
Characterization of Triatoma infestans (Klug, 1834) (Hemiptera: Heteroptera, Reduviidae) from Residual Foci in the States of Bahia and Rio Grande do Sul, Brazil, Using Geometric Morphology
 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Method
2.1. Specimens and Identification
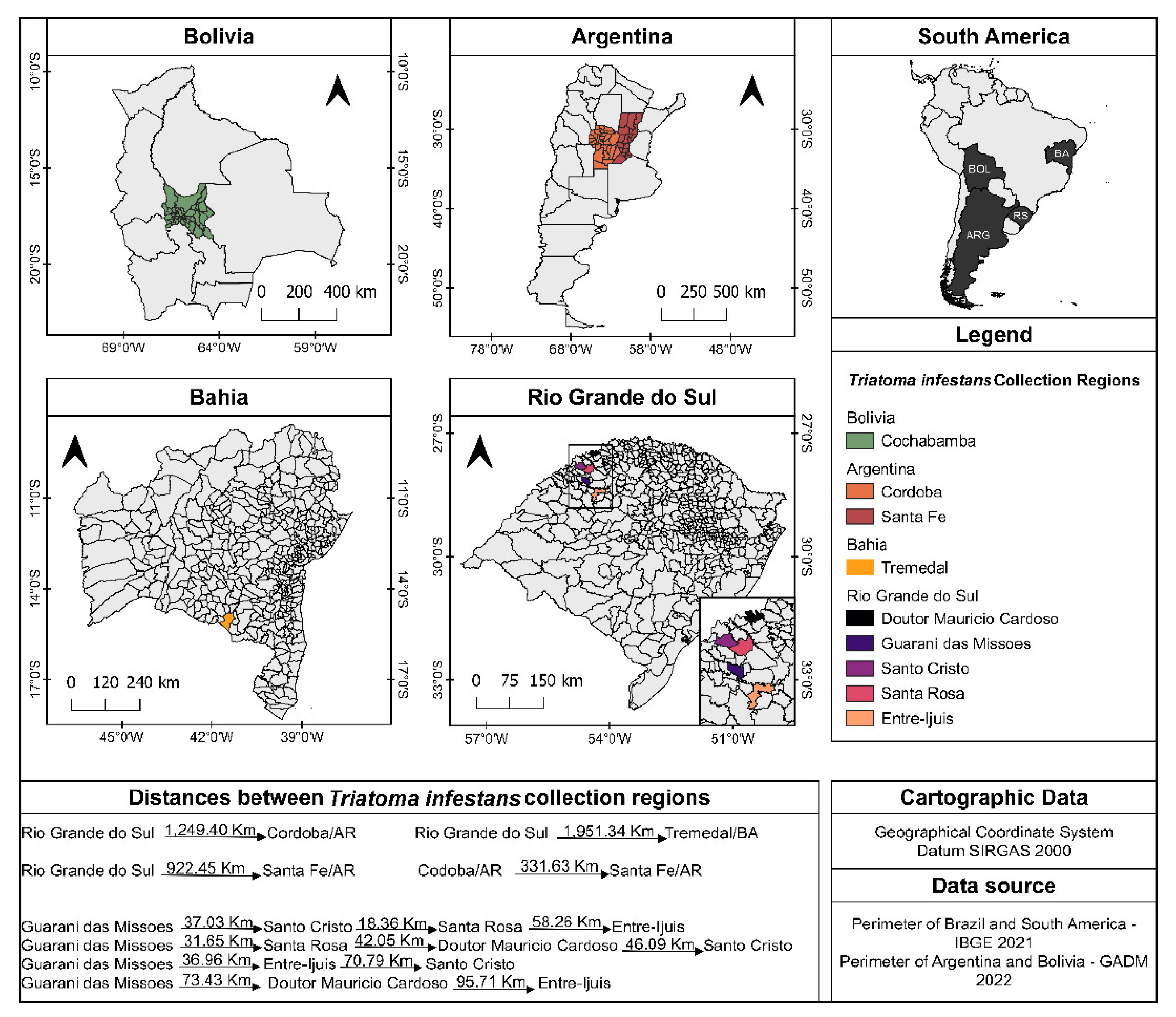
2.2. Sampling
2.3. Obtention of Images and Landmarks
2.4. Size and Shape Variables
2.5. Evaluation of Variations between Groups
3. Results
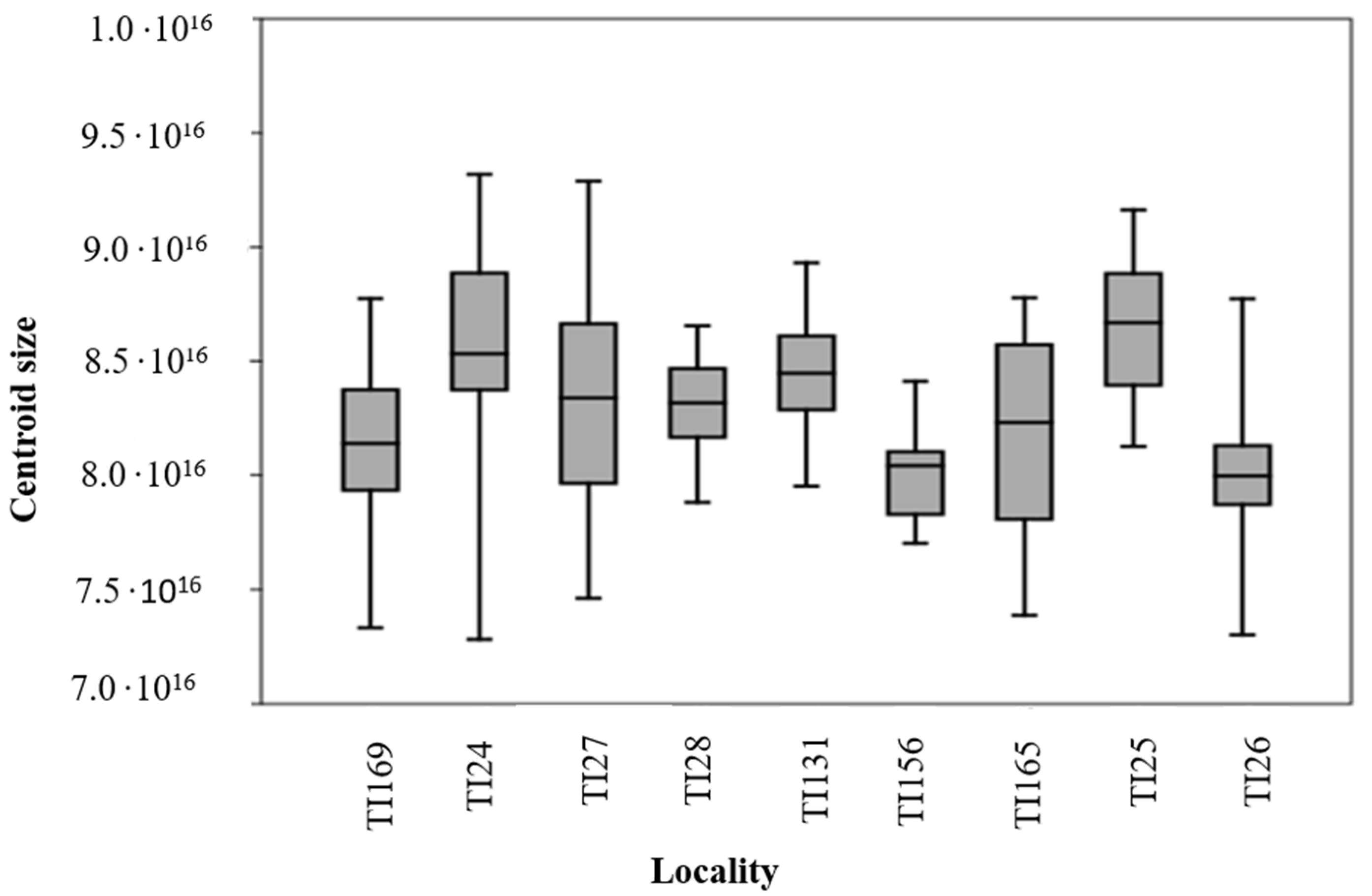
3.1. Analysis of Size and Shape Variables between Populations of Nine Localities
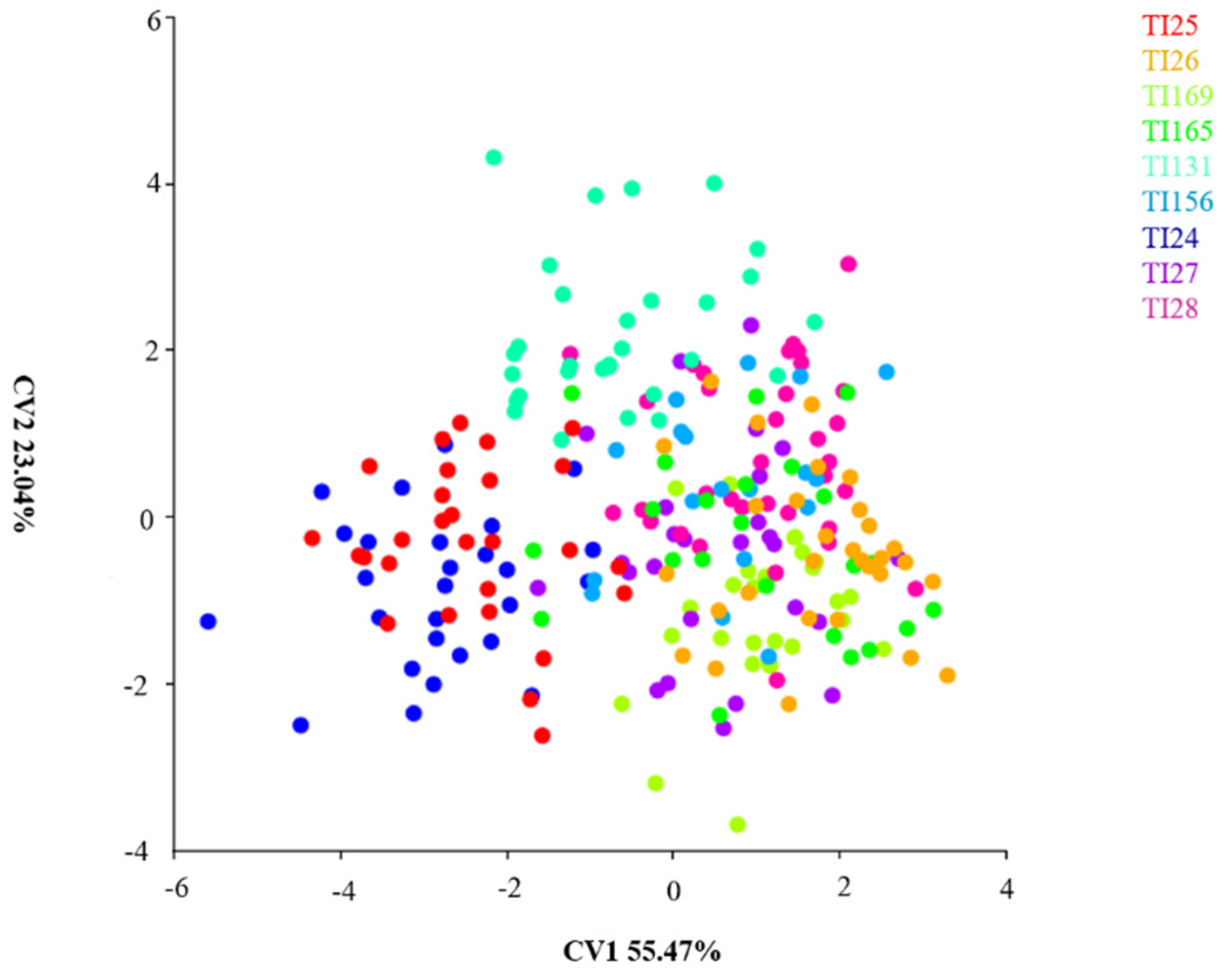
3.2. Principal Component Analysis between Populations of Nine Localities
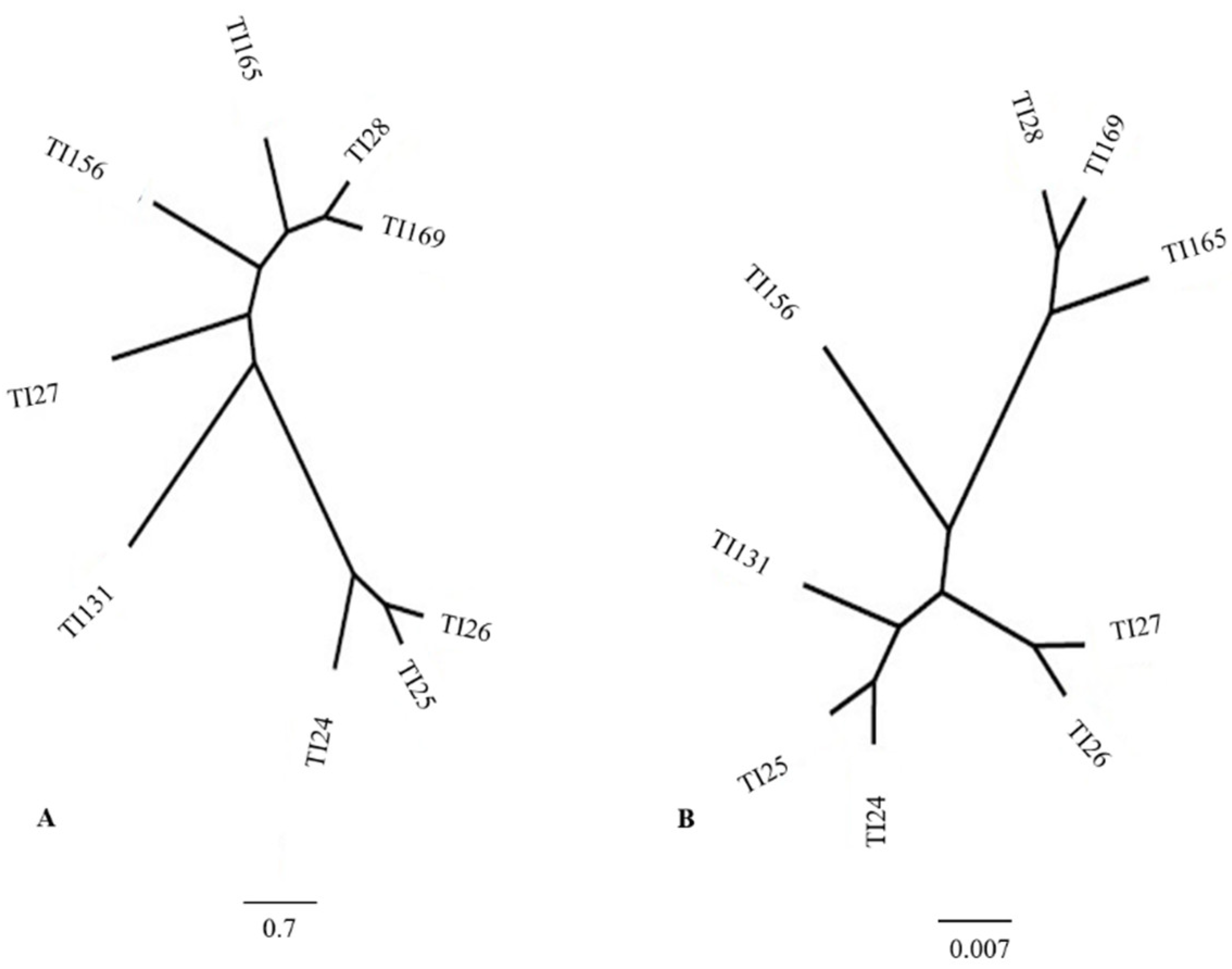
3.3. Canonical Variance Analysis between Populations of 9 Localities
