Boca Raton, FL, USA, 1989; p. 2. [Google Scholar]
- Ou, H.Y.; Tian, M.Y.; Lin, R.; He, Y.R.; Xie, M.Q. The effect of host kairomones and learning behavior on olfactory responses of Trichogramma. J. Environ. Entomol. 2010, 32, 243–249. [Google Scholar]
- Furlong, M.J.; Wright, D.J.; Dosdall, L.M. Diamondback moth ecology and management: Problems, progress, and prospects. Annu. Rev. Entomol. 2013, 58, 517–541. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.; Anton, S. Experience-based modulation of behavioural responses to plant volatiles and other sensory cues in insect herbivores. Plant Cell. Environ. 2014, 37, 1826–1835. [Google Scholar] [CrossRef] [Green Version]
- Tapia, D.H.; Silva, A.X.; Ballesteros, G.I.; Figueroa, C.C.; Niemeyer, H.M.; Ramírez, C.C. Differences in learning and memory of host plant features between specialist and generalist phytophagous insects. Anim. Behav. 2015, 106, 1–10. [Google Scholar] [CrossRef]
- Withers, T.M.; Barton, B.L.; Stanley, J. How time-dependent processes can affect the outcome of assays. In Proceedings of Session: Host Specificity Testing of Exotic Arthropod Biological Control Agents: The Biological Basis for Improvement in Safety, Proceedings of the X International Symposium on Biological Control of Weeds, Bozeman, MT, USA, 4–14 July 1999; Van Driesche, R.G., Heard, T.A., McClay, A.S., Reardon, R., Eds.; USDA Forest Service, Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2000; pp. 27–41. [Google Scholar]
- Wan, F.H.; Harris, P.; Cai, L.M.; Zhang, M.X. Host specificity of Altica carduorum Guer. (Chrysomelidae: Coleoptera), a defoliator of Cirsium arvense (L.) Scop. (Asteraceae) from North-western China. Biocontrol Sci. Technol. 1996, 6, 521–530. [Google Scholar] [CrossRef]
- Clement, S.A. Review of open field tests in host-specificity determination of insects for biological control of weeds. Biocontrol Sci. Technol. 2013, 5, 395–406. [Google Scholar] [CrossRef]
- Fukano, Y.; Doi, H. Population abundance and host use pattern of Ophraella communa (Coleoptera: Chrysomelidae) in its native and introduced range. Biocontrol Sci. Technol. 2013, 23, 595–601. [Google Scholar] [CrossRef]
- Kim, H.G.; Lee, D.H. Review of the biology and ecology of a ragweed leaf beetle, Ophraella communa (Coleoptera: Chrysomelidae), which is a biological control agent of an invasive common ragweed, Ambrosia artemisiifolia (Asterales: Asteraceae). Biocontrol Sci. Technol. 2019, 29, 185–200. [Google Scholar] [CrossRef]
- Zhou, Z.S.; Chen, H.S.; Zheng, X.W.; Guo, J.Y.; Guo, W. Control of the invasive weed Ambrosia artemisiifolia with Ophraella communa and Epiblema strenuana. Biocontrol Sci. Technol. 2014, 24, 950–964. [Google Scholar] [CrossRef]
- Hu, Y.P.; Meng, L. Potential impacts of alien herbivorous insect Ophraella communa (Coleoptera: Chrysomelidae) on non-target plants in mainland China. Chinese J. Ecol. 2007, 26, 56–60. [Google Scholar]
- Watanabe, M.; Hirai, Y. Host-use pattern of the ragweed beetle Ophraella communa Lesage (Coleoptera: Chrysomelidae) for overwintering and reproduction in Tsukuba. Appl. Entomol. Zool. 2004, 39, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.S.; Oh, K.S.; Lee, Y.D.; Lee, S.Y.; Lee, H.; Kim, H.J.; Kim, D.E. The distribution of the exotic species, Ophraella communa LeSage (Coleoptera, Chrysomelidae) and their applicability for biological control against ragweed Ambrosia artemisiifolia L. on Jeju Island. Environ. Biol. Res. 2017, 35, 437–445. [Google Scholar] [CrossRef]
