


Adaptive Reproductive Strategies of an Ectoparasitoid Sclerodermus guani under the Stress of Its Entomopathogenic Fungus Beauveria bassiana

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. B. bassiana Suspensions
2.3. Interactions between S. guani and B. bassiana
2.3.1. The Pathogenicity of B. bassiana to S. guani
2.3.2. The Pathogenicity of B. bassiana Infects the Immature Offspring
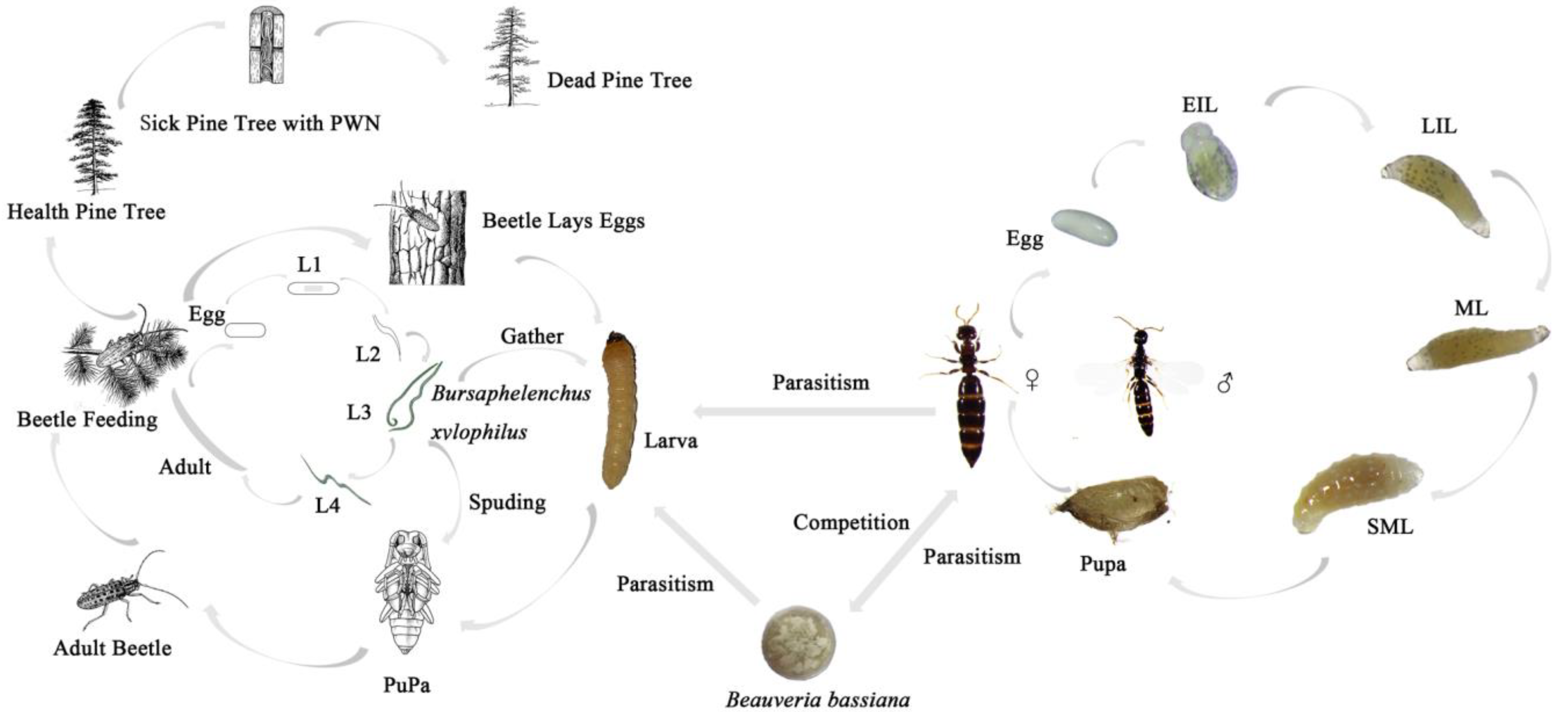
2.4. Interactions between S. guani, B. bassiana, and M. alternatus
2.4.1. The Pathogenicity of B. bassiana to M. alternatus
2.4.2. The Model
2.4.3. The Mortality Effect of S. guani on the Host under B. bassiana Stress
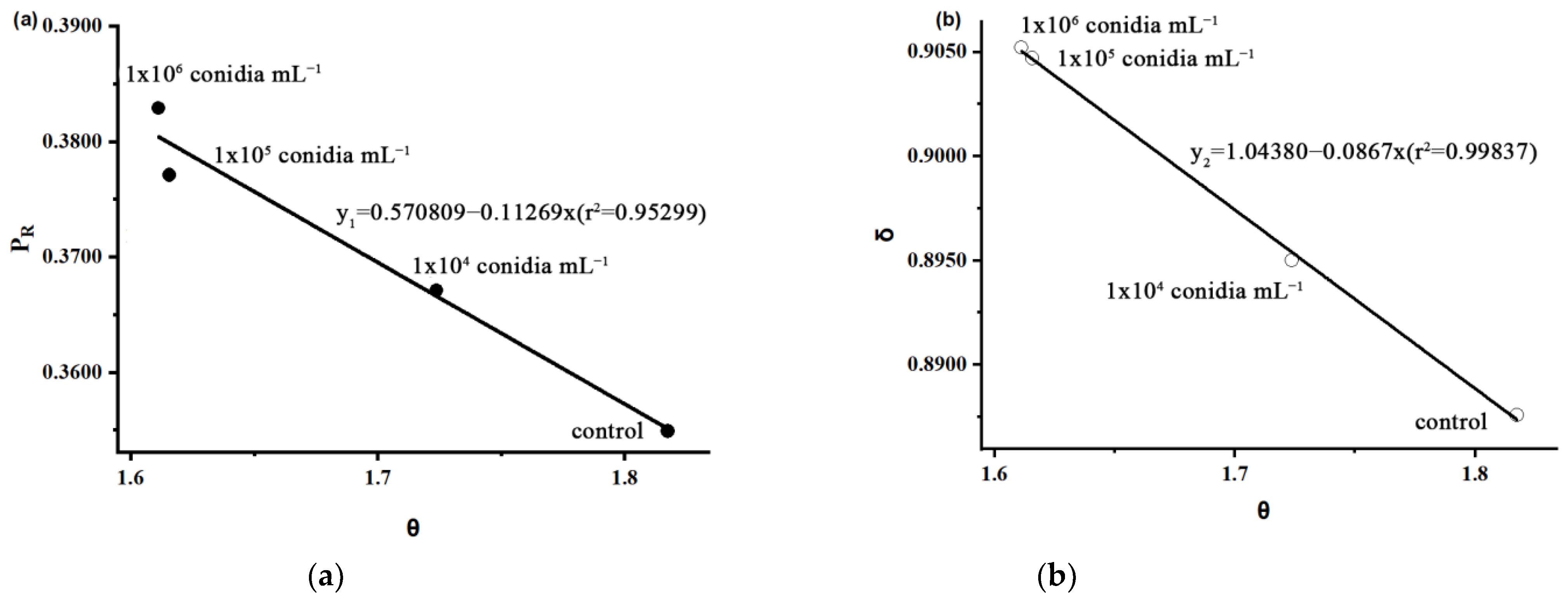
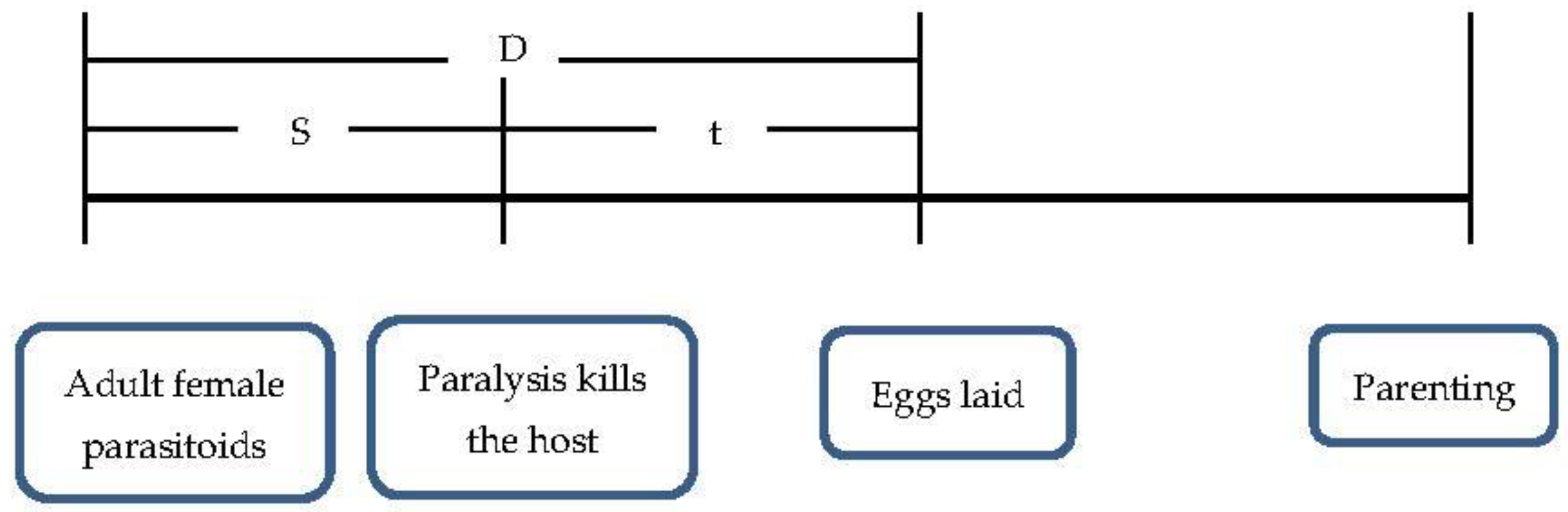
- D(d): the pre-oviposition of female adult parasitoids;
- S(d): the female adult parasitoids completely paralyze the host at the time;
- B(d): the time when the female adult parasitoid delivers a fatal blow to the host;
- t(d): the female adult parasitoids die after the host of the duration of egg laying;
- θ: the ratio of the time it takes for the female adult parasitoids to completely paralyze the host during the pre-oviposition of female adult parasitoids;
- PR: the parasitic probability of S. guani after paralyzing host M. alternatus larvae;
- δ: the antagonistic effect of S. guani and B. bassiana (the greater the antagonism, the worse the parasitoid, the weaker the lethal host effect).
2.5. Date Analysis
3. Results
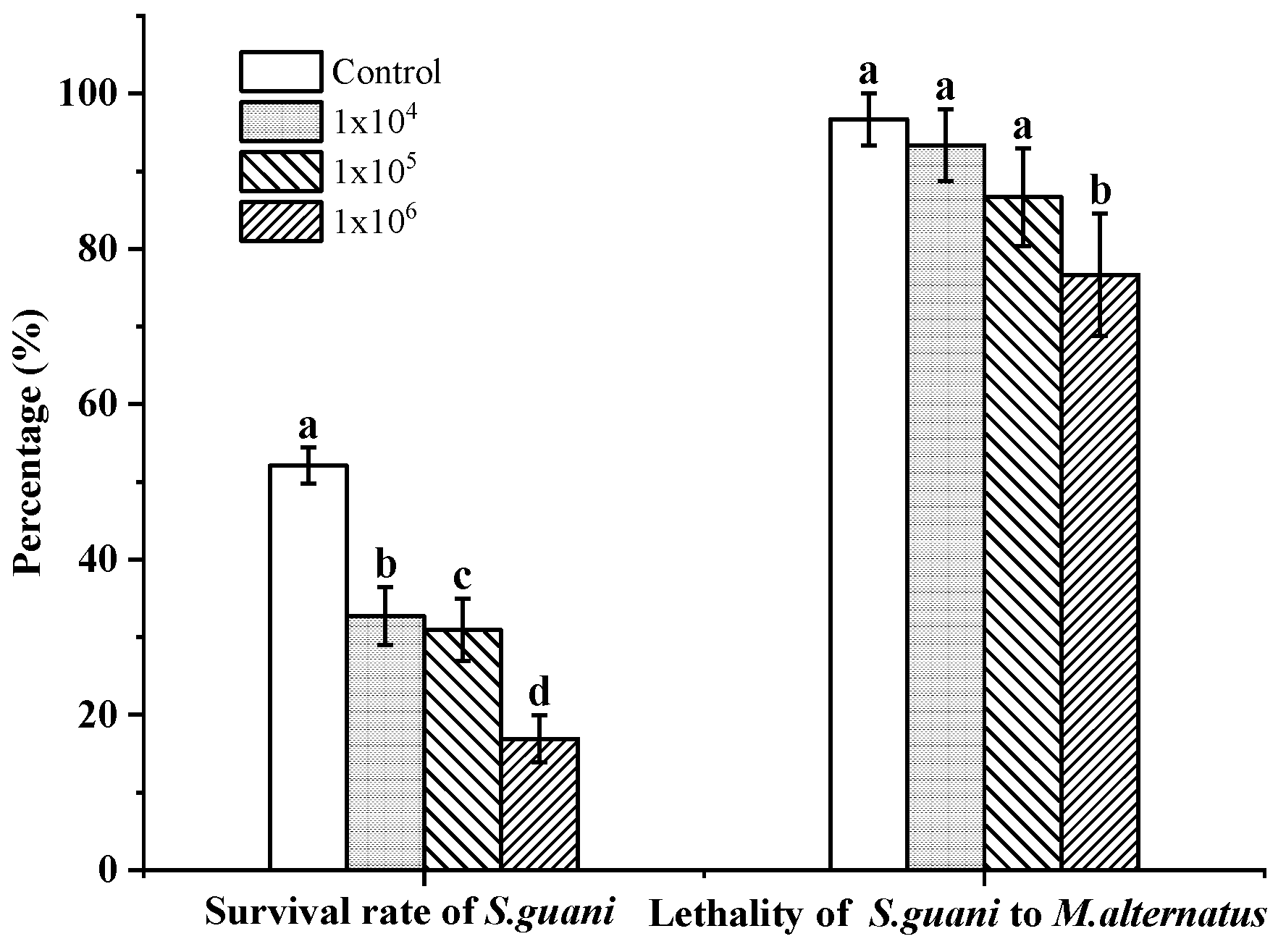
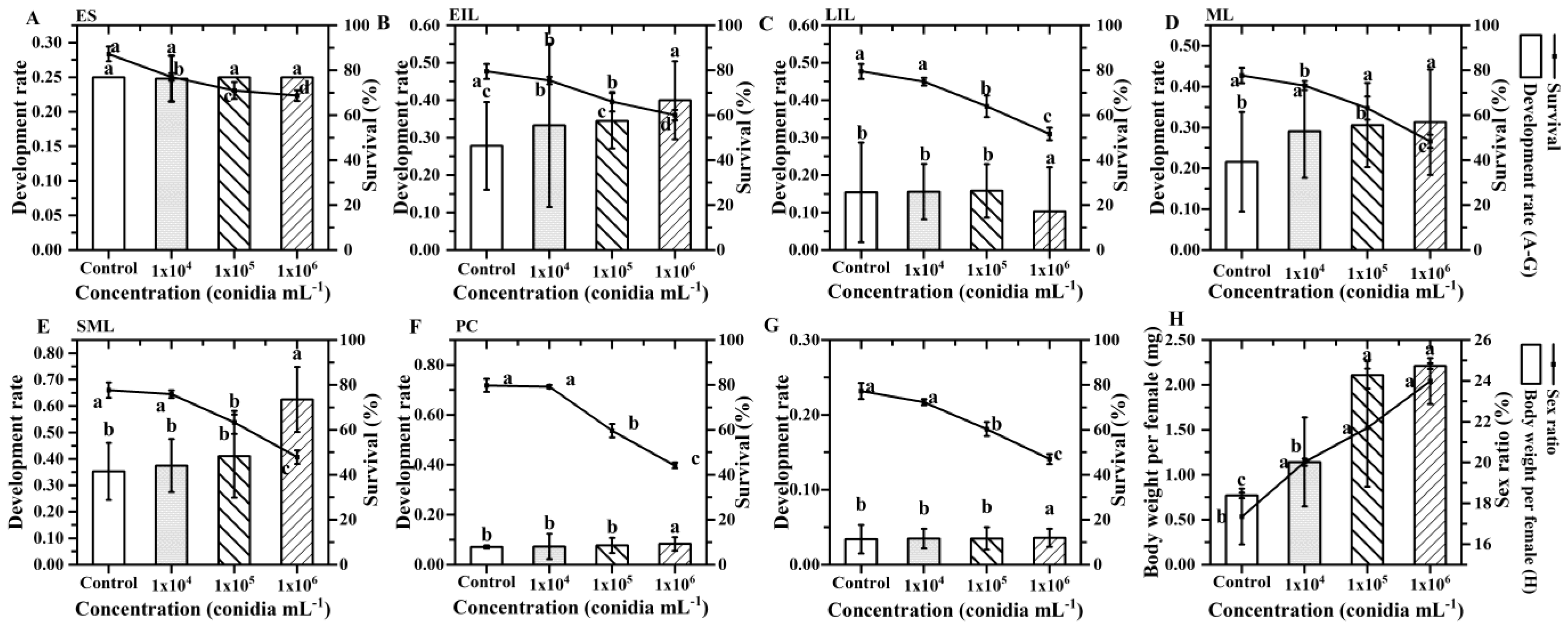
3.1. The Pathogenicity of B. bassiana to S. guani