4. Analysis of Size and Shape Variables between Populations of Different Regions
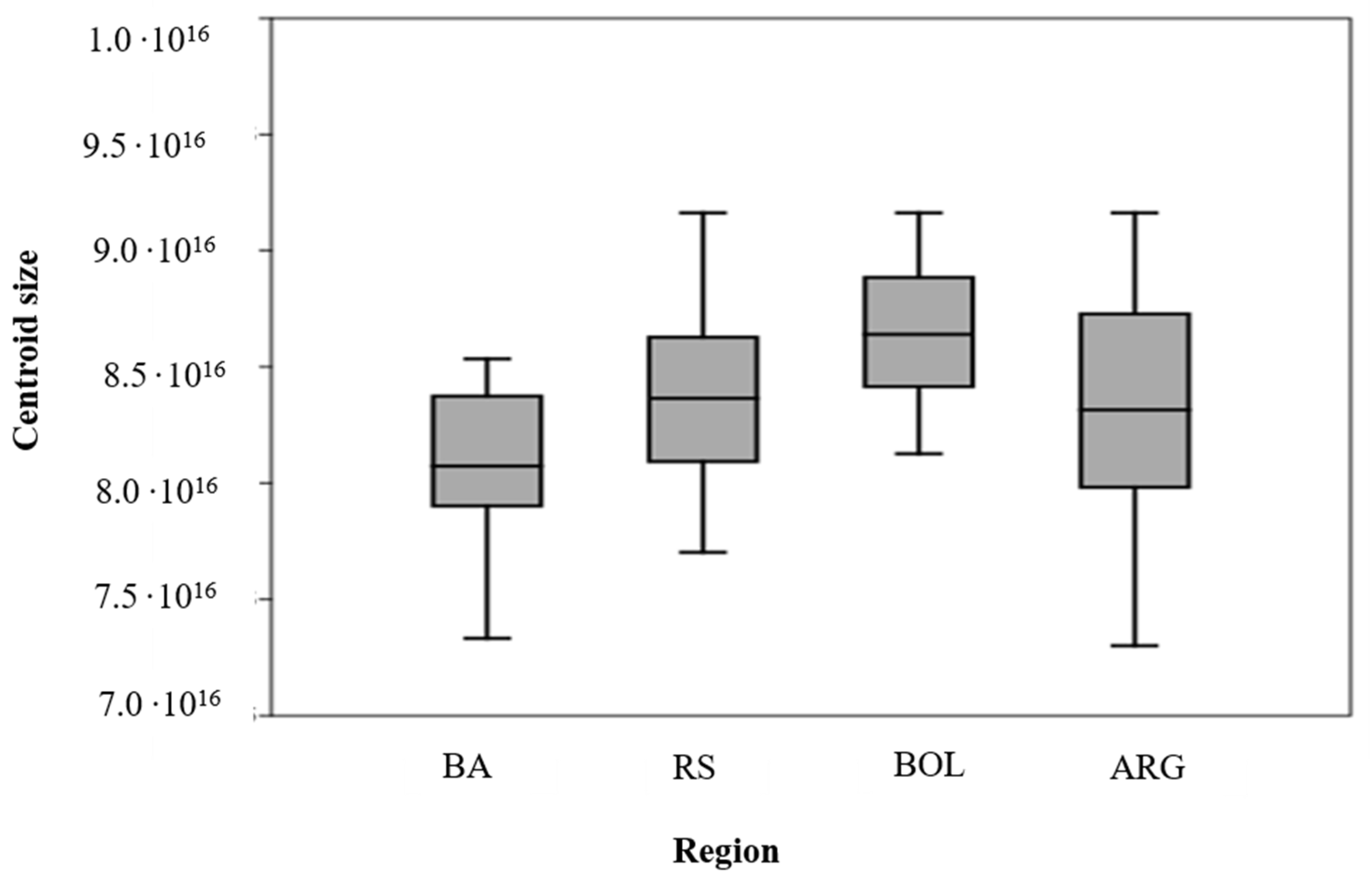
4.1. Centroid Size between Populations of Different Regions
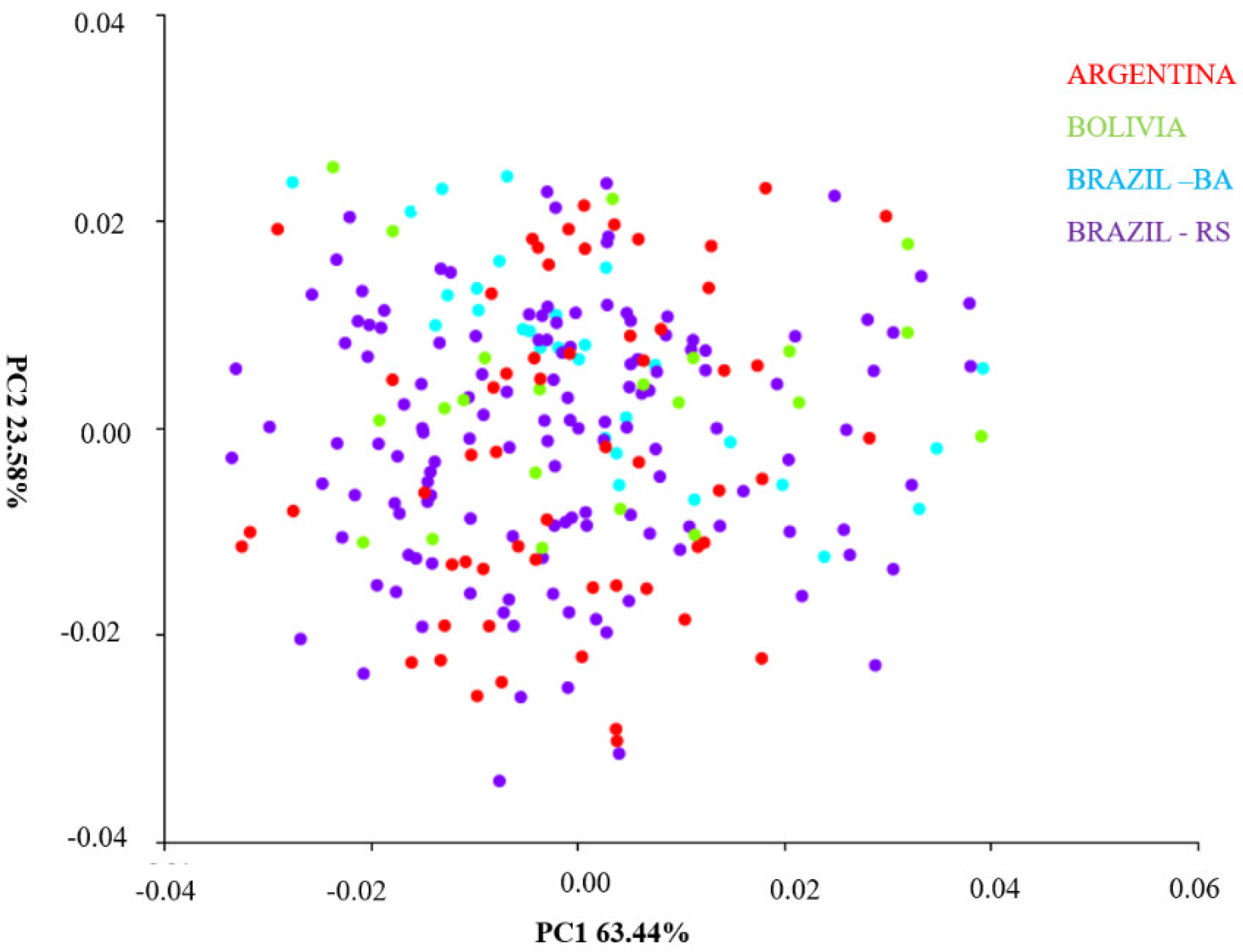
4.2. Principal Component Analysis between Populations of Different Regions
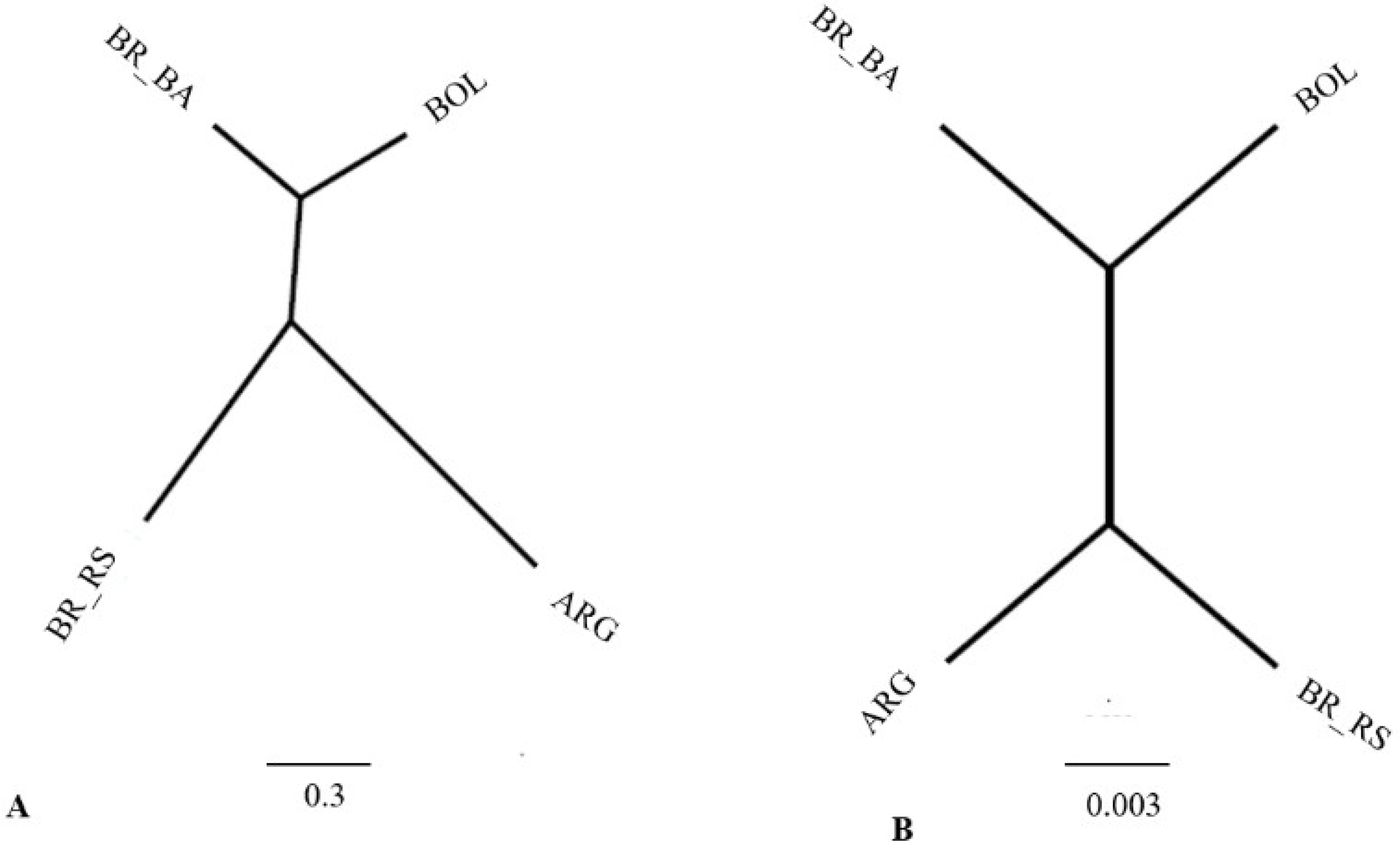
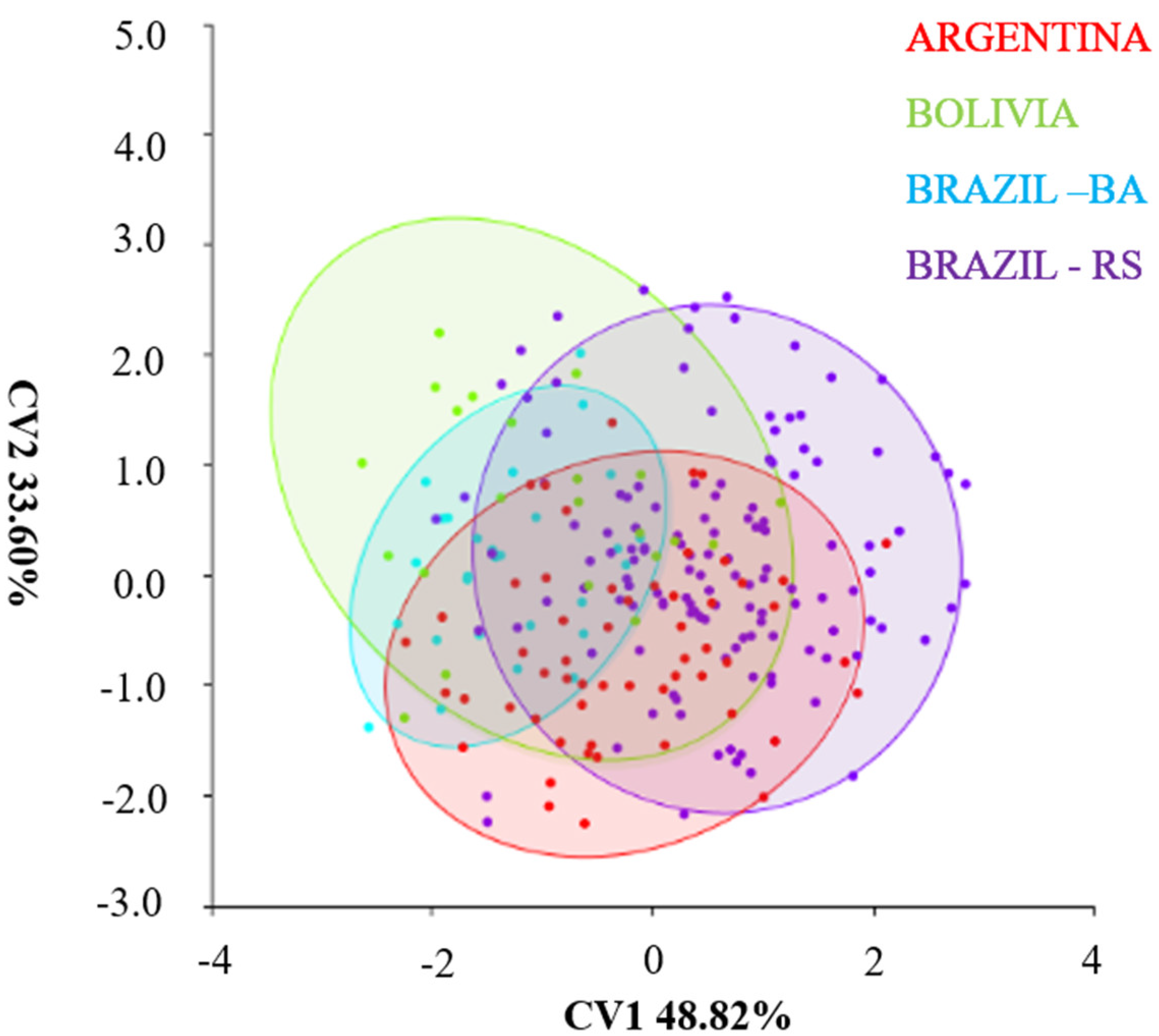
4.3. Canonical Variance Analysis between Populations of Different Regions
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization (PAHO/WHO). Chagas Disease (American Trypanosomiasis). 2020 Disponível. Available online: https://www.who.int/health-topics/chagas-disease#tab=tab_1 (accessed on 4 May 2022).
- Usinger, R.L.; Wygodzinsky, P.; Ryckman, R.E. The biosystematics of triatominae. Annu. Rev. Entomol. 1966, 11, 309–330. [Google Scholar] [CrossRef]
