
Drones Do Not Drift between Nests in a Wild Population of Apis cerana

{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
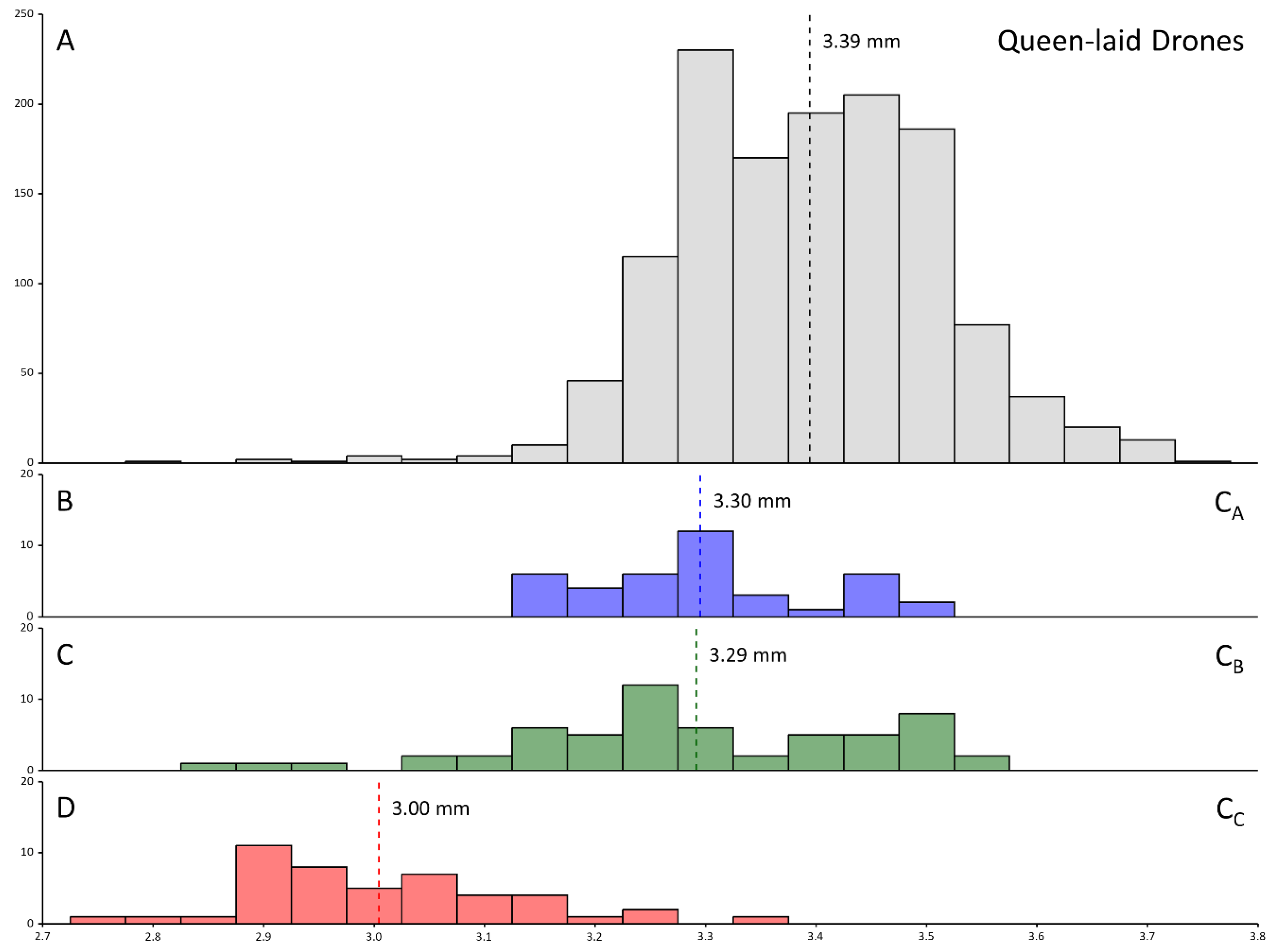
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heimpel, G.E.; de Boer, J.G. Sex determination in the hymenoptera. Annu. Rev. Entomol. 2008, 53, 209–230. [Google Scholar] [CrossRef] [PubMed]
- Woyke, J. Evidence and action of cannibalism substance in Apis cerana indica. J. Apic. Res. 1980, 19, 6–16. [Google Scholar] [CrossRef]
- Cook, J.; Crozier, R. Sex determination and population biology in the Hymenoptera. Trends Ecol. Evol. 1995, 10, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Beye, M.; Hasselmann, M.; Fondrk, M.K.; Page, R.E., Jr.; Omholt, S.W. The gene csd is the primary signal for sexual development in the honeybee and encodes an SR-type protein. Cell 2003, 114, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Harpur, B.A.; Sobhani, M.; Zayed, A. A review of the consequences of complementary sex determination and diploid male production on mating failures in the Hymenoptera. Entomol. Exp. Appl. 2013, 146, 156–164. [Google Scholar] [CrossRef]
- Ayabe, T.; Hoshiba, H.; Ono, M. Cytological evidence for triploid males and females in the bumblebee, Bombus terrestris. Chromosome Res. 2004, 12, 215–223. [Google Scholar] [CrossRef]
- Zayed, A.; Packer, L. Complementary sex determination substantially increases extinction proneness of haplodiploid populations. Proc. Natl. Acad. Sci. USA 2005, 102, 10742–10746. [Google Scholar] [CrossRef] [Green Version]
- Leung, K.; van der Meulen, H. Revisiting the hymenopteran diploid male vortex: A review of avoidance mechanisms and incidence. Entomol. Exp. Appl. 2022, 170, 1010–1031. [Google Scholar] [CrossRef]
- Zayed, A. Bee genetics and conservation. Apidologie 2009, 40, 237–262. [Google Scholar] [CrossRef] [Green Version]
- Hagan, T.; Gloag, R. Founder effects on sex determination systems in invasive social insects. Curr. Opin. Insect Sci. 2021, 46, 31–38. [Google Scholar] [CrossRef]
- Koeniger, N.; Koeniger, G.; Gries, M.; Tingek, S. Drone competition at drone congregation areas in four Apis species. Apidologie 2005, 36, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Zmarlicki, C.; Morse, R. Drone congregation areas. J. Apic. Res. 1963, 2, 64–66. [Google Scholar] [CrossRef]
- Utaipanon, P.; Holmes, M.J.; Chapman, N.C.; Oldroyd, B.P. Estimating the density of honey bee (Apis mellifera) colonies using trapped drones: Area sampled and drone mating flight distance. Apidologie 2019, 50, 578–592. [Google Scholar] [CrossRef]
- Baudry, E.; Solignac, M.; Garnery, L.; Gries, M.; Cornuet, J.; Koeniger, N. Relatedness among honeybees (Apis mellifera) of a drone congregation. Proc. R. Soc. Lond. B Biol. Sci. 1998, 265, 2009–2014. [Google Scholar] [CrossRef] [Green Version]