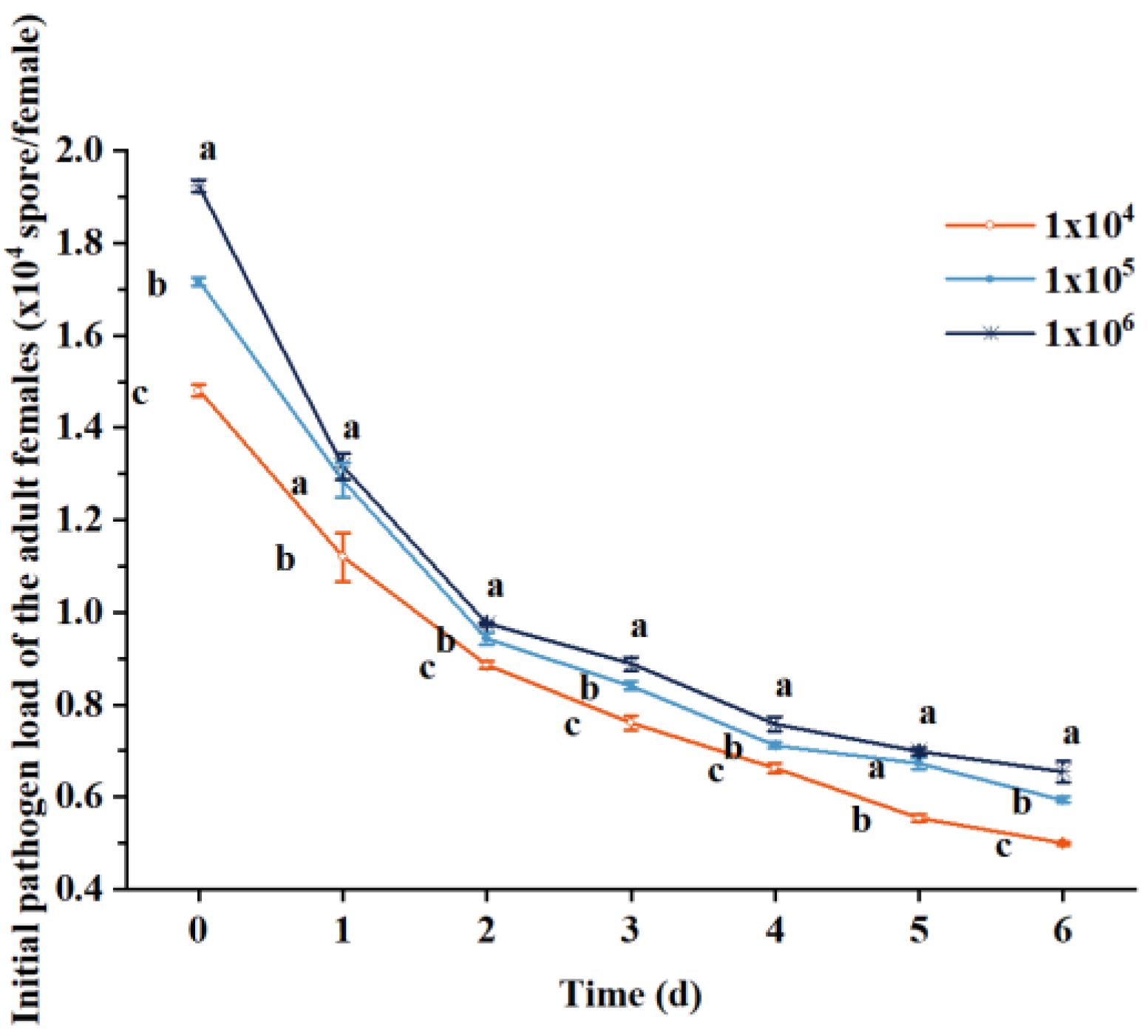
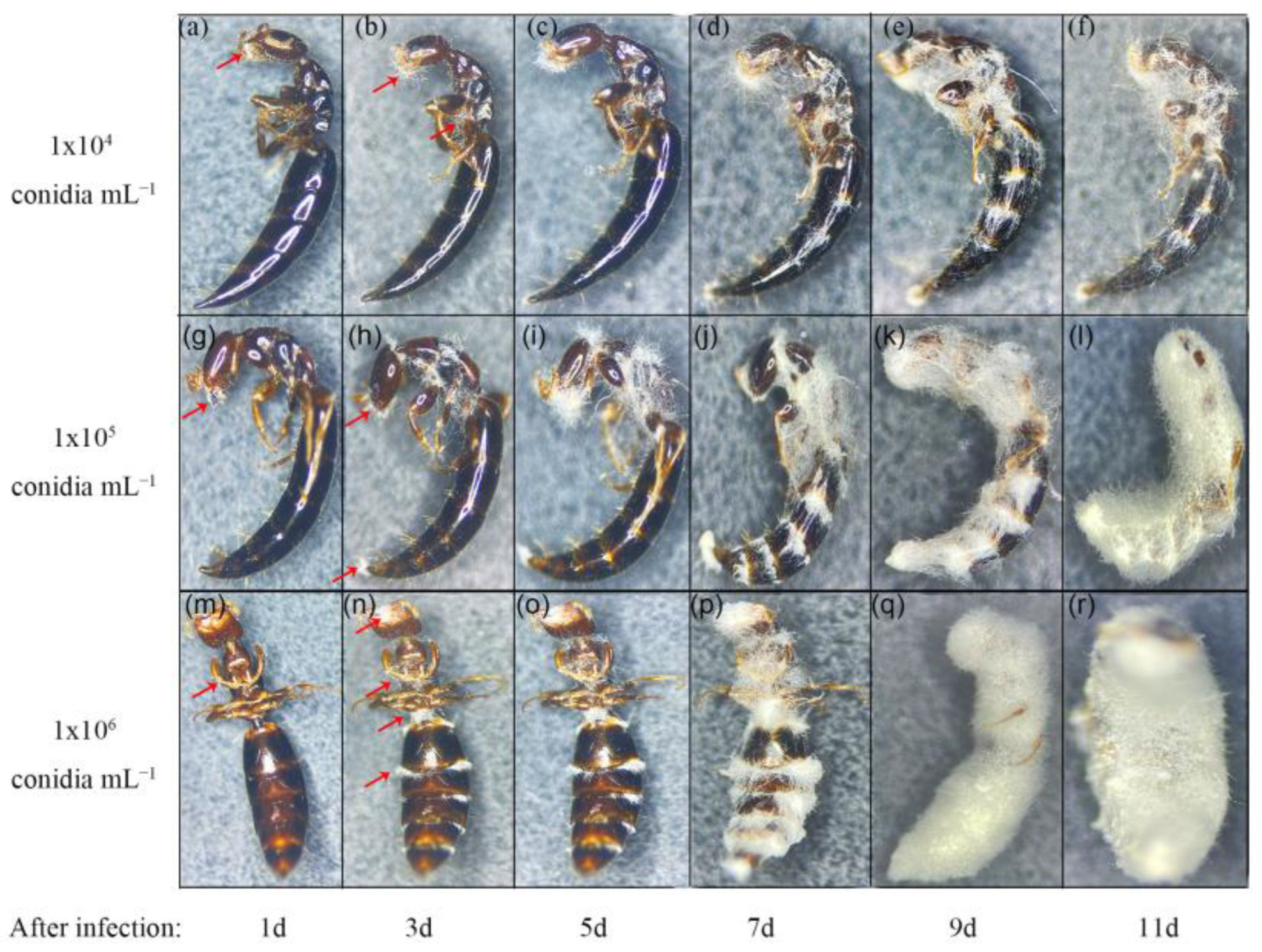
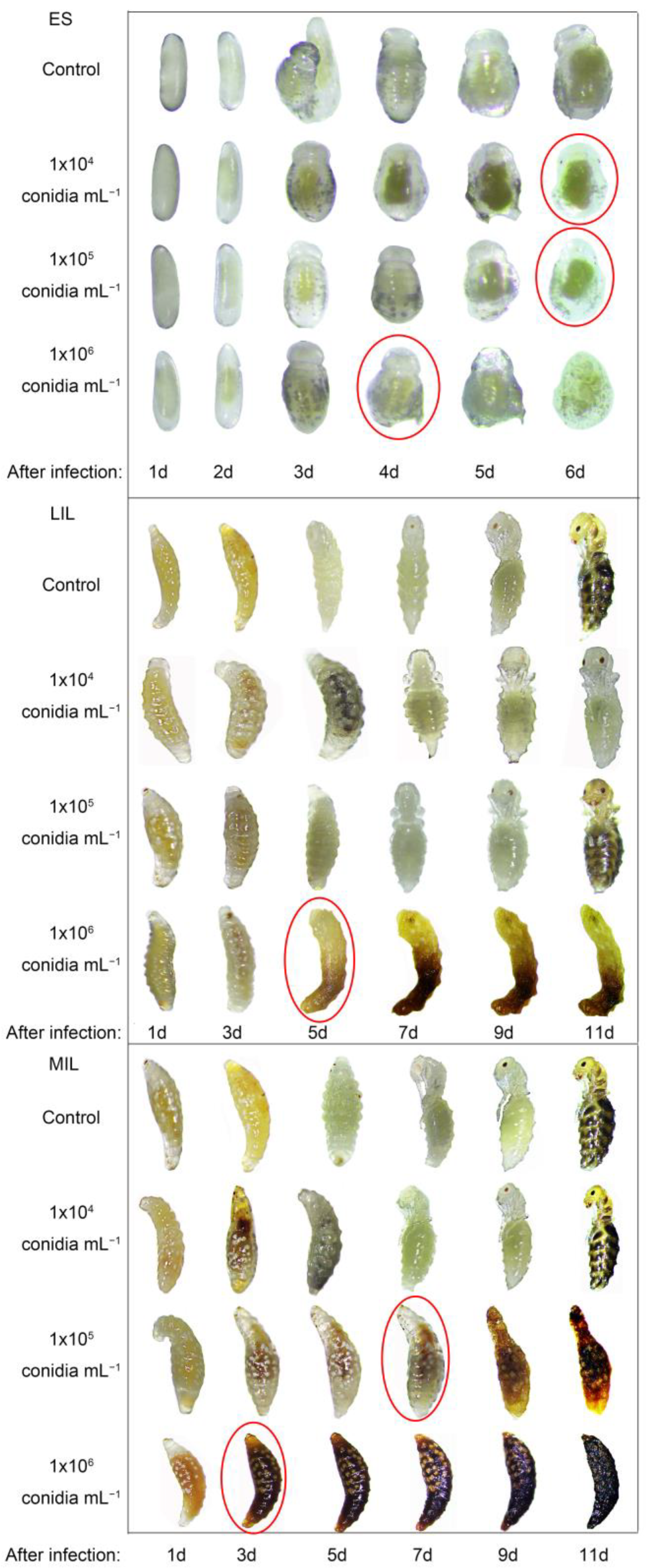
3.2. Pathogens Infect Immature Offspring
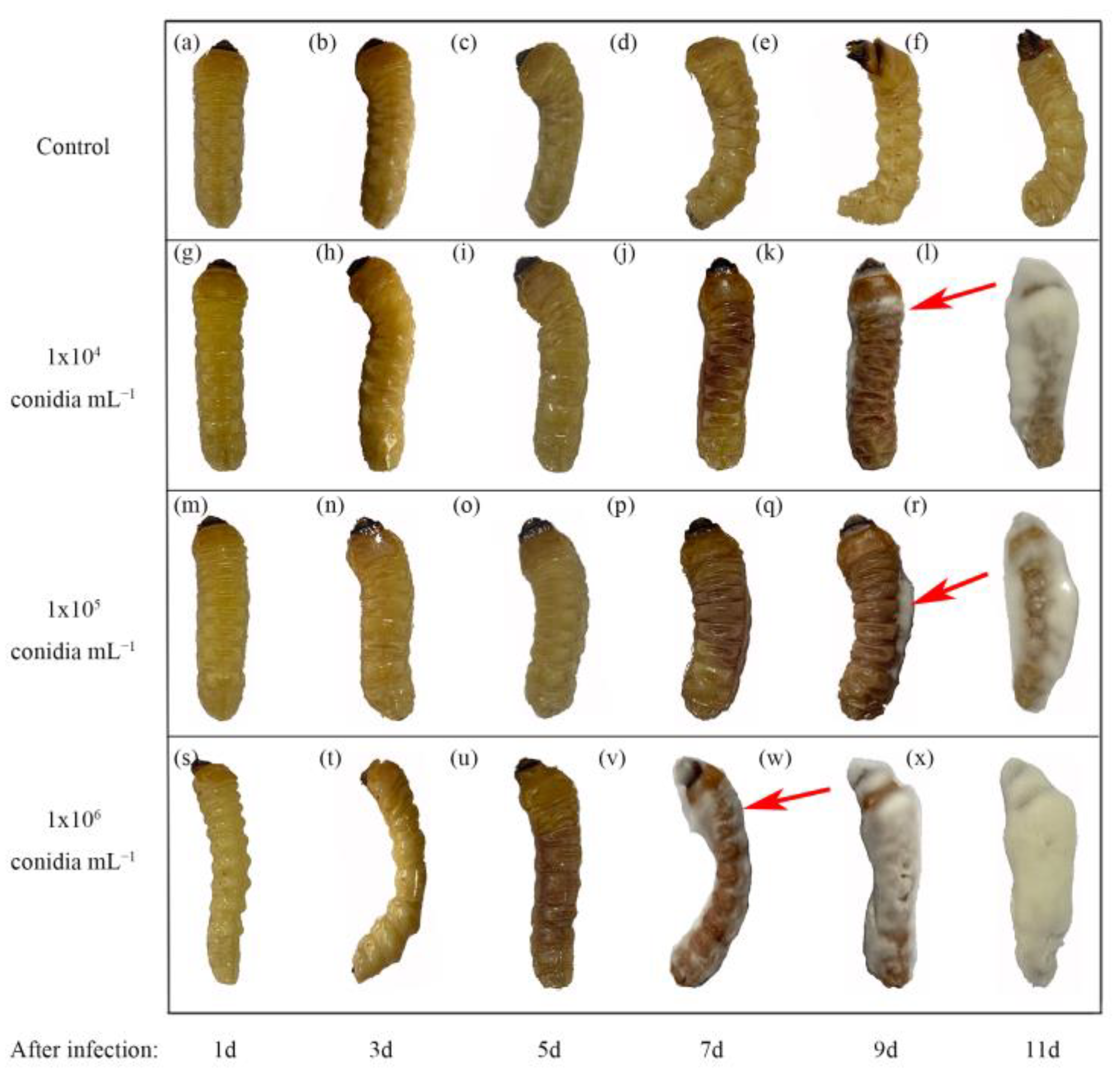
3.3. Pathogens Infect M. alternatus
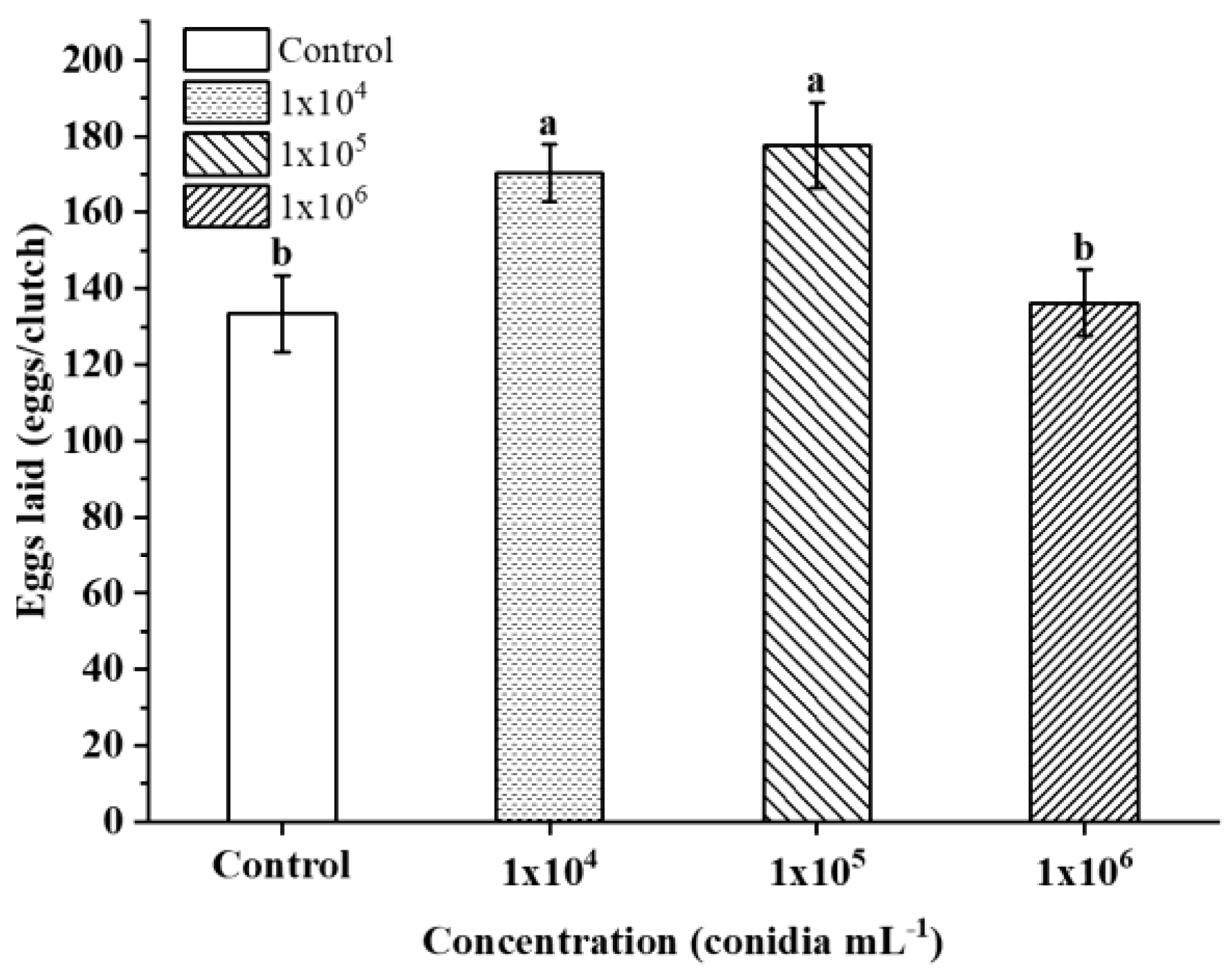
3.4. Mortality Effect of S. guani on the Host under B. bassiana Stress
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


Appendix B

References
- Rosenheim, J.A.; Kaya, H.K.; Ehler, L.E.; Marois, J.J.; Jaffee, B.A. Intraguild predation among biological-control agents: Theory and evidence. Biol. Control 1995, 5, 303–335. [Google Scholar] [CrossRef]
- Liu, H.J.; Su, Q.L.; Wang, L.F.; Piao, C.G.; Fang, J.M.; Dong, G.P. Research on the biocontrol of Monochamus alternatus larvae by Beauveria bassiana with several synergists. Forest. Res. 2009, 41, 225–232. [Google Scholar]
- Liu, Z.D.; Deng, L.L.; Wang, S.C.; Zheng, X.D.; Holyoak, M.; Wickham, J.D.; Tao, Y.; Sun, J.H. Mortality risk promotes cooperation of wasps when paralysing hosts. Anim. Behav. 2021, 172, 135–144. [Google Scholar] [CrossRef]
- Correa, M.C.G.; Palero, F.; Dubreuil, N.; Etienne, L.; Hulak, M.; Tison, G.; Warot, S.; Crochard, D.; Ris, N.; Kreiter, P. Molecular characterization of parasitoids from armored scales infesting citrus orchards in Corsica, France. Bio. Control 2016, 61, 639–647. [Google Scholar] [CrossRef]