- Aragão, M.B.; Dias, E. Aspectos climáticos da doença de Chagas. I. Considerações sôbre a distribuição geográfica do Triatoma infestans. Rev. Bras. Malar. D. Trop. 1956, 8, 633–644. [Google Scholar]
- Aragão, M.B. Sôbre a dispersão de Triatoma infestans. Rev. Soc. Bras. Med. Trop. 1971, 5, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Forattini, O.P. Biogeografia, origem e distribuição da domiciliação de triatomíneos no Brasil. Rev. Saúde Pública 2006, 40, 964–998. [Google Scholar] [CrossRef] [Green Version]
- Schofield, C.J. Biosystematics and evolution of the Triatominae. Cad. Saúde Pública 2000, 16 (Suppl. 2), 89–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dujardin, J.P.; Chavez, T.; Moreno, M.; Machane, M.; Noireau, F.; Schofield, C.J. Comparison of isoenzyme electrophoresis and morphometric analysis for phylogenetic reconstruction of the Rhodniini (Hemiptera: Reduviidae: Triatominae). J. Med. Entomol. 1999, 36, 653–659. [Google Scholar] [CrossRef]
- Noireau, F. Wild Triatoma infestans, a potential threat that needs to be monitored. Mem. Inst. Oswaldo Cruz 2009, 104, 60–64. [Google Scholar] [CrossRef] [Green Version]
- Waleckx, E.; Salas, R.; Huamán, N.; Buitrago, R.; Bosseno, M.F.; Aliaga, C.; Barnabé, C.; Rodriguez, R.; Zoveda, F.; Monje, M.; et al. New insights on the Chagas disease main vector Triatoma infestans (Reduviidae, Triatominae) brought by the genetic analysis of Bolivian sylvatic populations. Infect. Genet. Evol. 2011, 11, 1045–1057. [Google Scholar] [CrossRef] [PubMed]
