
Scanning Electron Microscopy of Antennae and Mouthparts of Mezira yunnana Hsiao (Hemiptera: Aradidae): Specialized Microstructures Reflecting Adaptation to Mycetophagy

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects Collecting
2.2. Samples for SEM
2.3. Feeding Behavior
2.4. Image Processing and Terminology
3. Results
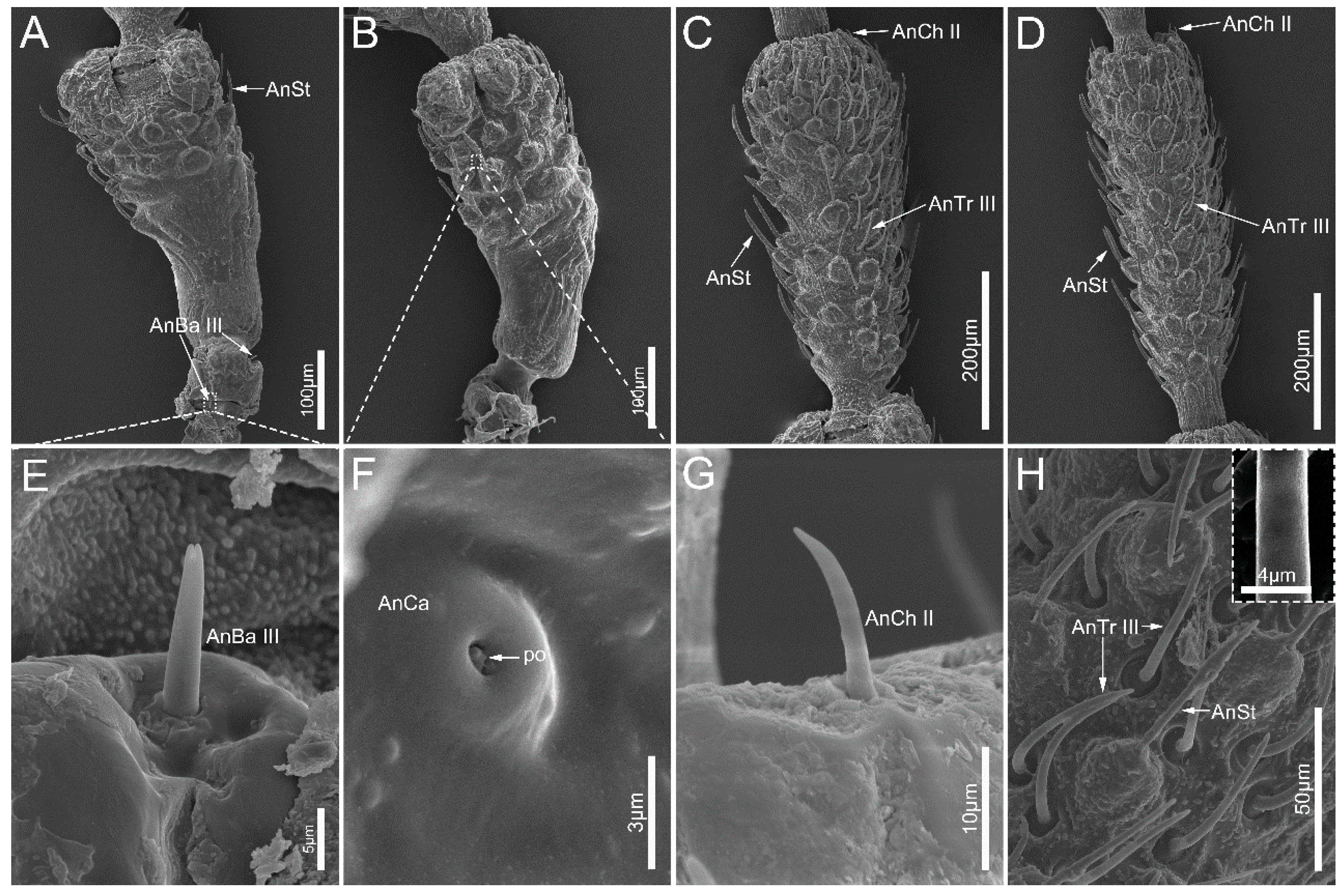
3.1. Antennae
3.2. Mouthparts
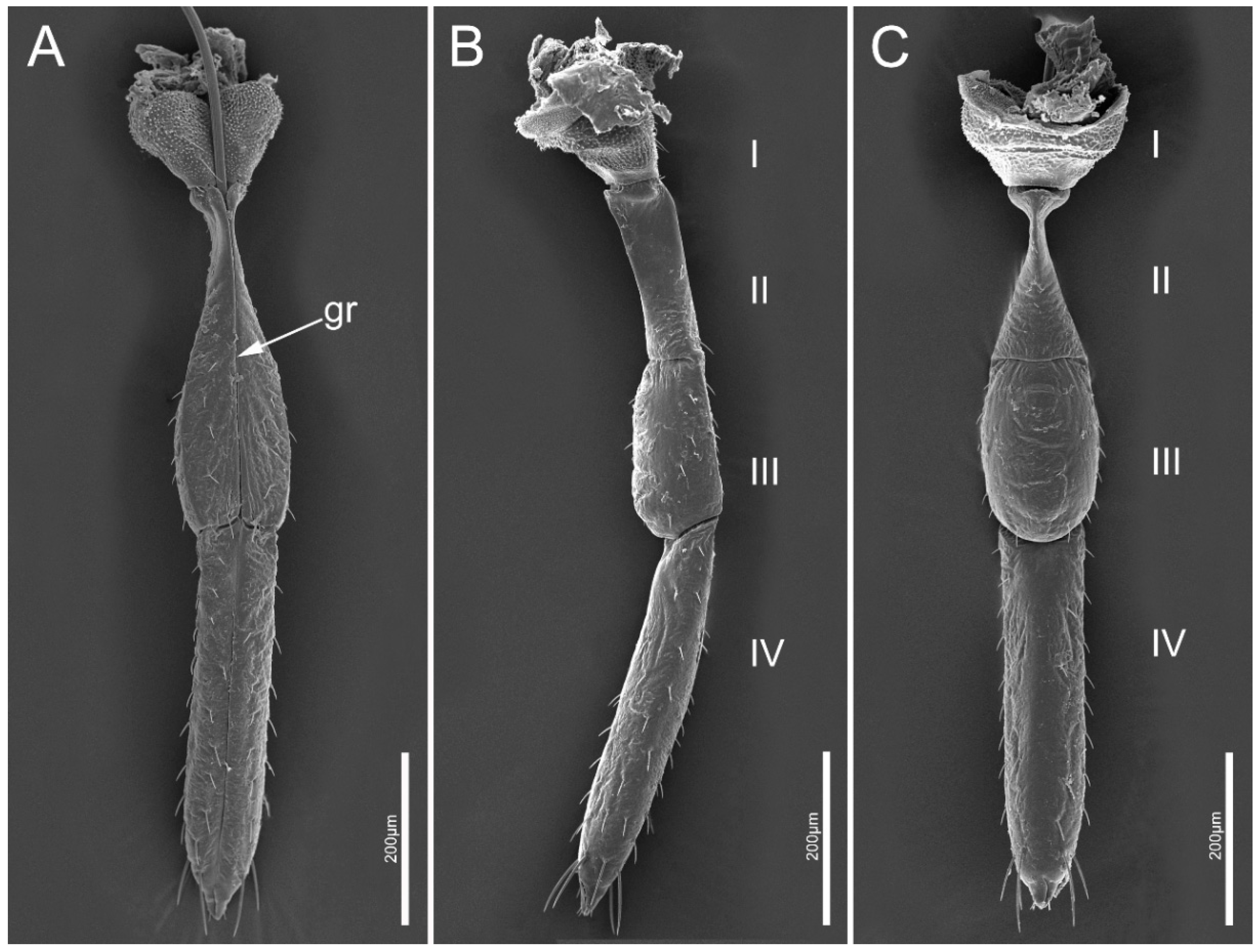
3.2.1. Gross Morphology of Mouthparts
3.2.2. Labium
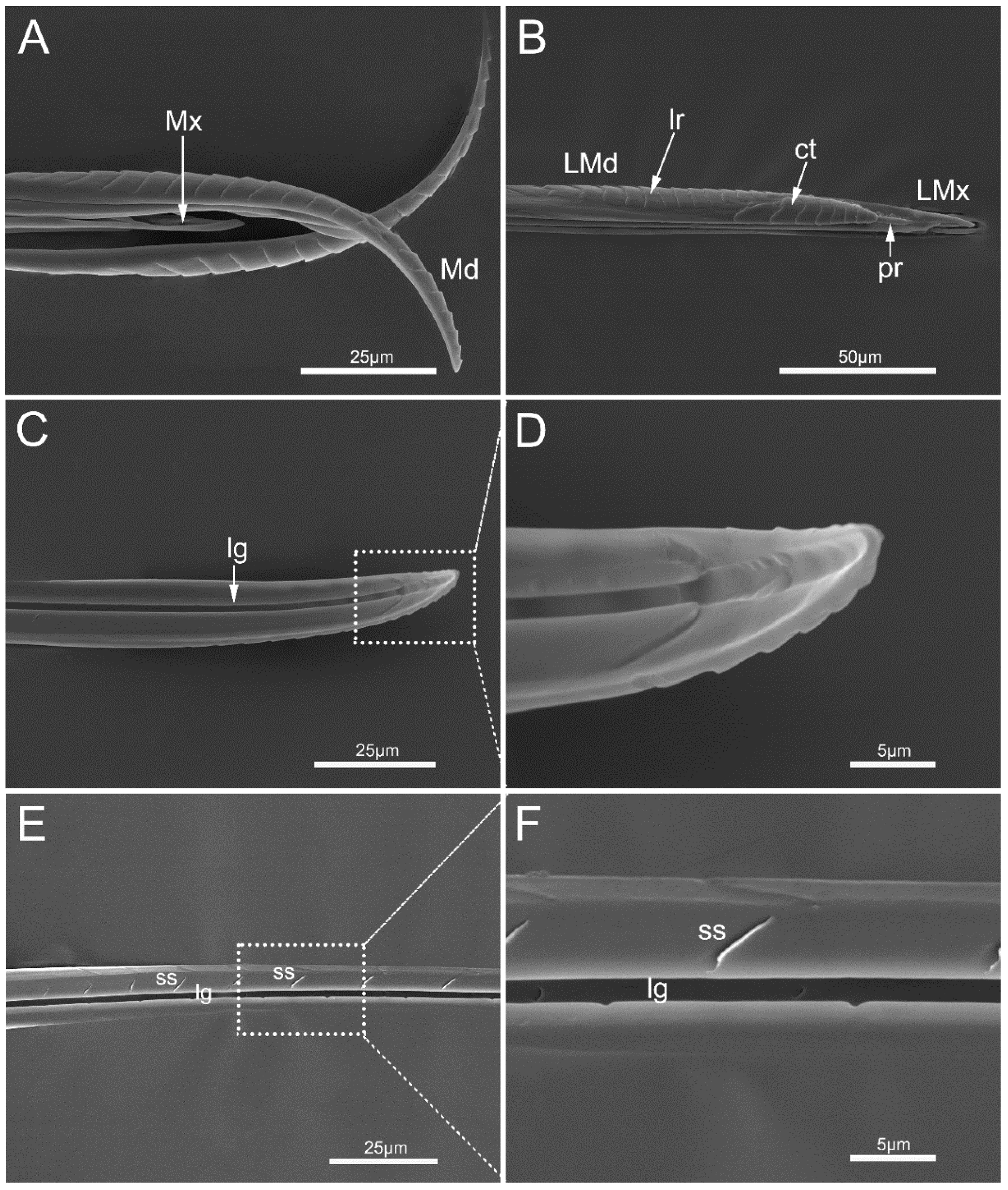
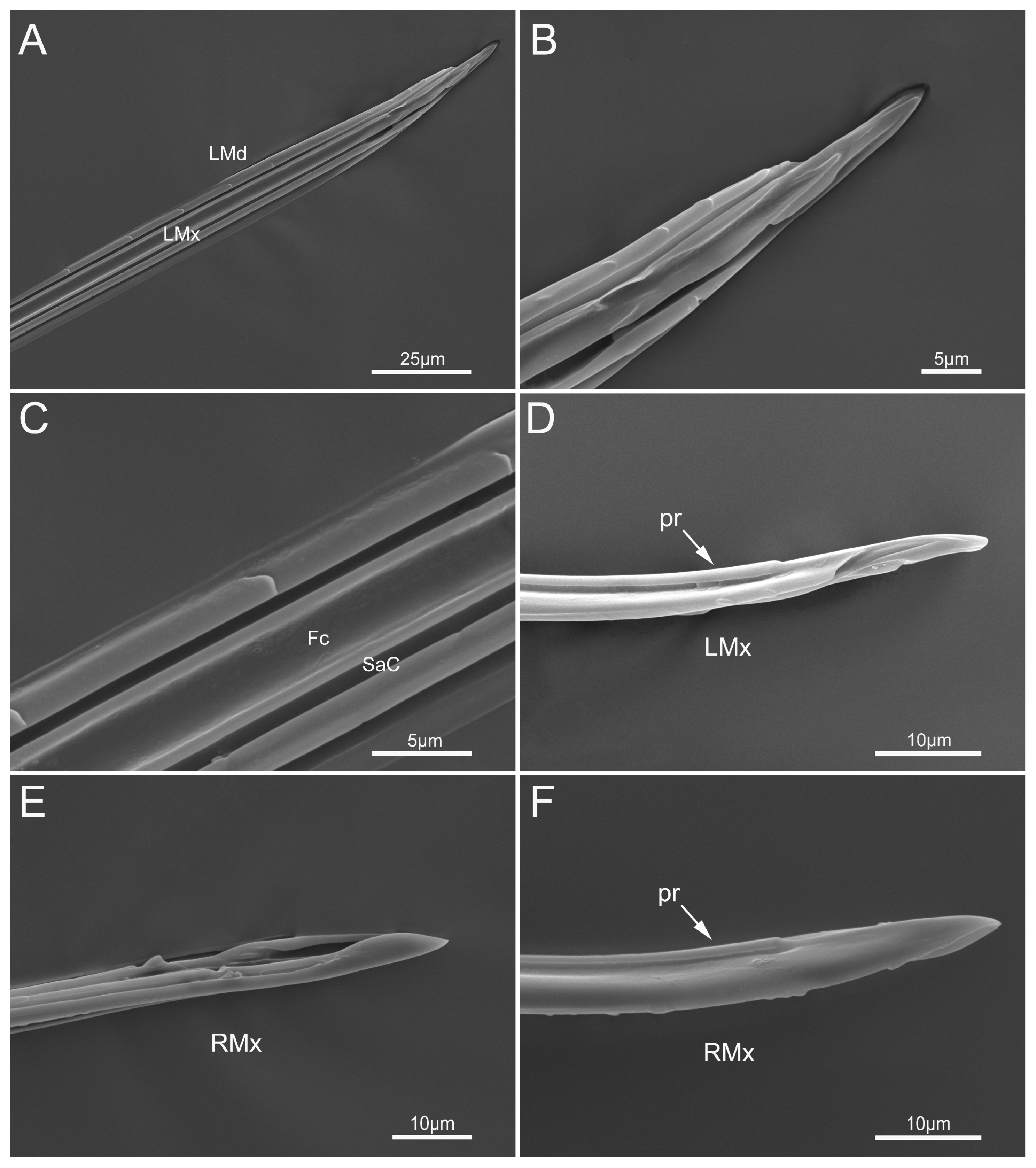
3.2.3. Stylet Fascicle
3.3. Feeding Processes by M. yunnana
4. Discussion
4.1. Specialized Arrangement of Antennal Sensilla in Flat Bugs
4.2. Unique Morphology of Mouthparts Adapted to Fungal Diet

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | AnCh | AnTr | AnBa | AnCa | AnSt | References |
|---|---|---|---|---|---|---|
| Aradus betulae (Linnaeus) | + | + | + | − | + | [19] |
| Aradus betulae (Linnaeus) | + | + | + | − | + | [19] |
| Aradus corticalis (Linnaeus) | + | + | + | − | + | [19,40] |
| Aradus corticalis (Linnaeus) | + | + | + | − | + | [19,40] |
| Aradus depressus (Fabricius) | + | + | + | + | + | [19] |
| Brachyrhynchus membranaceus (Fabricius) | + | + | + | + | + | [19] |
| Mezira yunnana Hsiao | + | + | + | + | + | This study |
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cobben, R.H. Evolutionary Trends in Heteroptera. Part 2. Mouth Part Structures and Feeding Strategies; Veeman, H., Zonen, B.V., Eds.; Mededlingen Landbouwhogeschool: Wageningen, The Netherlands, 1978; pp. 1–407. [Google Scholar]
