

Measuring and Modelling Structural Colours of Euphaedra neophron (Lepidoptera: Nymphalidae) Finely Tuned by Wing Scale Lower Lamina in Various Subspecies
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Species and Specimens
2.2. Optical Microscopy
2.3. Scanning Electron Microscopy (SEM)
2.4. Spectroscopy
2.5. Modelling
3. Results
3.1. Scale Structures
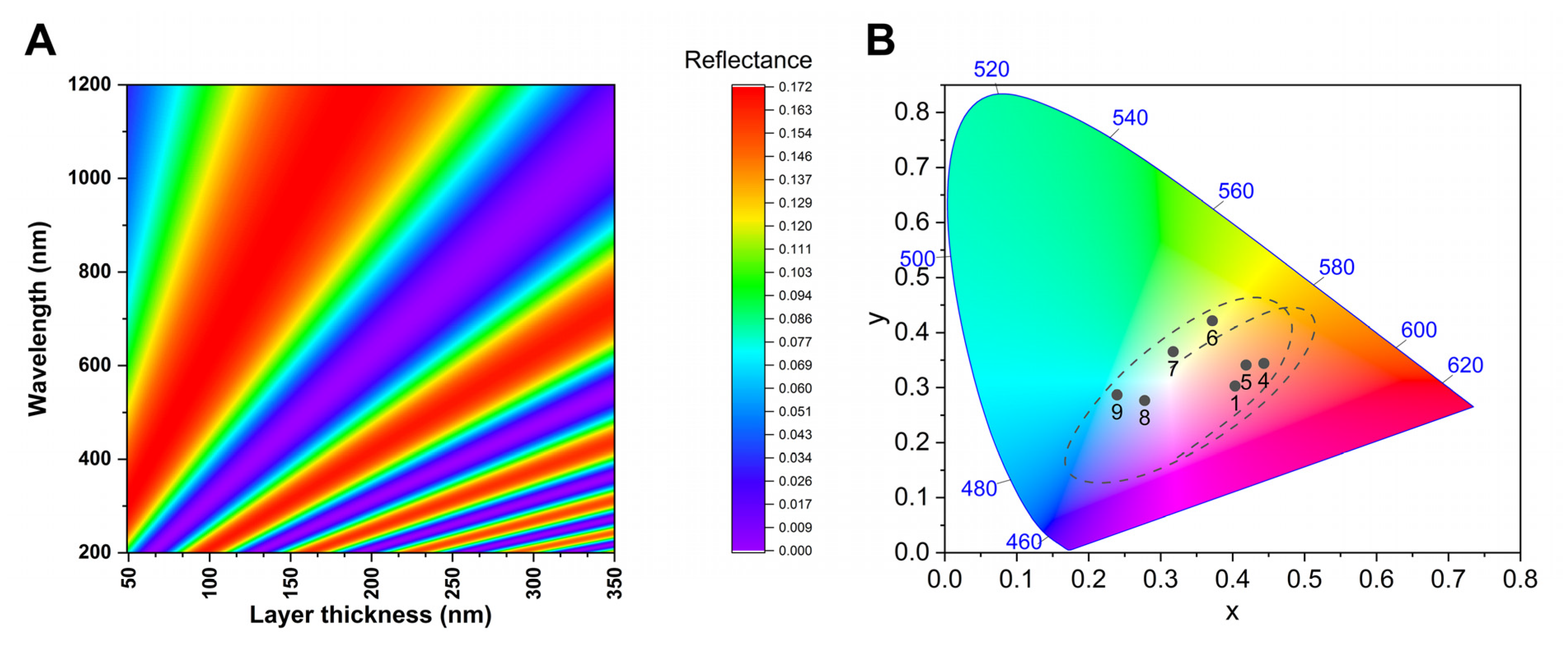
3.2. Spectral Properties
3.3. Modelling
3.4. Morph Colour Tuning
4. Discussion
4.1. Former Observations
4.2. Sexual Dimorphism
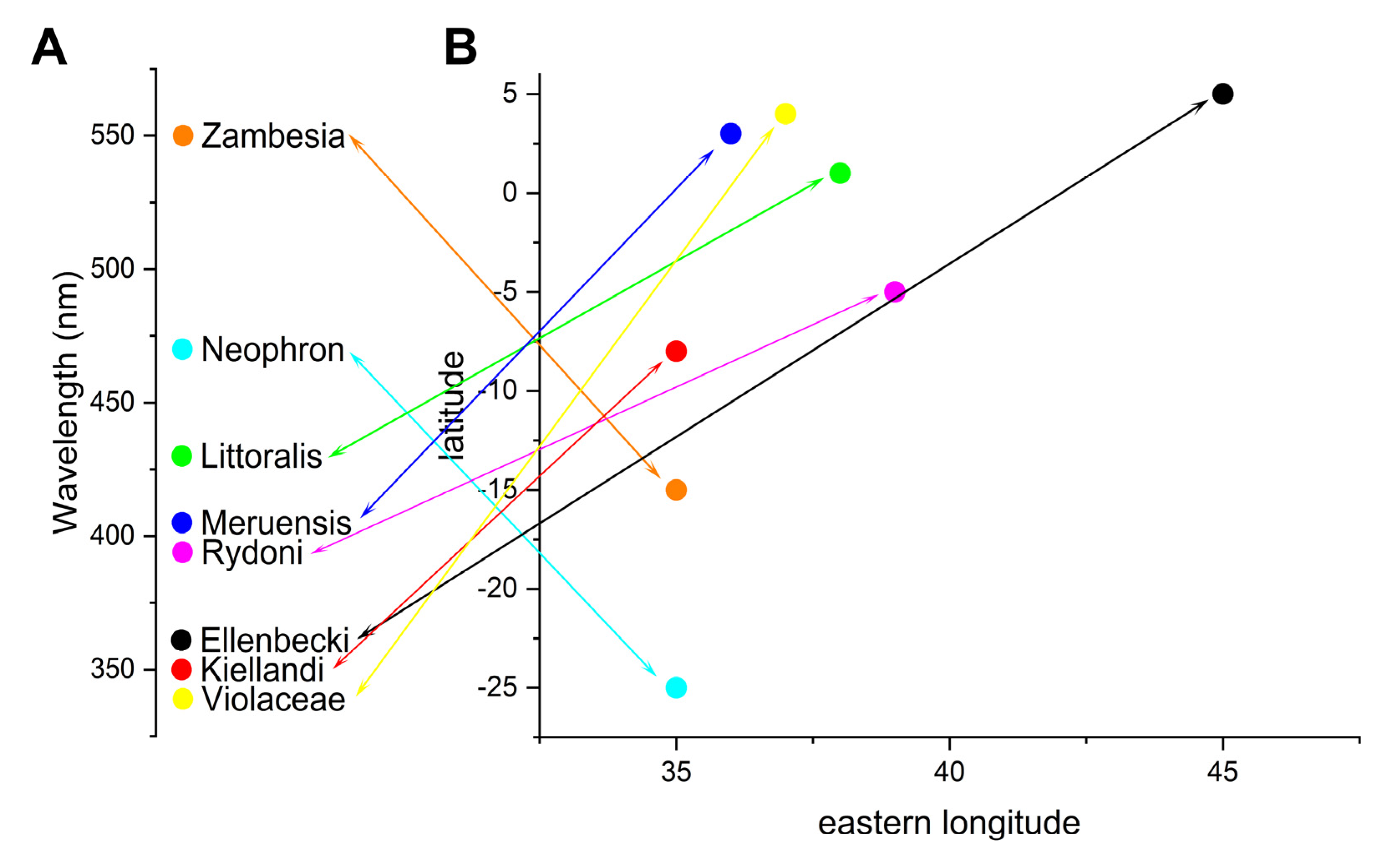
4.3. Diversification and Colour Tuning Mechanism
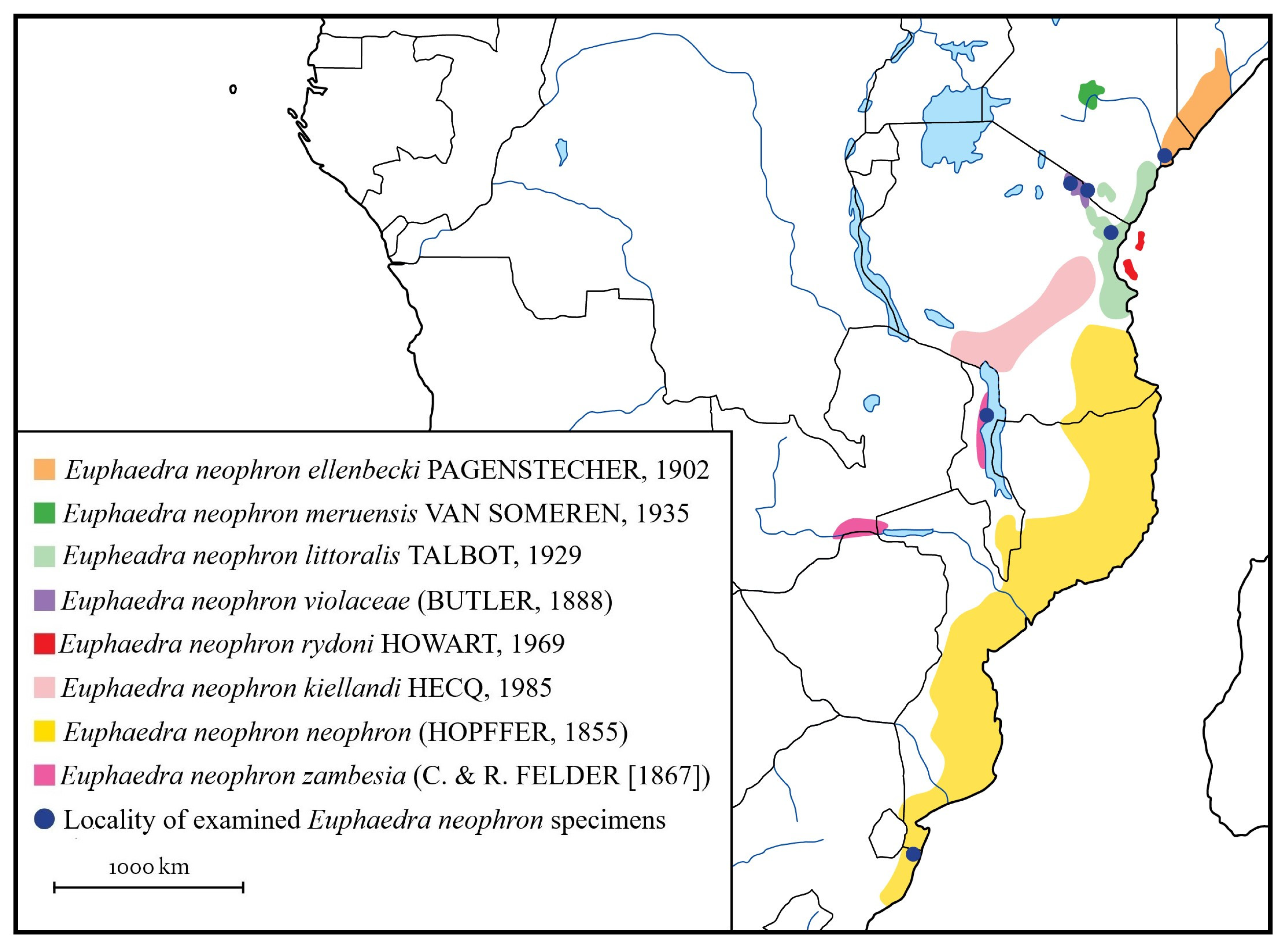
4.4. Biogeographical Background
4.5. Nomenclature
4.6. Systematics
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dunghel, B.; Wahlberg, N. Molecular systematics of the subfamily Limenitidinae (Lepidoptera: Nymphalidae). PeerJ 2018, 6, e4311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- d’Abrera, B. Butterflies of the Afrotropical Region. Part II. Nymphalidae, Libytheidae. New and Revised Edition; Hill House Publishers: Melbourne, Australia; London, UK, 2004; pp. i–xl, 257–539. [Google Scholar]
- Larsen, T.B. The Butterflies of West Africa; Apollo Books: Stenstrup, Denmark, 2005; 865p. [Google Scholar] [CrossRef]
- Hecq, J. Euphaedra; Lambillionea: Namur, Belgium, 1999; 126p. [Google Scholar]
- White, E.T.; Macedonia, J.; Birch, B.; Dawes, J.; Kemp, J.K. The nanoanatomical basis of sexual dimorphism in iridescent butterfly colouration. Aust. J. Zool. 2012, 60, 101–107. [Google Scholar] [CrossRef]
- Kertész, K.; Piszter, G.; Bálint, Z.; Biró, L.P. Biogeographical patterns in the structural blue of male Polyommatus icarus butterflies. Sci. Rep. 2019, 9, 2338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bálint, Z.; Parker, A.; Ingram, A.; Kertész, K.; Piszter, G.; Horváth, Z.E.; Illés, L.; Biró, L.P. Scale granules and colours: Sexual dimorphism in Trichonis (Lepidoptera: Lycaenidae, Theclinae). Arthropod. Struct. Dev. 2021, 65, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Larsen, T.B. Butterflies of Kenya and Their Natural History; Oxford University Press: Oxford, UK, 1991; xxii + 500p. [Google Scholar]