- Piccinali, R.V.; Marcet, P.L.; Noireau, F.; Kitron, U.; Gürtler, R.E.; Dotson, E.M. Molecular Population Genetics and Phylogeography of the Chagas Disease Vector Triatoma infestans in South America. J. Med. Entomol. 2009, 46, 796–809. [Google Scholar] [CrossRef] [Green Version]
- Piccinali, R.V.; Marcet, P.L.; Ceballos, L.A.; Kitron, U.; Gürtler, R.E.; Dotson, E.M. Genetic variability, phylogenetic relationships and gene flow in Triatoma infestans dark morphs from the Argentinean Chaco. Infect. Genet. Evol. 2011, 11, 895–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, J.C. Southern Cone Initiative for the elimination of domestic populations of Triatoma infestans and the interruption of transfusional Chagas disease. Historical aspects, present situation, and perspectives. Mem. Inst. Oswaldo Cruz 2007, 102 (Suppl. S1), 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silveira, A.C.; Dias, J.C. O controle da transmissão vetorial. Rev. Soc. Bras. Med. Trop. 2011, 44 (Suppl. S2), 52–63. [Google Scholar] [CrossRef] [PubMed]
- Bedin, C.; Wilhelm, T.; Villela, M.M.; da Silva, G.C.C.; Riffel, A.P.K.; Sackis, P.; de Mello, F. Residual foci of Triatoma infestans infestation: Surveillance and control in Rio Grande do Sul, Brazil, 2001–2018. Rev. Soc. Bras. Med. Trop. 2021, 54, e0530-2020:2021. [Google Scholar] [CrossRef]
- Lent, H.; Wygodzinsky, P. Revision of the Triatominae (Hemiptera: Reduviidae) and their significance as vectors of Chagas disease. Bull. Am. Mus. Nat. Hist. 1979, 163, 123–520. Available online: http://hdl.handle.net/2246/1282 (accessed on 6 March 2022).