- Kang, L. Overview: Biotic signalling for smart pest management. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Sun, J.H. Host suitability of a gregarious parasitoid on beetle hosts: Flexibility between fitness of adult and offspring. PLoS ONE 2011, 6, e18563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Liu, Z.D.; Sun, J.H. Olfactory cues in host and host-plant recognition of a polyphagous ectoparasitoid Sclerodermus guani. Bio. Control 2015, 60, 307–316. [Google Scholar]
- Zhao, L.L.; Mota, M.; Vieira, P.; Butcher, R.A.; Sun, J.H. Interspecific communication between pinewood nematode, its insect vector, and associated microbes. Trends. Parasitol. 2014, 30, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Jucker, C.; Hardy, I.C.W.; Malabusini, S.; de Milato, S.; Zen, G.; Savoldelli, S.; Lupi, D. Factors affecting the reproduction and mass-rearing of Sclerodermus brevicornis (Hymenoptera: Bethylidae), a natural enemy of exotic flat-faced Longhorn Beetles (Coleoptera: Cerambycidae: Lamiinae). Insects 2020, 11, 657. [Google Scholar] [CrossRef]
- Zhao, L.L.; Zhang, X.X.; Wei, Y.A.; Zhou, J.; Zhang, W.; Qin, P.J.; Chinta, S.; Kong, X.B.; Liu, Y.P.; Yu, H.Y.; et al. Ascarosides coordinate the dispersal of a plant-parasitic nematode with the metamorphosis of its vector beetle. Nat. Commun. 2016, 7, 12341. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Yang, Z.Q.; Wang, X.Y.; Hou, Y.X. Molecular identification of sibling species of Sclerodermus (Hymenoptera: Bethylidae) that parasitize buprestid and cerambycid beetles by using partial sequences of mitochondrial DNA cytochrome oxidase subunit 1 and 28S ribosomal RNA gene. PLoS ONE 2015, 10, e0119573. [Google Scholar] [CrossRef]
- Hu, Z.R.; Wu, X.Y.; Zhang, Y.N.; Han, Z.Q.; Zhao, C.X. The research advance on biological control of Scleroderma guani. J. Fujian. For. Sci. Technol. 2014, 41, 225–232. [Google Scholar]