- Ackery, R.P.; Smith, R.C.; Vane-Wright, I.R. Carcasson’s African Butterflies: An Annotated Catalogue of the Papilionoidea nd Hesperioidea of the Afrotropical Region; CSIRO: East Melbourne, Australia, 1995; pp. 1–803. [Google Scholar] [CrossRef]
- Pendry, J.B.; MacKinnon, A. Calculation of photon dispersion relations. Phys. Rev. Lett. 1992, 69, 2772–2775. [Google Scholar] [CrossRef]
- Balili, R.B. Transfer Matrix Method in Nanophotonics. Int. J. Mod. Phys. Conf. Ser. 2012, 17, 159–168. [Google Scholar] [CrossRef]
- Ghiradella, H.T. Structure and development of iridescent butterfly scales: Lattices and laminae. J. Morphol. 1989, 202, 69–88. [Google Scholar] [CrossRef]
- Kristensen, N.P.; Simonsen, J.T. Hairs and scales. In Lepidoptera, Moths and Butterflies. Volume 2: Morphology, Physiology, and Development. Handbuch der Zoologie; Kristensen, N.P., Ed.; Walter de Gruyter: Berlin, Germany; New York, NY, USA, 2003; Volume 4, pp. 9–22. [Google Scholar] [CrossRef]
- Stavenga, D.G.; Leertouwer, H.L.; Wilts, B.D. Coloration principles of nymphaline butterflies—Thin films, melanin, ommochromes and wing scale stacking. J. Exp. Biol. 2014, 217, 2171–2180. [Google Scholar] [CrossRef] [Green Version]
- Leertouwer, H.L.; Wilts, B.D.; Stavenga, D.G. Refractive index and dispersion of butterfly chitin and bird keratin measured by polarizing interference microscopy. Opt. Express 2011, 19, 24061–24066. [Google Scholar] [CrossRef]
- Bálint, Z.; Sáfián, S.; Hoskins, A.; Kertész, K.; Koós, A.A.; Horváth, E.Z.; Piszter, G.; Biró, L.P. The Only Blue Mimeresia (Lepidoptera: Lycaenidae: Lipteninae) Uses a Color-Generating Mechanism Widely Applied by Butterflies. J. Insect Sci. 2018, 18, 1–6. [Google Scholar] [CrossRef]