- Ambrose, D.P.; Livingstone, D. On the bioecology of Lophocephala guerini Lap. (Reduviidae: Harpactorinae) a coprophagous reduviid from the Palghat Gap, India. J. Nat. Hist. 1979, 13, 581–588. [Google Scholar] [CrossRef]
- Cohen, A.C. Feeding adaptations of some predaceous Hemiptera. Ann. Entomol. Soc. Am. 1990, 83, 1215–1223. [Google Scholar] [CrossRef]
- Li, H.; Leavengood, J.M., Jr.; Chapman, E.G.; Burkhardt, D.; Song, F.; Jiang, P.; Liu, J.; Zhou, X.; Cai, W. Mitochondrial phylogenomics of Hemiptera reveals adaptive innovations driving the diversification of true bugs. Proc. Biol. Sci. 2017, 284, 20171223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Tian, L.; Li, H.; Cai, W. Ultrastructural variations of antennae and labia are associated with feeding habit shifts in stink bugs (Heteroptera: Pentatomidae). Biology 2021, 10, 1161. [Google Scholar] [CrossRef]
- Brézot, P.; Tauban, D.; Renou, M. Sense organs on the antennal flagellum of the green stink bug, Nezara viridula (L.) (Heteroptera: Pentatomidae) sensillum types and numerical growth during the post-embryonic development. Int. J. Insect Morphol. Embryol. 1997, 25, 427–441. [Google Scholar] [CrossRef]
- Brożek, J. Internal structure of the mouthparts of true bugs (Hemiptere: Heteroptera). Pol. J. Entomol. 2013, 73, 79–106. [Google Scholar]
- Carbajal, A.L.; Catala, S. Relationship between antennal sensilla pattern and habitat in six species of Triatominae. Memórias Do Inst. Oswaldo Cruz 2002, 97, 1121–1125. [Google Scholar] [CrossRef] [Green Version]
- Ventura, M.U.; Panizzi, A.R. Morphology of olfactory sensilla and its role in host plant recognition by Neomegalotomus parvus (Westwood) (Heteroptera: Alydidae). Braz. Arch. Biol. Technol. 2005, 48, 589–597. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Brozek, J.; Dai, W. Sensory armature and stylets of the mouthparts of Stephanitis nashi (Hemiptera: Cimicomorpha: Tingidae), their morphology and function. Micron 2020, 132, 102840. [Google Scholar] [CrossRef]