- Li, J.Q.; Yan, Z.; Yang, Z.Q. Spatial distribution pattern and sampling technique of Batocera lineolata (Coleoptera: Cerambycidae) population in ash trees. Forest. Res. 2015, 28, 877–882. [Google Scholar]
- Wu, S.L.; Zhou, C.Z.; Peng, S.G.; Li, Y.H.; Li, B.P.; Meng, L. Influence of foundresss-offspring kinship on maternal care behavior and offspring developmental performance in Sclerodermus guani (Hymenoptera: Bethylidae). Acta. Entomol. Sin. 2017, 60, 1041–1045. [Google Scholar]
- Li, H.P.; Han, X.; Zhao, Y.Q. Potential use of Beauveria bassiana in combination with Sclerodermus guani for improved control of Apriona germari. Can. J. For. Res. 2016, 47, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.Y.; Li, L. Maternal care improves offspring developmental performance in Sclerodermus guani (Hymenoptera: Bethylidae). Acta Entomol. Sin. 2017, 60, 441–449. [Google Scholar]
- Guo, H.; Liu, Z.D.; Sun, J.H. Effects of spore suspension concentration and host body size on the pathogenicity of Beauveria bassiana against Monochamus alternatus (Coleoptera: Cerambycidae) larvae. Acta Entomol. Sin. 2020, 63, 835–842. [Google Scholar]
- Silva, R.J.; Alencar, J.R.; Silva, K.P.; Cividanes, F.J.; Duarte, R.T.; Agostini, L.T.; Polanczyk, R.A. Interactions between the entomopathogenic fungi Beauveria bassiana (Ascomycota: Hypocreales) and the aphid parasitoid Diaeretiella rapae (Hymenoptera: Braconidae) on Myzus persicae (Hemiptera: Aphididae). J. Econ. Entomol. 2014, 107, 933–938. [Google Scholar]