- Bálint, Z.; Kertész, K.; Piszter, G.; Vértesy, Z.; Biro, L.P. The well-tuned blues: The role of structural colours as optical signals in the species recognition of a local butterfly fauna (Lepidoptera: Lycaenidae: Polyommatinae). J. R. Soc. Interface 2012, 9, 1745–1756. [Google Scholar] [CrossRef] [Green Version]
- Parmentier, L.; Vila, R.; Lukhtanov, V. Integrative analysis reveals cryptic speciation linked to habitat differentiation within Albanian populations of the anomalous blues (Lepidoptera, Lycaenidae, Polyommatus Latreille, 1804). Comp. Cytogenet. 2022, 16, 211–242. [Google Scholar] [CrossRef]
- Gralak, B.; Tayeb, G.; Enoch, S. Morpho butterflies wing color modeled with lamellar grating theory. Opt. Express 2001, 9, 567–578. [Google Scholar] [CrossRef]
- Cao, X.; Du, Y.; Guo, Y.; Hu, G.; Zhang, M.; Wang, L.; Zhou, J.; Gao, Q.; Fischer, P.; Wang, J.; et al. Replicating the Cynandra opis Butterfly’s Structural Color for Bioinspired Bigrating Color Filters. Adv. Mater. 2022, 34, 2109161. [Google Scholar] [CrossRef]
- Wasik, R.B.; Liew, F.S.; Lilien, A.D.; Dinwiddie, J.A.; Noh, H.; Cao, H.; Monteiro, A. Artificial selection for structural color on butterfly wings and comparison with natural evolution. Proc. Natl. Acad. Sci. USA 2014, 111, 12109–12114. [Google Scholar] [CrossRef] [Green Version]
- Parnell, A.J.; Bradford, J.E.; Curran, E.V.; Washington, A.L.; Adams, G.; Brien, M.N.; Burg, S.L.; Morochz, C.; Fairclough, J.P.A.; Vukusic, P.; et al. Wing scale ultrastructure underlying convergent and divergent iridescent colours in mimetic Heliconius butterflies. J. R. Soc. Interface 2018, 15, 20170948. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Acevedo, E.; Gomez, P.J.; Espeland, M.; Toussaint, A.F.E.; Willmott, R.K. The roles of wing color pattern and geography in the evolution of Neotropical Preponini butterflies. Ecol. Evol. 2020, 10, 12801–12816. [Google Scholar] [CrossRef]
- Thayer, C.R.; Allen, I.F.; Patel, H.N. Structural color in Junonia butterflies evolves by tuning scale lamina thickness. eLife 2020, 9, e52187. [Google Scholar] [CrossRef]