- Oliveira, J.; Marcet, P.L.; Takiya, D.M.; Mendonça, V.J.; Belintani, T.; Bargues, M.D.; Mateo, L.; Chagas, V.; Folly-Ramos, E.; Cordeiro-Estrela, P.; et al. Combined phylogenetic and morphometric information to delimit and unify the Triatoma brasiliensis species complex and the Brasiliensis subcomplex. Acta Trop. 2017, 170, 140–148. [Google Scholar] [CrossRef]
- Monteiro, L.R.; Reis, S.F. Princípios de Morfometria Geométrica; Holos: Ribeirão Preto, Brazil, 1999; p. 189. Available online: https://www.holoseditora.com.br/produto/principios-de-morfometria-geometrica (accessed on 12 September 2022).
- Belintani, T.; de Paiva, V.F.; de Oliveira, J.; da Rosa, J.A. New in morphometry: Geometric morphometry of the external female genitalia of Triatominae (Hemiptera: Reduviidae). Acta Trop. 2022, 229, 106383. [Google Scholar] [CrossRef]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Bookstein, F.L. (Ed.) Morphometric Tools for Landmark Data: Geometry and Biology; Statistics in Medicine; Cambridge University Press: Cambridge, UK, 1991; Volume 12, pp. 714–715. Available online: https://www.abebooks.co.uk/9780521585989/Morphometric-Tools-Landmark-Data-Geometry-0521585988/plp?cm_sp=plped-_-1-_-isbn (accessed on 3 November 2021).
- Klingenberg, C.P.; Leamy, L.J.; Routman, E.J.; Cheverud, J.M. Genetic Architecture of Mandible Shape in Mice: Effects of Quantitative Trait Loci Analyzed by Geometric Morphometrics. Genetics 2001, 157, 785–802. [Google Scholar] [CrossRef]
- Belintani, T.; Oliveira, J.; Pinotti, H.; Silva, L.A.; Alevi, K.C.C.; Galvao, C.; da Rosa, J.A. Phylogenetic and phenotypic relationships of the Triatoma sordida subcomplex (Hemiptera: Reduviidae: Triatominae). Acta Trop. 2020, 212, 105679. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 24 October 2022).
- Klingenberg, C.P.; Monteiro, L.R. Distances and directions in multidimensional shape spaces: Implications for morphometric applications. Syst. Biol. 2005, 54, 678–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, J.C.P.; Schofield, C.J. The evolution of Chagas disease (American Trypanosomiasis) control after 90 years since Carlos Chagas discovery. Mem. Inst. Oswaldo Cruz 1999, 94, 103–121. [Google Scholar] [CrossRef] [Green Version]
- Araújo, R.F.; Mendonça, V.J.; Da Rosa, J.A.; De Matos, J.F.M.; Lima, S.C.R.; Figueiredo, M.A.D.A. Description of a newly discovered Triatoma infestans (Hemiptera: Reduviidae) Foci in Ibipeba, State of Bahia, Brazil. Rev. Soc. Bras. Med. Trop. 2014, 47, 513–516. [Google Scholar] [CrossRef] [Green Version]
- Gurgel-Gonçalves, R.; Ferreira, J.B.C.; Rosa, A.F.; Bar, M.E.; Galvão, C. Geometric morphometrics and ecological niche modelling for delimitation of near-sibling triatomine species. Med. Vet. Entomol. 2011, 25, 84–93. [Google Scholar] [CrossRef]
- Jaramillo, O.N.; Castillo, D.; Wolff, E.M. Geometric morphometric differences between Panstrongylus geniculatus from field and laboratory. Mem. Do Inst. Oswaldo Cruz. 2002, 97, 667–673. [Google Scholar] [CrossRef]
- Kamimura, E.H.; Viana, M.C.; Lilioso, M.; Fontes, F.H.M.; Pires-Silva, D.; Valença-Barbosa, C.; Carbajal-De-La-Fuente, A.L.; Folly-Ramos, E.; Solferin, V.N.; Thyssen, P.J.; et al. Drivers of molecular and morphometric variation in Triatoma brasiliensis (Hemiptera: Triatominae): The resolution of geometric morphometrics for populational structuring on a microgeographical scale. Parasites Vectors 2020, 13, 455. [Google Scholar] [CrossRef] [PubMed]
- Stevens, L.; Dorn, P.L. Population genetics of Triatominae. In American Trypanosomiasis Chagas Disease: One Hundred Years of Research, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 169–196. [Google Scholar] [CrossRef]
- Sobre los registros aislados de Triatoma infestans (Klug, 1834) (Heteroptera: Reduviidae: Triatominae) en el sur de Chile. Arq. Entomolóxicos 2016, 15, 121–124. Available online: https://dialnet.unirioja.es/servlet/articulo?codigo=6500415 (accessed on 2 December 2022).