- Lai, Y.X.; Yang, Z.Q.; Wang, X.Y.; Wang, Y.H.; Chen, D.H.; Wei, J.R.; Tang, Y.L. Retarded develepment in Monochamus alternatus larva (Coleoptera: Cerambycidae) induced by parasitoid Sclerodermus guani (Hymenoptera: Bethylidae). Chin. J. Biol. Control 2015, 31, 305–311. [Google Scholar]
- Fu, B.; Tao, M.; Xue, H.; Jin, H.F.; Liu, K.; Qiu, H.Y.; Yang, S.Y.; Yang, X.; Gui, L.Y.; Zhang, Y.J.; et al. Spinetoram resistance drives interspecific competition between Megalurothrips usitatus and Frankliniella intonsa. Pest. Manag. Sci. 2022, 78, 2129–2140. [Google Scholar] [CrossRef]
- Ali, S.; Sajjad, A.; Shakeel, Q.; Farooqi, M.A.; Aqueel, M.A.; Tariq, K.; Ullah, M.I.; Iqbal, A.; Jamal, A.; Saeed, M.F.; et al. Influence of bacterial secondary symbionts in Sitobion avenae on its survival fitness against entomopathogenic fungi, Beauveria bassiana and Metarhizium brunneum. Insects 2022, 13, 1037. [Google Scholar] [CrossRef] [PubMed]
- Abdi, M.K.; Jucker, C.; Marchi, B.D.; Hardy, I.C.W.; Lupi, D. Performance of Sclerodermus brevicornis, a parasitoid of invasive longhorn beetles, when reared on rice moth larvae. Entomol. Exp. Appl. 2020, 169, 64–78. [Google Scholar] [CrossRef]
- Li, T.; Liu, Y.H.; Li, H.Y.; Zhang, Y.J. Impacts of Beauveria bassiana on life parameters and control efficiency of Aphidius gifuensis. Mycosystema 2007, 26, 436–439. [Google Scholar]
- Huang, Z.D.; Wan, Q.; Zhuang, G.F.; Xue, Z.J.; Zhang, R.L.; Zhang, Z. Synergestic control efficacy of Nasonia vitripennis and Beauveria bassiana against Musca domestica. Chin. J. Vector. Biol. Control 2018, 29, 336–339. [Google Scholar]