- Brożek, J. Morphology and arrangement of the labial sensilla of the water bugs. Bull. Insectol 2008, 61, 67–168. [Google Scholar]
- China, W.E. Morphological parallelism in the structure of the labium in the Hemipterous genera Coptosomoides, gen. nov., and Bozius, Dist. (Fam. Plataspidæ), in connection with mycetophagous habits. Ann. Mag Nat. Hist 2009, 7, 281–286. [Google Scholar] [CrossRef]
- Wang, Y.; Dai, W. Fine structure of mouthparts and feeding performance of Pyrrhocoris sibiricus Kuschakevich with remarks on the specialization of sensilla and stylets for seed feeding. PLoS ONE 2017, 12, e0177209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Dai, W. How does the intricate mouthpart apparatus coordinate for feeding in the hemimetabolous insect pest Erthesina fullo? Insects 2020, 11, 503. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, J.; Wang, W.; Brozek, J.; Dai, W. Unique fine morphology of mouthparts in Haematoloecha nigrorufa (Stål) (Hemiptera: Reduviidae) adapted to millipede feeding. Insects 2020, 11, 386. [Google Scholar] [CrossRef] [PubMed]
- Parveen, S.; Ahmad, A.; Brozek, J.; Ramamurthy, V.V. Morphological diversity of the labial sensilla of phytophagous and predatory Pentatomidae (Hemiptera: Heteroptera), with reference to their possible functions. Zootaxa 2015, 4039, 359–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, A.; Parveen, S.; Brożek, J.; Dey, D. Antennal sensilla of phytophagous and predatory pentatomids (Hemiptera: Pentatomidae): A comparative study of four genera. Zool. Anz. 2016, 261, 48–55. [Google Scholar] [CrossRef]