- Palmer, W.A.; Goeden, R.D. The host of range Ophraella communa Lesage (Coleoptera: Chrysomelidae). Coleopts Bull. 1991, 45, 115–120. [Google Scholar]
- Dernovici, S.A.; Teshler, M.P.; Watson, A.K. Is Sunflower (Helianthus annuus) at risk to damage from Ophraella communa, a natural enemy of common ragweed (Ambrosia artemisiifolia)? Biocontrol Sci. Technol. 2006, 16, 669–686. [Google Scholar] [CrossRef]
- Cao, Z.J.; Wang, H.Y.; Meng, L.; Li, B.P. Risk to nontarget plants from Ophraella communa (Coleoptera: Chrysomelidae), a potential biological control agent of alien invasive weed Ambrosia artemisiifolia (Asteraceae) in China. Appl. Entomol. Zool. 2001, 46, 201–210. [Google Scholar] [CrossRef]
- Zhou, Z.S.; Guo, J.Y.; Zheng, X.W.; Luo, M.; Chen, H.S.; Wan, F.H. Reevaluation of biosecurity of Ophraella communa against sunflower (Helianthus annuus). Biocontrol Sci. Technol. 2011, 21, 1147–1160. [Google Scholar] [CrossRef]
- Müller-Schärer, H.; Lommen, S.T.; Rossinelli, M.; Bonini, M.; Boriani, M.; Bosio, G.; Schaffner, U. Ophraella communa, the ragweed leaf beetle, has successfully landed in Europe: Fortunate coincidence or threat? Weed Res. 2014, 54, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Wan, F.H.; Wang, R. A review on biological control of the ragweed, Ambrosia artemisiifolia. Weed Sci. 1990, 1, 30–32. [Google Scholar]
- Komives, T.; Beres, I.; Reisinger, P. New strategy of the integrated protection against common ragweed (Ambrosia artemisiifolia L.). Hung Weed Res. Technol. 2006, 6, 5–50. [Google Scholar]
- Jin, L.Y.; Yu, J.Y.; Guo, Q.Y.; Ma, S.J.; Dong, J.G.; Yang, J.; Zhang, L.H. Characteristics of weed community and optimization of nicosulfuron and atrazine combination in spring corn field in eastern Hebei province. J. Maize Sci. 2022, 30, 157–163. [Google Scholar]
- Schoonhoven, L.M.; van Loon, J.J.A.; Dicke, M. Insect-Plant Biology, 2nd ed.; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Chu, S.; Cong, S.; Li, R.; Hou, Y. Host range of Herpetogramma basalis (Lepidoptera: Crambidae), a biological control agent for the invasive weed Alternanthera philoxeroides (Centrospermae: Amaranthaceae) in China. J. Insect Sci. 2019, 19, 1–7. [Google Scholar] [CrossRef]
- Jaenike, J. Optimal oviposition behavior in phytophagous insects. Theor. Popul. Biol. 1978, 14, 350–356. [Google Scholar] [CrossRef]
- Liu, Q.S.; Hu, X.Y.; Su, S.L.; Peng, Y.F.; Li, Y.H. Cooperative herbivory between two important pests of rice. Nat. Commun. 2021, 12, 6772. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Li, B.P.; Meng, L. Host-selection behavior of exotic Ophraella communa (Chrysomelidae: Coleoptera) for non-target plants. Chinese J. Biol. Control 2009, 25, 120–124. [Google Scholar]
- Yamanaka, T.; Tanaka, K.; Otuka, A.; Bjørnstad, O.N. Detecting spatial interactions in the ragweed (Ambrosia artemissifolia L.) and the ragweed beetle (Ophraella communa LeSage) populations. Ecol. Res. 2007, 22, 185–196. [Google Scholar] [CrossRef]
- Bruce, T.J.; Wadhams, L.J.; Woodcock, C.M. Insect host location: A volatile situation. Trends Plant Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef]
- Bruce, T.J.; Pickett, J.A. Perception of plant volatile blends by herbivorous insects–finding the right mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef]