- Castillo-Ramírez, O.; Guzmán-Franco, A.W.; Santillán-Galicia, M.; Tamayo-Mejía, F. Interaction between predatory mites (Acari: Phytoseiidae) and entomopathogenic fungi in Tetranychus urticae populations. Bio. Control 2020, 65, 433–445. [Google Scholar] [CrossRef]
- Deng, J.D.; Zhuang, W.X.; Liu, Y.J.; Song, L.W.; Zhang, L.W. Pathogenicity of white muscardine fungus Beauveria bassiana against Japanese pine sawyer beetle Monochamus alternatus and its compatibility with ectoparasitic beetle Dastarcus helophoroides. J. Plant. Prot. 2021, 48, 602–609. [Google Scholar]
- Mesterton-Gibbons, M.; Hardy, I.C.W. Defection on the bounty? Kinship and cooperative exploitation of a rich, essential but dangerous resource. Anim. Behav. 2021, 176, 57–65. [Google Scholar] [CrossRef]
- Li, Y.X.; Liu, S.S. Effects of insecticides on parasitoids. Entomol. Knowl. 2001, 1, 26–30. [Google Scholar]
- Wang, G.H.; Shen, J.K. Studes on the cold storage and functional response of Pediobius Foveolatus (Crawford). Acta Agric. Univ. Jiangxiensis 1992, 4, 383–389. [Google Scholar]
- Herzner, G.; Engl, T.; Strohm, E. Cryptic combat against competing microbes is a costly component of parental care in a digger wasp. Anim. Behav. 2011, 82, 321–328. [Google Scholar] [CrossRef]
- Tang, X.Y. Adaptation of Parental Care and Sex Allocation in the Parasitoid Sclerodermus guani (Hymenoptera: Bethylidae). Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2015. [Google Scholar]
- Niu, G.S.; Li, L.; Tang, M. Effects of parasite density on the parasitism and reproduction efficiency of Sclerodermus guani (Hymenoptera: Bethylidae). J. Environ. Entomol 2022, 1–20. [Google Scholar]