- Crailsheim, K. Trophallactic interactions in the adult honeybee (Apis mellifera L.). Apidologie 1998, 29, 97–112. [Google Scholar] [CrossRef]
- Hayashi, S.; Sasaki, T.; Farkhary, S.I.; Kaneko, K.; Hosaka, Y.; Satoh, T. Age-related variation of homing range in honeybee males (Apis mellifera). Insectes Sociaux 2022, 69, 37–45. [Google Scholar] [CrossRef]
- Winston, M.L.; Higo, H.A.; Colley, S.J.; Pankiw, T.; Slessor, K.N. The role of queen mandibular pheromone and colony congestion in honey bee (Apis mellifera L.) reproductive swarming (Hymenoptera: Apidae). J. Insect Behav. 1991, 4, 649–660. [Google Scholar] [CrossRef]
- Schmidt, J.O. Dispersal distance and direction of reproductive European honey bee swarms (Hymenoptera: Apidae). J. Kans. Entomol. Soc. 1995, 68, 320–325. [Google Scholar]
- Villa, J.D. Swarming Behavior of Honey Bees (Hymenoptera: Apidae) in Southeastern Louisiana. Ann. Entomol. Soc. Am. 2004, 97, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Robinson, G.E.; Page, R.E.; Fondrk, M.K. Intracolonial behavioral variation in worker oviposition, oophagy, and larval care in queenless honey bee colonies. Behav. Ecol. Sociobiol. 1990, 26, 315–323. [Google Scholar] [CrossRef]
- Page, R.; Erickson, E. Reproduction by worker honey bees (Apis mellifera L.). Behav. Ecol. Sociobiol. 1988, 23, 117–126. [Google Scholar] [CrossRef]
- Oldroyd, B.P.; Halling, L.A.; Good, G.; Wattanachaiyingcharoen, W.; Barron, A.B.; Nanork, P.; Wongsiri, S.; Ratnieks, F.L. Worker policing and worker reproduction in Apis cerana. Behav. Ecol. Sociobiol. 2001, 50, 371–377. [Google Scholar] [CrossRef]
- Pfeiffer, K.J.; Crailsheim, K. Drifting of honeybees. Insectes Sociaux 1998, 45, 151–167. [Google Scholar] [CrossRef]
- Neumann, P.; Moritz, R.F.A.; Mautz, D.R. Colony evaluation is not affected by drifting of drone and worker honeybees (Apis mellifera L.) at a performance testing apiary. Apidologie 2000, 31, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Currie, R.; Jay, S. The influence of a colony’s queen state on the drifting of drone honey bees (Apis mellifera L.). Apidologie 1991, 22, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Currie, R.; Jay, S. Drifting behaviour of drone honey bees (Apis mellifera L.) in commercial apiaries. J. Apic. Res. 1991, 30, 61–68. [Google Scholar] [CrossRef]
- Reyes, M.; Crauser, D.; Prado, A.; Le Conte, Y. Flight activity of honey bee (Apis mellifera) drones. Apidologie 2019, 50, 669–680. [Google Scholar] [CrossRef]
- Fujiwara, S.; Miura, H.; Kumagai, T.; Sawaguchi, T.; Naya, S.; Goto, T.; Asanuma, H.; Suzuki, K. Drone congregation of Apis cerana japonica (Radoszkowski, 1877) above large trees. Apidologie 1994, 25, 331–337. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, S.; Satoh, T. Landscape features causing the local congregation of honeybee males (Apis mellifera L.). Ethology 2021, 127, 582–591. [Google Scholar] [CrossRef]