- Tull, T.; Henn, F.; Betz, O.; Eggs, B. Structure and function of the stylets of hematophagous Triatominae (Hemiptera: Reduviidae), with special reference to Dipetalogaster maxima. Arthropod. Struct. Dev. 2020, 58, 100952. [Google Scholar] [CrossRef]
- Taszakowski, A.; Kaszyca-Taszakowska, N. Teratological cases of the antennae in the family Aradidae (Hemiptera: Heteroptera). Sci. Rep. 2020, 10, 1027. [Google Scholar] [CrossRef] [Green Version]
- Rakitov, R. The mouthparts of the Aradidae (Insecta: Hemiptera: Heteroptera). Arthropod. Struct. Dev. 2022, 71, 101211. [Google Scholar] [CrossRef]
- Rider, D.A. The true bugs of the world (Hemiptera: Heteroptera). Classification and natural history. Ann. Entomol. Soc. Am. 1996, 89, 609–610. [Google Scholar] [CrossRef]
- Contreras, E.; Coscarón, M.C. The Aradidae (Insecta, Hemiptera, Heteroptera) of Argentina. Zootaxa 2012, 3500, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Usinger, R.L. Studies in the american Aradidae with descriptions of new species. Ann. Entomol. Soc. Amer. 1936, 29, 490–516. [Google Scholar] [CrossRef]
- Nardi, J.B.; Bee, C.M.; Miller, L.A.; Taylor, S.J. Distinctive features of the alimentary canal of a fungus-feeding hemipteran, Mezira granulata (Heteroptera: Aradidae). Arthropod. Struct. Dev. 2009, 38, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.S.; Huang, C.W.; Feng, B.C.; Zhou, J.M.; Zhang, C.S.; Jian, H.C. Observation of the bionomics of Mezira membranacea (Fabr.). Plant. Prot. Ect. Ion. 1998, 24, 7–9. [Google Scholar]
- Liguo, C.; Changju, Y.; Keqin, W.; Jianfang, J.; Jie, F. Preliminary studies on injury and living habit of Mezira (Zemira) poriaicola Liu. J. Huazhong Agric. Univ. 2001, 20, 239–240. [Google Scholar] [CrossRef]