- Axelrod, D.I.; Raven, P.H. Late Cretaceous and Tertiary vegetation history of Africa. In Biogeography and Ecology of Southern Africa; Werger, M.J.A., Ed.; Springer: Dordrecht, The Netherlands, 1978; pp. 77–130. [Google Scholar] [CrossRef]
- Mumbi, C.; Marchant, R.; Hooghiemstra, H.; Wooller, M. Late Quaternary vegetation reconstruction from the Eastern Arc Mountains, Tanzania. Quat. Res. 2008, 69, 326–341. [Google Scholar] [CrossRef]
- Hamilton, A.C.; Taylor, D. History of Climate and Forests in Tropical Africa during the Last 8 million years. In Tropical Forests and Climate; Myers, N., Ed.; Springer: Dordrecht, The Netherlands, 1991; pp. 65–78. [Google Scholar] [CrossRef]
- Congdon, C.; Collins, S.C.; Bayliss, J. Butterflies of south east Africa’s mountains (Mozambique and Malawi). Metamorphosis 2010, 21, 46–106. [Google Scholar]
- Congdon, T.C.E.; Bayliss, J. Butterflies of Mt Mecula and Mt Yao, Niassa Province, Northern Mozambique. Metamorphosis 2013, 23, 26–34. [Google Scholar]
- Felder, C.; Felder, R. Reise der Österreichischen Fregatte Novara um Die Erde in den Jahren 1857, 1858, 1859 Unter den Befehlen des Commodore B. von Wüllerstorf-Urbair. Zoologischer Theil. Zweiter Band. Zweite Abtheilung: Lepidoptera; Carl Gerold’s Sohn: Wien, Austria, 1865; (1): [4] + 1–136, pp. 1–21; (2): [2] + 137–378, pp. 22–47. [Google Scholar]
- Bálint, Z.; Goodger, K. The Neotropical eumaeine lycaenids of the Cajetan and Rudolf Felder collection. Part I. Review of type material of the taxa described by the Felders (Lepidoptera: Lycaenidae). Quadrifina 2003, 6, 67–140. [Google Scholar]
- Chainey, J.E. The species of Papilionidae and Pieridae (Lepidoptera) described by Cramer and Stoll and their putative typematerial in the Natural History Museum in London. Zool. J. Linn. Soc. 2005, 145, 283–337. [Google Scholar] [CrossRef] [Green Version]