- Guarino, S.; Arif, M.A.; Millar, J.G.; Colazza, S.; Peri, E. Volatile unsaturated hydrocarbons emitted by seedlings of brassica species provide host location cues to Bagrada hilaris. PloS ONE 2018, 13, e0209870. [Google Scholar] [CrossRef]
- Arif, M.A.; Guarino, S.; Peri, E.; Colazza, S. Evaluation of brassicaceae seedlings as trap plants for Bagrada hilaris Burmeister in caper bush cultivations. Sustainability 2020, 12, 6361. [Google Scholar] [CrossRef]
- Mappes, J.; Kaitala, A. Host-plant selection and predation risk for offspring of the parent bug. Ecology 1995, 76, 2668–2670. [Google Scholar] [CrossRef]
- Ballabeni, P.; Wlodarczyk, M.; Rahier, M. Does enemy-free space for eggs contribute to a leaf beetle’s oviposition preference for a nutritionally inferior host plant? Funct. Ecol. 2001, 15, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Why, A.M.; Walton, W.E. Ovipositional Responses of Mosquitoes to Kairomones Produced from Aquatic Taxa in Different Guilds. In Proceedings of the Entomological Society of America Annual Meeting, Online, 13–16 November 2011. [Google Scholar]
- Hirayama, H.; Kasuya, E. Effect of adult females’ predation risk on oviposition site selection in a water strider. Entomol. Exp. Appl. 2013, 149, 250–255. [Google Scholar] [CrossRef]
- Miyatake, T.; Ohno, T. Seasonal abundance of exotic leaf beetle Orphraella communa LeSage (Coleoptera: Chrysomelidae) on two different host plants. Appl. Entomol. Zool. 2010, 45, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Meng, L.; Li, B.P. Host choice and leaf consumption of Ophraella communa (Coleoptera: Chrysomelidae) on Ambrosia artemisiifolia and Xanthium sibiricum. Chin. J. Ecol. 2011, 30, 1337–1341. [Google Scholar]
- Qian, X. The preliminary studies on habits of growth and damage of cocklebur and its chemical control. Soybean Sci. 1988, 1, 61–68. [Google Scholar]
- Zhang, C.Q.; Chen, R.H.; Feng, X.H.; Cheng, X.Q.; Zhang, G.P.; Fan, Z.L. Investigation on species and distribution of weeds in tobacco fields in Jiangxi province. Acta Agric. Jiangxi 2012, 24, 80–82. [Google Scholar]
- Wang, Y.; Huang, C.Y.; Piao, D.W.; Huang, Y.J. Weed survey of sunflower field in northwest region of Heilongjiang province. Heilongjiang Agric. Sci. 2013, 6, 29–32. [Google Scholar]
- Funk, D.; Futuyma, D.; Orti, G.; Meyer, A. A history of host associations and evolutionary diversification for Ophraella (Coleoptera: Chrysomelidae): New evidence from mitochondrial DNA. Evolution 1995, 49, 1008–1017. [Google Scholar] [CrossRef] [Green Version]
- Fukano, Y.; Doi, H.; Thomas, C.E.; Takata, M.; Koyama, S.; Satoh, T. Contemporary evolution of host plant range expansion in an introduced herbivorous beetle Ophraella communa. J. Evol. Biol. 2016, 29, 757–765. [Google Scholar] [CrossRef] [Green Version]
- Nishide, Y.; Fukano, Y.; Doi, H.; Satoh, T.; Inoue, H.; Boriani, M. Origins and genetic diversity of the ragweed beetles, Ophraella communa (Coleoptera: Chrysomelidae), that were introduced into Italy and Japan based on an analysis of mitochondrial DNA sequence data. Eur. J. Entomol. 2015, 112, 613–618. [Google Scholar] [CrossRef] [Green Version]
- Root, R.B. Organization of a plant-arthropod association in simple and diverse habitats: The fauna of collards (Brassica oleracea). Ecol. Monogr. 1973, 43, 95–124. [Google Scholar] [CrossRef]