- Song, F.; Li, H.; Shao, R.; Shi, A.; Bai, X.; Zheng, X.; Heiss, E.; Cai, W. Rearrangement of mitochondrial tRNA genes in flat bugs (Hemiptera: Aradidae). Sci. Rep. 2016, 6, 25725. [Google Scholar] [CrossRef] [Green Version]
- Altner, H.; Loftus, R. Ultrastructure and function of insect thermo- and hygroreceptors. Annu. Rev. Entomol. 1985, 30, 273–295. [Google Scholar] [CrossRef]
- Taszakowski, A.; Maslowski, A.; Brozek, J. Labial sensory organs of two Leptoglossus species (Hemiptera: Coreidae): Their morphology and supposed function. Insects 2022, 14, 30. [Google Scholar] [CrossRef]
- Spaethe, J.; Brockmann, A.; Halbig, C.; Tautz, J. Size determines antennal sensitivity and behavioral threshold to odors in bumblebee workers. Naturwissenschaften 2007, 94, 733–739. [Google Scholar] [CrossRef]
- Elgara, M.A.; Zhang, D.; Wang, Q.; Wittwer, B.; Pham, H.T.; Johnsona, T.L.; Freelance, C.B.; Coquilleau, M. Insect antennal morphology: The evolution of diverse solutions to odorant perception. Yale J. Biol. Med. 2018, 91, 457–469. [Google Scholar]
- Symonds, M.R.; Elgar, M.A. The evolution of body size, antennal size and host use in parasitoid wasps (Hymenoptera: Chalcidoidea): A phylogenetic comparative analysis. PLoS ONE 2013, 8, e78297. [Google Scholar] [CrossRef] [PubMed]
- María Fernanda López, M.F.; Armendáriz-Toledano, F.; Sámano, J.E.M.; Shibayama-Salas, M.; Zúñiga Sámano, G. Comparative study of the antennae of Dendroctonus rhizophagus and Dendroctonus valens (Curculionidae: Scolytinae): Sensilla types, distribution and club shape. Ann. Entomol. Soc. Am. 2014, 107, 1130–1143. [Google Scholar] [CrossRef]