- Koetz, A.H. Ecology, Behaviour and Control of Apis cerana with a Focus on Relevance to the Australian Incursion. Insects 2013, 4, 558–592. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.; Annand, N.; Lacey, M.; Ete, S. Control of Asian Honeybees in the Solomon Islands; Australian Government, Australian Centre for International Agricultural Research: Bruce, Australia, 2010. [Google Scholar]
- Gloag, R.; Ding, G.; Christie, J.R.; Buchmann, G.; Beekman, M.; Oldroyd, B.P. An invasive social insect overcomes genetic load at the sex locus. Nat. Ecol. Evol. 2017, 1, 11. [Google Scholar] [CrossRef]
- Ding, G.; Hasselmann, M.; Huang, J.; Roberts, J.; Oldroyd, B.P.; Gloag, R. Global allele polymorphism indicates a high rate of allele genesis at a locus under balancing selection. Heredity 2020, 26, 163–177. [Google Scholar] [CrossRef]
- Gloag, R.S.; Christie, J.R.; Ding, G.; Stephens, R.E.; Buchmann, G.; Oldroyd, B.P. Workers’ sons rescue genetic diversity at the sex locus in an invasive honey bee population. Mol. Ecol. 2019, 28, 1585–1592. [Google Scholar] [CrossRef]
- Walsh, S.; Metzger, D.; Higuchi, R. Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques 1991, 10, 506–513. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, J.I.; Shimizu, S.; Koyama, S.; Kimura, K.; Shimizu, K.; Yoshida, T. Variable microsatellite loci isolated from the Asian honeybee, Apis cerana (Hymenoptera; Apidae). Mol. Ecol. Resour. 2009, 9, 819–821. [Google Scholar] [CrossRef]
- Solignac, M.; Vautrin, D.; Loiseau, A.; Mougel, F.; Baudry, E.; Estoup, A.; Garnery, L.; Haberl, M.; Cornuet, J.M. Five hundred and fifty microsatellite markers for the study of the honeybee (Apis mellifera L.) genome. Mol. Ecol. Notes 2003, 3, 307–311. [Google Scholar] [CrossRef]
- Paar, J.; Oldroyd, B.P.; Huettinger, E.; Kastberger, G. Drifting of workers in nest aggregations of the giant honeybee Apis dorsata. Apidologie 2002, 33, 553–561. [Google Scholar] [CrossRef] [Green Version]
- Chapman, N.C.; Nanork, P.; Reddy, M.S.; Bhat, N.S.; Beekman, M.; Oldroyd, B.P. Nestmate recognition by guards of the Asian hive bee Apis cerana. Insectes Sociaux 2008, 55, 382–386. [Google Scholar] [CrossRef]
- Barron, A.B.; Oldroyd, B.P.; Ratnieks, F.L. Worker reproduction in honey-bees (Apis) and the anarchic syndrome: A review. Behav. Ecol. Sociobiol. 2001, 50, 199–208. [Google Scholar] [CrossRef]
- Chapman, N.C.; Nanork, P.; Gloag, R.S.; Wattanachaiyingcharoen, W.; Beekman, M.; Oldroyd, B.P. Queenless colonies of the Asian red dwarf honey bee (Apis florea) are infiltrated by workers from other queenless colonies. Behav. Ecol. 2009, 20, 817–820. [Google Scholar] [CrossRef] [Green Version]
- Miller Iii, D.; Ratnieks, F. The timing of worker reproduction and breakdown of policing behaviour in queenless honey bee (Apis mellifera L.) societies. Insectes Sociaux 2001, 48, 178–184. [Google Scholar] [CrossRef]
