
Effect of the Landscape on Insect Pests and Associated Natural Enemies in Greenhouses Crops: The Strawberry Study Case
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
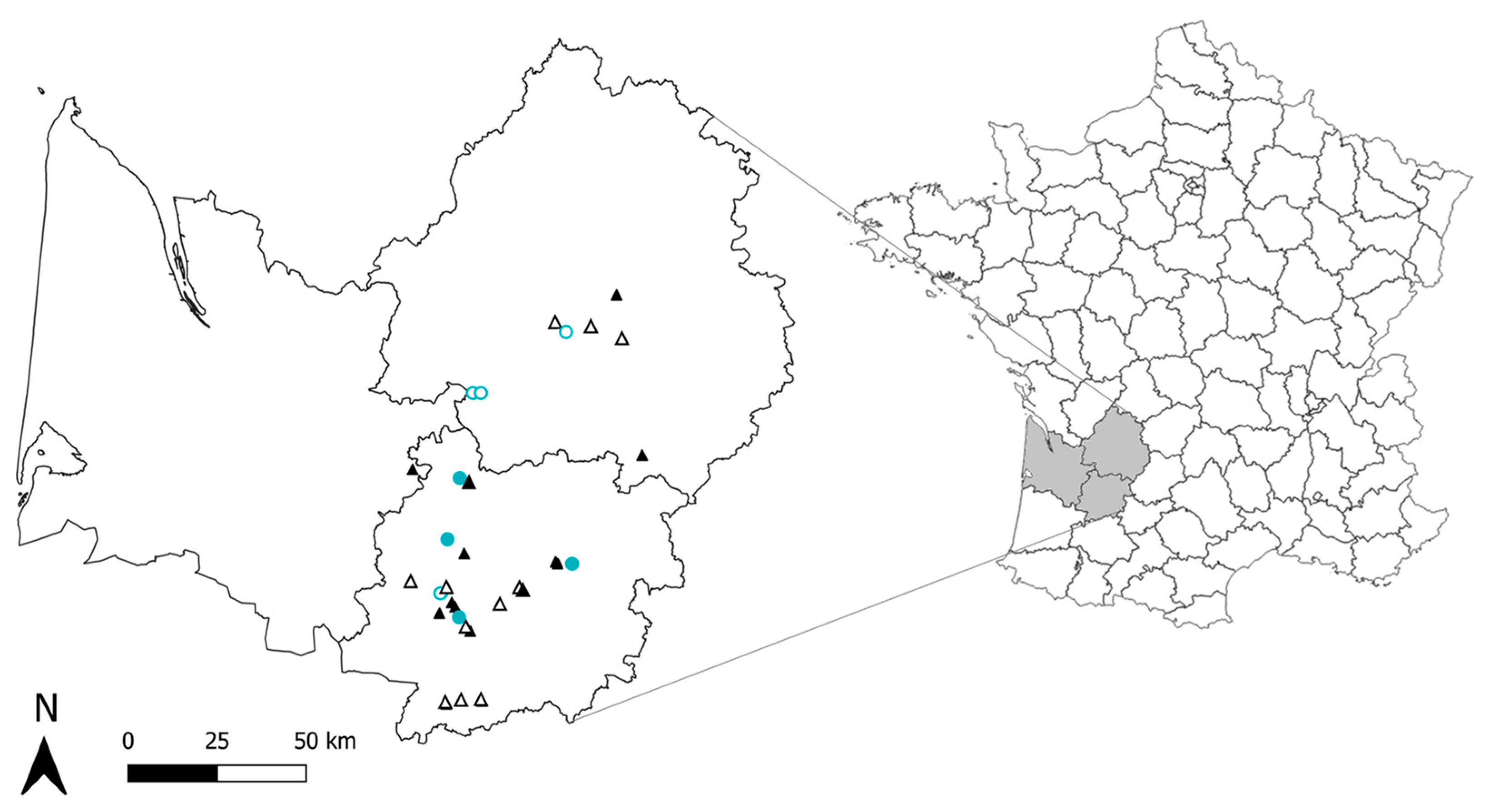
2.1. Monitored Greenhouses
2.2. Insect Sampling
2.3. Landscape Description
2.4. Data Analysis
3. Results
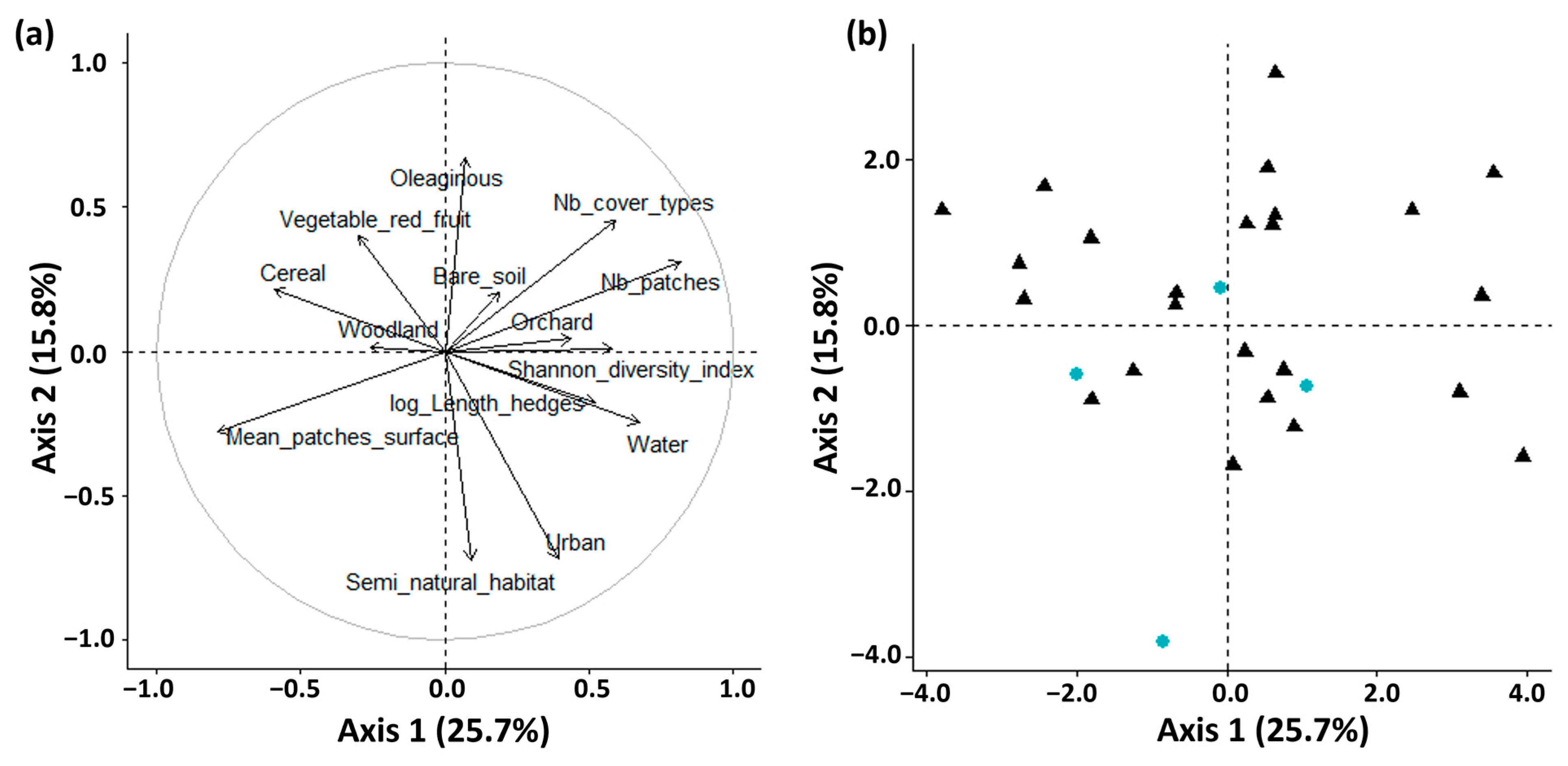
3.1. Variation in the Landscape Surrounding the Monitored Greenhouses
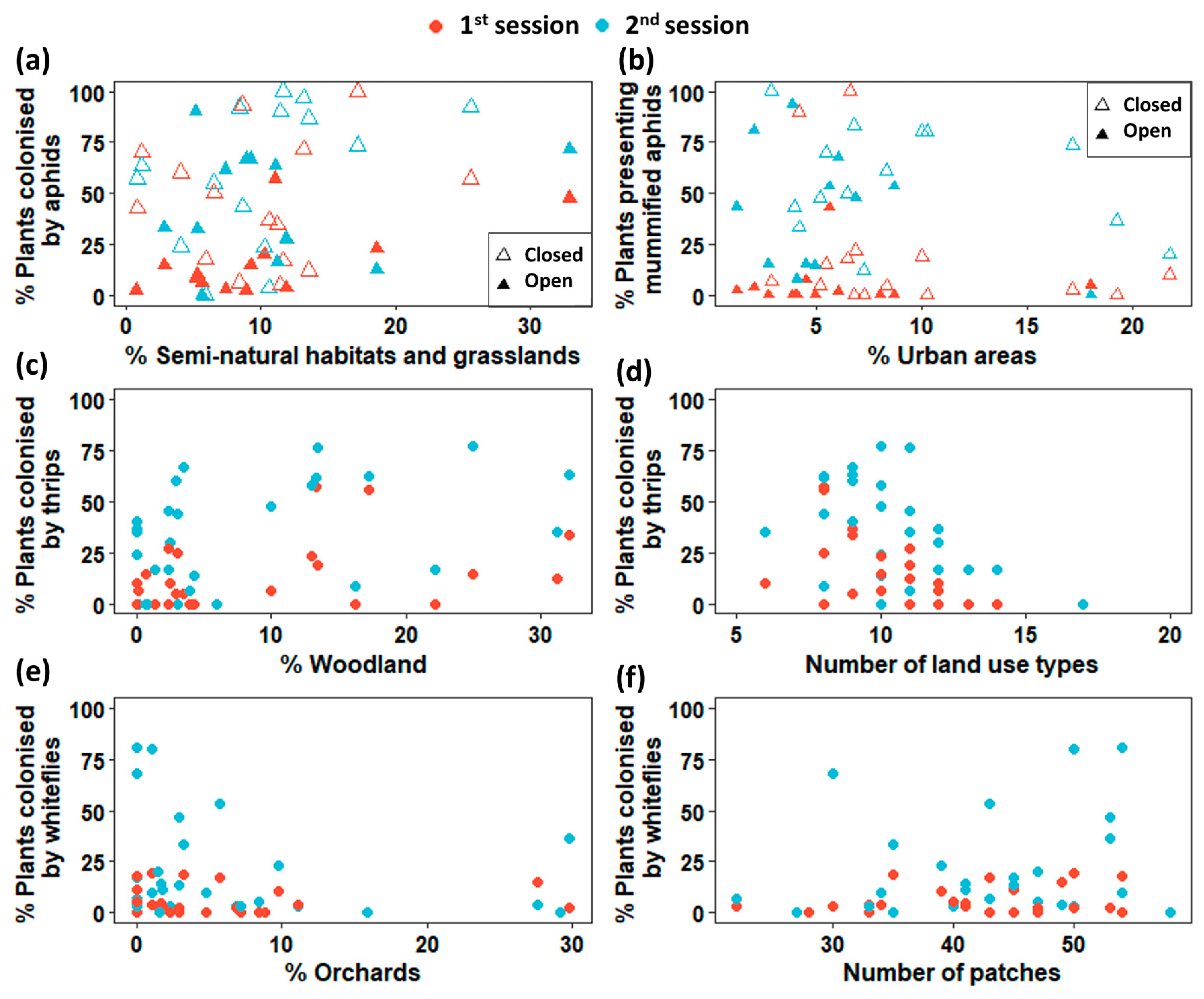
3.2. Landscape and Insect Occurrences
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forman, R.T.T. Some General Principles of Landscape and Regional Ecology. Landsc. Ecol. 1995, 10, 133–142. [Google Scholar] [CrossRef]