- SECEM (Sistema de Estadísticas de Comercio Exterior del MERCOSUR). Available online: https://estadisticas.mercosur.int/ (accessed on 26 November 2022).
- Campos-Soto, R.; Díaz-Campusano, G.; Rives-Blanchard, N.; Cianferoni, F.; Torres-Pérez, F. Biogeographic origin and phylogenetic relationships of Mepraia (Hemiptera, Reduviidae) on islands of northern Chile. PLoS ONE 2020, 15, e0234056. [Google Scholar] [CrossRef]
- Dias, J.C.P. Evolution of Chagas Disease Screening Programs and Control Programs Historical Perspective. In Global Heart; Elsevier: Amsterdam, The Netherlands, 2015; Volume 10, pp. 193–202. [Google Scholar] [CrossRef]
- Rocklöv, J.; Dubrow, R. Author Correction: Climate change: An enduring challenge for vector-borne disease prevention and control. Nat. Immunol. 2020, 21, 695. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.; Dornak, L.L.; Almeida, C.E.; Peterson, A.T. Distributional potential of the Triatoma brasiliensis species complex at present and under scenarios of future climate conditions. Parasites Vectors 2014, 7, 1–10. [Google Scholar] [CrossRef]
- Dujardin, J.P.; Panzera, P.; Schofield, C.J. Triatominae as a model of morphological plasticity under ecological pressure. Mem. Inst. Oswaldo Cruz. 1999, 94, 223–228. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | ID | Locality | Initiate | Generation * |
|---|---|---|---|---|
| T. infestans | TI24 | Guarani das Missões/RS */Brazil | 2006 | F22 |
| T. infestans | TI25 | Cordoba/Argentina | 1996 | F42 |
| T. infestans | TI26 | Santa Fé/Argentina | 1994 | F46 |
| T. infestans | TI27 | Entre Ijuís/RS/Brazil | 1996 | F42 |
| T. infestans | TI28 | Santo Cristo/RS/Brazil | 1996 | F42 |
| T. infestans | TI131 | Santa Rosa/RS/Brazil | 2008 | F18 |
| T. infestans | TI156 *** | Santa Rosa/RS/Brazil | 2014 | F6 |
| T. infestans | TI165 | Cochabamba/Bolivia | 2015 | F4 |
| T. infestans | TI169 *** | Tremedal/BA **/Brazil | 2010 | F14 |
| Samples | TI25 | TI26 | TI169 | TI165 | TI131 | TI156 | TI24 | TI27 | TI28 |
|---|---|---|---|---|---|---|---|---|---|
| TI25 | 0.00 | ||||||||
| TI26 | 1.3641 | 0.00 | |||||||
| TI169 | 3.7685 | 1.9871 | 0.00 | ||||||
| TI165 | 3.6017 | 2.2425 | 1.735 | 0.00 | |||||
| TI131 | 3.2075 | 3.7394 | 3.885 | 3.3427 | 0.00 | ||||
| TI156 | 3.4669 | 2.1847 | 2.3269 | 2.0245 | 2.6227 | 0.00 | |||
| TI24 | 1.6277 | 1.6703 | 4.132 | 4.219 | 3.840 | 3.896 | 0.00 | ||
| TI27 | 3.5218 | 2.4463 | 2.472 | 2.2997 | 3.4285 | 1.2399 | 3.736 | 0.00 | |
| TI28 | 3.7424 | 1.9028 | 2.1189 | 1.6677 | 2.3642 | 1.3464 | 4.2309 | 2.0298 | 0.00 |
| Regions | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| 1. ARG | 0.0000 | |||
| 2. BOL | 1.7701 | 0.0000 | ||
| 3. BR_BA | 1.5817 | 1.5082 | 0.0000 | |
| 4. BR_RS | 1.2438 | 1.9315 | 1.9828 | 0.0000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peixoto, S.; Belintani, T.; Rocha, D.; Fontes, F.; Frota, N.; Galvão, C. Characterization of Triatoma infestans (Klug, 1834) (Hemiptera: Heteroptera, Reduviidae) from Residual Foci in the States of Bahia and Rio Grande do Sul, Brazil, Using Geometric Morphology. Insects 2023, 14, 325. https://doi.org/10.3390/insects14040325
Peixoto S, Belintani T, Rocha D, Fontes F, Frota N, Galvão C. Characterization of Triatoma infestans (Klug, 1834) (Hemiptera: Heteroptera, Reduviidae) from Residual Foci in the States of Bahia and Rio Grande do Sul, Brazil, Using Geometric Morphology. Insects. 2023; 14(4):325. https://doi.org/10.3390/insects14040325
Chicago/Turabian StylePeixoto, Solange, Tiago Belintani, Dayse Rocha, Felipe Fontes, Natália Frota, and Cleber Galvão. 2023. "Characterization of Triatoma infestans (Klug, 1834) (Hemiptera: Heteroptera, Reduviidae) from Residual Foci in the States of Bahia and Rio Grande do Sul, Brazil, Using Geometric Morphology" Insects 14, no. 4: 325. https://doi.org/10.3390/insects14040325