- Rani, P.U.; Madhavendra, S.S. External morphology of antennal and rostral sensillae in four hemipteran insects and their possible role in host plant selection. Int. J. Trop. Insect. Sci. 2007, 25, 198–207. [Google Scholar] [CrossRef]
- Wang, Y.; Brożek, J.; Dai, W. Functional morphology and sexual dimorphism of antennae of the pear lace bug Stephanitis nashi (Hemiptera: Tingidae). Zool. Anz. 2020, 286, 11–19. [Google Scholar] [CrossRef]
- Spooner, C.S. The phylogeny of the Hemiptera based on a study of the head capsule. Illinois Biol. Monogr. 1938, 16, 1–102. [Google Scholar]
- Baker, G.T.; Chen, X.; Ma, P.W.K. Labial tip sensilla of Blissus leucopterus leucopterus (Hemiptera: Blissidae): Ultrastructure and behavior. Insect Sci. 2008, 15, 271–275. [Google Scholar] [CrossRef]
- Wang, Y.; Brozek, J.; Dai, W. Comparative morphology of the mouthparts in three predatory stink bugs (Heteroptera: Asopinae) reveals feeding specialization of stylets and sensilla. Insects 2020, 11, 762. [Google Scholar] [CrossRef] [PubMed]
- Weirauch, C.; Schuh, R.T. Systematics and evolution of Heteroptera: 25 years of progress. Annu. Rev. Entomol. 2011, 56, 487–510. [Google Scholar] [CrossRef]
- Sinitsina, E.E.; Chaika, S.Y. The antennal sensory apparatus of terrestrial bugs (Heteroptera). Zool. Zhurnal. 1998, 77, 1273–1284. [Google Scholar]
- Chapman, R.F.; Thomas, J.G. The numbers and distribution of sensilla on the mouthparts of Acridoidea. Acrida 1978, 7, 115–148. [Google Scholar]
- Chapman, R.F.; Fraser, J. The chemosensory system of the monophagus grasshopper, Bootettix argentatus Bruner (Orthoptera, Acrididae). Int. J. Insect Morphol. Embryol. 1989, 18, 111–118. [Google Scholar] [CrossRef]