- Hecq, J. Étude des Euphaedra. Euphaedra neophron Hop. et remarques diverses (Note 26). Lambillionea 1985, 85, 7–15. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | General Distribution | Type Locality |
|---|---|---|
| Euphaedra neophron ellenbecki (Pagenstrecher, 1902) | coast of northern Kenya and southern Somalia | “Umfudu” and “Evar” (Somalia) |
| Euphaedra neophron kielland (Hecq, 1985) | southern Tanzania | “Tanzanie, Ifakara, Musagabi” (= Ifakara, Masagati; Tanzania) |
| Euphaedra neophron littoralis (Talbot, 1929) | northern coast of Tanzania across the remaining patches of coastal of Kenya (to about Malindi) | “Rabai” (= Rabai Mpya (New Rabai); Kenya) |
| Euphaedra neophron meruensis (van Someren, 1935) | eastern slopes of Mount Kenya (Meru) | “Meru Forest” (Kenya) |
| Euphaedra neophron neophron (Hopffer, 1855) | coast regions from eastern South Africa via Mozambique to southern Kenya | “Mossambique” (Mosambique) |
| Euphaedra neophron rydoni (Howarth, 1969) | Pemba Island | “Pemba Island” (Tanzania) |
| Euphaedra neophron violaceae (Butler, 1888) | Usambaras and the South Pare Mountains in the Eastern Arc Mountains, also on Mount Meru and Mount Kilimanjaro in north Tanzania, and in Taveta, in the nearby Kenya | “Kilimanjaro” (Kenya and Tanzania) |
| Euphaedra neophron zambesia (Felder and Felder, 1865) | eastern Zimbabwe, Malawi, western Mozambique | “Zambia” (Zambia) |
| Scientific Trinomen | Sex | Working Number | Original Data of the Specimen (Depositor) | Resolved Data (with Coordinates) |
|---|---|---|---|---|
| Euphaedra neophron violaceae | male | (1) | Kibosho, Kitty, coll. Velez; Tanzania, Mkoringa, leg. Kittenberger (HNHM) | (3°14′40.92″ S, 37°18′8.14″ E; altitude: 1281 m |
| Euphaedra neophron violaceae | male | (2) | Moshi, Kilimanjaro, 1936.IV.; coll. Velez (HNHM) | Tanzania, Moshi (3°20′40.89″ S, 37°20′50.59″ E; altitude: 800 m |
| Euphaedra neophron violaceae | male | (3) | Tanganyika, 1934. IV.; coll. Velez (HNHM) | Tanzania, Tanganyika (inaccurate locality) |