- Utaipanon, P.; Holmes, M.J.; Oldroyd, B.P.M. Queenless colonies contribute to the male breeding population at honey bee drone congregation areas. Insectes Sociaux 2019, 66, 593–599. [Google Scholar] [CrossRef]
- Holmes, M.J.; Tan, K.; Wang, Z.; Oldroyd, B.P.; Beekman, M. Why acquiesce? Worker reproductive parasitism in the Eastern honeybee (Apis cerana). J. Evol. Biol. 2014, 27, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Currie, R.W.; Jay, S.C. The Influence of a Colony’s Queen State, Time of Year, and Drifting Behaviour, on the Acceptance and Longevity of Adult Drone Honeybees (Apis mellifera L.). J. Apic. Res. 1988, 27, 219–226. [Google Scholar] [CrossRef]
- Downs, S.G.; Ratnieks, F.L. Adaptive shifts in honey bee (Apis mellifera L.) guarding behavior support predictions of the acceptance threshold model. Behav. Ecol. 2000, 11, 326–333. [Google Scholar] [CrossRef] [Green Version]
- Chapman, N.C.; Beekman, M.; Oldroyd, B.P. Worker reproductive parasitism and drift in the western honeybee Apis mellifera. Behav. Ecol. Sociobiol. 2010, 64, 419–427. [Google Scholar] [CrossRef]
- Couvillon, M.J.; Roy, G.G.F.; Ratnieks, F.L.W. Recognition errors by honey bee (Apis mellifera) guards demonstrate overlapping cues in conspecific recognition. J. Apic. Res. 2009, 48, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Pradella, D.; Martin, S.J.; Dani, F.R. Using Errors by Guard Honeybees (Apis mellifera) to Gain New Insights into Nestmate Recognition Signals. Chem. Senses 2015, 40, 649–653. [Google Scholar] [CrossRef] [Green Version]
- Nanork, P.; Chapman, N.C.; Wongsiri, S.; Lim, J.; Gloag, R.S.; Oldroyd, B.P. Social parasitism by workers in queenless and queenright Apis cerana colonies. Mol. Ecol. 2007, 16, 1107–1114. [Google Scholar] [CrossRef]
- Beekman, M.; Oldroyd, B.P. When Workers Disunite: Intraspecific Parasitism by Eusocial Bees. Annu. Rev. Entomol. 2008, 53, 19–37. [Google Scholar] [CrossRef]
- Strauss, K.; Scharpenberg, H.; Crewe, R.M.; Glahn, F.; Foth, H.; Moritz, R.F.A. The role of the queen mandibular gland pheromone in honeybees (Apis mellifera): Honest signal or suppressive agent? Behav. Ecol. Sociobiol. 2008, 62, 1523–1531. [Google Scholar] [CrossRef] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hagan, T.; Lim, J.; Gloag, R. Drones Do Not Drift between Nests in a Wild Population of Apis cerana. Insects 2023, 14, 323. https://doi.org/10.3390/insects14040323
Hagan T, Lim J, Gloag R. Drones Do Not Drift between Nests in a Wild Population of Apis cerana. Insects. 2023; 14(4):323. https://doi.org/10.3390/insects14040323
Chicago/Turabian StyleHagan, Thomas, Julianne Lim, and Rosalyn Gloag. 2023. "Drones Do Not Drift between Nests in a Wild Population of Apis cerana" Insects 14, no. 4: 323. https://doi.org/10.3390/insects14040323