| Sex | Position | Length (µm) | N |
|---|---|---|---|
| Female | Sc | 404.01 ± 13.13 | 3 |
| Pe | 492.61 ± 38.94 | 3 | |
| Fl I | 583.53 ± 26.03 | 3 | |
| Fl II | 528.05 ± 37.01 | 3 | |
| Sf | 13,174.05 ± 605.21 | 3 | |
| Bo | 7104.79 ± 516.78 | 3 | |
| Male | Sc | 414.66 ± 2.53 | 3 |
| Pe | 488.44 ± 15.27 | 3 | |
| Fl I | 587.59 ± 5.53 | 3 | |
| Fl II | 529.39 ± 3.76 | 3 | |
| Sf | 12,371.34 ± 272.23 | 3 | |
| Bo | 6916.45 ± 169.14 | 3 |
| Type | Distribution | Length (µm) | Basal Diameter (µm) | N |
|---|---|---|---|---|
| AnTr I | Fl II | 51.71 ± 4.73 | 3.75 ± 0.29 | 8 |
| AnTr II | Fl II | 40.74 ± 3.90 | 2.59 ± 0.22 | 20 |
| AnTr III | Fl II, Fl I, Pe | 47.45 ± 2.89 | 4.12 ± 0.31 | 20 |
| AnBa I | Fl II | 15.51 ± 1.43 | 2.38 ± 0.17 | 8 |
| AnBa II | Fl II | 8.22 ± 0.15 | 1.7 ± 0.08 | 4 |
| AnBa III | Sc | 12.73 ± 1.04 | 2.65 ± 0.31 | 2 |
| AnCh I | Fl II | 74.95 ± 0.42 | 4.57 ± 0.42 | 8 |
| AnCh II | Fl I, Sc, Pd | 12.07 ± 0.43 | 1.74 ± 0.10 | 5 |
| AnCa | Sc | / | / | 2 |
| AnSt | all antennal segments | 56.26 ± 4.75 | 3.88 ± 0.59 | 20 |
| LaTr I | Lb1 | 34.91 ± 4.65 | 1.73 ± 0.17 | 4 |
| LaTr II | all labial segments | 16.93 ± 3.72 | 1.77 ± 0.28 | 20 |
| LaTr III | Lb3 | 71.96 ± 4.93 | 3.54 ± 0.33 | 10 |
| LaBa I | Lb2 | 10.14 ± 1.30 | 2.05 ± 0.44 | 8 |
| LaBa II | Lb2, Lb3 | 5.13 ± 0.45 | 0.80 ± 0.062 | 4 |
| LaBa III | Lb3 | 11.35 ± 0.31 | 1.31 ± 0.082 | 8 |
| LaCa | Lb3 | / | / | / |
| Species | Food Notes | Number of Labial Segments | Shape of Last Labial Segment | Types of Sensilla of Labial Tip | Distal Mandibular Stylet; Serration | References |
|---|---|---|---|---|---|---|
| Aradus betulae (Linnaeus) | bracket fungi | 4 | cylindrical | four types of sensilla | slightly expanded with subparallel scales and serrate posterior margins | [20] |