- Villena, O.C.; Sullivan, J.H.; Landa, E.R.; Yarwood, S.A.; Torrents, A.; Zhang, A.; Leisnham, P.T. The role of tire leachate in condition-specific competition and the persistence of a c from a competitively superior invader. Insects 2022, 13, 969. [Google Scholar] [CrossRef]
- Purkiss, S.A.; Khudr, M.S.; Aguinaga, O.E.; Hager, R. Symbiont-conferred immunity interacts with effects of parasitoid genotype and intraguild predation to affect aphid immunity in a clone-specific fashion. BMC Ecol. Evol. 2022, 22, 33. [Google Scholar] [CrossRef] [PubMed]
- Cantori, L.V.; Garcia, A.G.; Pinto, A.S.; Godoy, W.A.C.; Parra, J.R.P. Detailed look at paralysis of hosts by the ectoparasitoid Habrobracon hebetor (Hymenoptera: Braconidae): Does more efficient paralysis mean more effective parasitism? Bio. Control 2022, 67, 555–562. [Google Scholar] [CrossRef]
- Gross, P. Insect behavioral and morphological defenses against parasitoids. Annu. Rev. Entomol. 1993, 38, 251–273. [Google Scholar] [CrossRef]
- Godfray, H.C.J. “Introduction.” Parasitoids: Behavioral and Evolutionary Ecology; Princeton University Press: Princeton, NJ, USA, 1994; pp. 3–25. [Google Scholar]
- Abdi, M.K.; Hardy, I.C.W.; Jucker, C.; Lupi, D. Kinship effects in quasi-social parasitoids II: Co-Foundress relatedness and host dangerousness interactively affect host exploitation. Bot. J. Linn. Soc. 2020, 130, 642–660. [Google Scholar] [CrossRef]
- Holt, R.D.; Polis, G.A. A theoretical framework for intraguild predation. Am. Nat. 1997, 149, 745–764. [Google Scholar] [CrossRef]