| Euphaedra neophron violaceae | female | (8) | Kenia, Kilimanjaro, 1913, VI.; coll. Velez (HNHM) | Kenya, Kilimanjaro (inaccurate locality) |
| Euphaedra neophron violaceae | male | (5) | Kenya, Kitobo Forest Taveta, 1977. III., S.C. Collins (ABRI) | Kenya, Kitavo (3°23′57.03″ S, 37°40′37.50″ E, altitude: 700 m) |
| Euphaedra neophron ellenbecki | male | (4) | Kenya, Witu, 400 ft, K 1983., S. C. Collins (ABRI) | (2°23′4.35″ S, 40°28′30.18″ E; altitude: 22 m) |
| Euphaedra neophron zambesia | male | (6) | Malawi, Nkhata Bay, 50 ft, 1983. III, S. C. Collins (ABRI) | (11°13′12.8″ S 34°16′44.5″ E; altitude: 500 m) |
| Euphaedra neophron neophron | male | (7) | South-Africa, Natal, Manguze, 1991.VII.12., A. Hanekom (ABRI) | South-Africa, Manguzi Forest (16°59′51.56″ S, 32°43′20.54″ E; altitude: 72 m) |
| Euphaedra neophron littoralis | male | (9) | Tanzania, East Usambara, forest below Camp Mawingu, 15.V.2022. Leg.: Sáfián, Sz. (HNHM) | (25°8′16.41″ S, 38°34′48.80″ E; altitude: 770 m) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bálint, Z.; Katona, G.; Sáfián, S.; Collins, S.; Piszter, G.; Kertész, K.; Biró, L.P. Measuring and Modelling Structural Colours of Euphaedra neophron (Lepidoptera: Nymphalidae) Finely Tuned by Wing Scale Lower Lamina in Various Subspecies. Insects 2023, 14, 303. https://doi.org/10.3390/insects14030303
Bálint Z, Katona G, Sáfián S, Collins S, Piszter G, Kertész K, Biró LP. Measuring and Modelling Structural Colours of Euphaedra neophron (Lepidoptera: Nymphalidae) Finely Tuned by Wing Scale Lower Lamina in Various Subspecies. Insects. 2023; 14(3):303. https://doi.org/10.3390/insects14030303
Chicago/Turabian StyleBálint, Zsolt, Gergely Katona, Szabolcs Sáfián, Steve Collins, Gábor Piszter, Krisztián Kertész, and László Péter Biró. 2023. "Measuring and Modelling Structural Colours of Euphaedra neophron (Lepidoptera: Nymphalidae) Finely Tuned by Wing Scale Lower Lamina in Various Subspecies" Insects 14, no. 3: 303. https://doi.org/10.3390/insects14030303