| Aradus sp. | / | / | / | / | more than 20 regular, transverse, parallel | [1] |
| Carventus brachypterus Kormilev | / | 3 | cone-shaped | / | / | [20] |
| Dysodius sp. | / | / | / | / | grooves | [1] |
| Isodermus planus Erichon | / | 4 | apex constricted and cleft | sensilla placoid, sensilla setiform | barely expanded with obliquely longitudinal linear ridges and transverse ridges | [20] |
| Mezira yunnana Hsiao | oyster mushroom | 4 | cone-shaped and constricted distally and cleft | sensilla campaniformia, sensilla basiconica | 8–10 central ridge-like teeth and a row of lateral ridges | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zha, S.; Wang, Z.; Tian, L.; Zhao, Y.; Bai, X.; Chen, Z.; Cai, W.; Li, X.; Li, H. Scanning Electron Microscopy of Antennae and Mouthparts of Mezira yunnana Hsiao (Hemiptera: Aradidae): Specialized Microstructures Reflecting Adaptation to Mycetophagy. Insects 2023, 14, 333. https://doi.org/10.3390/insects14040333
Zha S, Wang Z, Tian L, Zhao Y, Bai X, Chen Z, Cai W, Li X, Li H. Scanning Electron Microscopy of Antennae and Mouthparts of Mezira yunnana Hsiao (Hemiptera: Aradidae): Specialized Microstructures Reflecting Adaptation to Mycetophagy. Insects. 2023; 14(4):333. https://doi.org/10.3390/insects14040333
Chicago/Turabian StyleZha, Shiyu, Zhiyao Wang, Li Tian, Yisheng Zhao, Xiaoshuan Bai, Zhaoyang Chen, Wanzhi Cai, Xinyu Li, and Hu Li. 2023. "Scanning Electron Microscopy of Antennae and Mouthparts of Mezira yunnana Hsiao (Hemiptera: Aradidae): Specialized Microstructures Reflecting Adaptation to Mycetophagy" Insects 14, no. 4: 333. https://doi.org/10.3390/insects14040333