- Wei, Y.; Li, L.; Zhang, M.M.; Pan, S.M. The intergenerational effects of entomopathogen Beauveria bassiana on mortality and parasitism of ectoparasitoid Sclerodermus guani (Hymenoptera: Bethylidae). Acta. Entomol. Sin. 2023, in press. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentrations (conidia mL−1) | Parameters | LE | PE | |||||
|---|---|---|---|---|---|---|---|---|
| D(d) | S(d) | t(d) | θ | PR | δ | % | % | |
| Control | 6.67 | 3.67 | 3.00 | 1.8174 | 0.3549 | 0.8876 | 90.00 | 83.33 |
| 1 × 104 | 5.93 | 3.44 | 2.49 | 1.7238 | 0.3671 | 0.8950 | 93.33 | 86.67 |
| 1 × 105 | 5.17 | 3.20 | 1.97 | 1.6156 | 0.3771 | 0.9047 | 93.33 | 93.33 |
| 1 × 106 | 5.00 | 3.10 | 1.90 | 1.6112 | 0.3892 | 0.9052 | 100.00 | 96.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, Y.; Li, L.; Pan, S.; Liu, Z.; Fan, J.; Tang, M. Adaptive Reproductive Strategies of an Ectoparasitoid Sclerodermus guani under the Stress of Its Entomopathogenic Fungus Beauveria bassiana. Insects 2023, 14, 320. https://doi.org/10.3390/insects14040320
Wei Y, Li L, Pan S, Liu Z, Fan J, Tang M. Adaptive Reproductive Strategies of an Ectoparasitoid Sclerodermus guani under the Stress of Its Entomopathogenic Fungus Beauveria bassiana. Insects. 2023; 14(4):320. https://doi.org/10.3390/insects14040320
Chicago/Turabian StyleWei, Yun, Li Li, Shumei Pan, Zhudong Liu, Jianting Fan, and Ming Tang. 2023. "Adaptive Reproductive Strategies of an Ectoparasitoid Sclerodermus guani under the Stress of Its Entomopathogenic Fungus Beauveria bassiana" Insects 14, no. 4: 320. https://doi.org/10.3390/insects14040320