- Long, Z.T.; Mohler, C.L.; Carson, W.P. Extending the resource concentration hypothesis to plant communities: Effects of litter and herbivores. Ecology 2003, 84, 652–665. [Google Scholar] [CrossRef] [Green Version]
- Ohsaki, H.; Miyagi, A.; Kawai-Yamada, M.; Yamawo, A. Intraspecific interaction of host plants leads to concentrated distribution of a specialist herbivore through metabolic alterations in the leaves. Funct. Ecol. 2022, 36, 779–793. [Google Scholar] [CrossRef]
- Grez, A.A.; Gonzalez, R.H. Resource concentration hypothesis: Effect of host plant patch size on density of herbivorous insects. Oecologia 1995, 103, 471–474. [Google Scholar] [CrossRef]
- Bergelson, J.; Kareiva, P. Barriers to movement and the response of herbivores to alternative cropping patterns. Oecologia 1987, 71, 457–460. [Google Scholar] [CrossRef]
- Capman, W.C.; Batzli, G.O.; Simms, L.E. Responses of the common sooty wing skipper to patches of host plants. Ecology 1990, 71, 1430–1440. [Google Scholar] [CrossRef]
- Meng, L.; Xu, J.; Li, H. Dispersal and bionomics of the alien Ophraella communa in China mainland. Chinese J. Biol. Control 2007, 23, 5–10. [Google Scholar]
- Zhou, Z.S.; Guo, J.Y.; Michaud, J.P.; Li, M.; Wan, F.H. Variation in cold hardiness among geographic populations of the ragweed beetle, Ophraella communa LeSage (Coleoptera: Chrysomelidae), a biological control agent of Ambrosia artemisiifolia L. (Asterales: Asteraceae), in China. Biol. Invasions 2011, 13, 659–667. [Google Scholar] [CrossRef]
- Tanaka, K.; Murata, K.; Matsuura, A. Rapid evolution of an introduced insect Ophraella communa LeSage in new environments: Temporal changes and geographical differences in photoperiodic response. Entomol. Sci. 2015, 18, 104–112. [Google Scholar] [CrossRef]
- Sun, Y.; Müller-Schärer, H.; Schaffner, U. Fighting neobiota with neobiota: Consider it more often and do it more rigorously. Biol. Conserv. 2022, 268, 109506. [Google Scholar] [CrossRef]

 ” shows the sites and densities per cluster of common ragweed planted in the 6 m and 9 m radius homocentric rings in each plot.
” shows the sites and densities per cluster of common ragweed planted in the 6 m and 9 m radius homocentric rings in each plot.
” shows the sites and densities per cluster of common ragweed planted in the 6 m and 9 m radius homocentric rings in each plot.
” shows the sites and densities per cluster of common ragweed planted in the 6 m and 9 m radius homocentric rings in each plot.



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, J.; Zhao, M.; Zhou, Z.; Wang, R.; Guo, J.; Wan, F. Host-Plant Selection Behavior of Ophraella communa, a Biocontrol Agent of the Invasive Common Ragweed Ambrosia artemisiifolia. Insects 2023, 14, 334. https://doi.org/10.3390/insects14040334
Jin J, Zhao M, Zhou Z, Wang R, Guo J, Wan F. Host-Plant Selection Behavior of Ophraella communa, a Biocontrol Agent of the Invasive Common Ragweed Ambrosia artemisiifolia. Insects. 2023; 14(4):334. https://doi.org/10.3390/insects14040334
Chicago/Turabian StyleJin, Jisu, Meiting Zhao, Zhongshi Zhou, Ren Wang, Jianying Guo, and Fanghao Wan. 2023. "Host-Plant Selection Behavior of Ophraella communa, a Biocontrol Agent of the Invasive Common Ragweed Ambrosia artemisiifolia" Insects 14, no. 4: 334. https://doi.org/10.3390/insects14040334