- Fahrig, L.; Baudry, J.; Brotons, L.; Burel, F.G.; Crist, T.O.; Fuller, R.J.; Sirami, C.; Siriwardena, G.M.; Martin, J.-L. Functional Landscape Heterogeneity and Animal Biodiversity in Agricultural Landscapes. Ecol. Lett. 2011, 14, 101–112. [Google Scholar] [CrossRef]
- Polis, G.A.; Power, M.E.; Huxel, G.R. (Eds.) Food Webs at the Landscape Level; University of Chicago Press: Chicago, IL, USA, 2004; ISBN 978-0-226-67327-1. [Google Scholar]
- Stoate, C.; Boatman, N.D.; Borralho, R.J.; Carvalho, C.R.; de Snoo, G.R.; Eden, P. Ecological Impacts of Arable Intensification in Europe. J. Environ. Manag. 2001, 63, 337–365. [Google Scholar] [CrossRef] [PubMed]
- Sala, O.E.; Stuart Chapin, F., III; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global Biodiversity Scenarios for the Year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland Biodiversity: Is Habitat Heterogeneity the Key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Rusch, A.; Chaplin-Kramer, R.; Gardiner, M.M.; Hawro, V.; Holland, J.; Landis, D.; Thies, C.; Tscharntke, T.; Weisser, W.W.; Winqvist, C.; et al. Agricultural Landscape Simplification Reduces Natural Pest Control: A Quantitative Synthesis. Agric. Ecosyst. Environ. 2016, 221, 198–204. [Google Scholar] [CrossRef] [Green Version]
- Le Provost, G.; Schenk, N.V.; Penone, C.; Thiele, J.; Westphal, C.; Allan, E.; Ayasse, M.; Blüthgen, N.; Boeddinghaus, R.S.; Boesing, A.L.; et al. The Supply of Multiple Ecosystem Services Requires Biodiversity across Spatial Scales. Nat. Ecol. Evol. 2023, 7, 236–249. [Google Scholar] [CrossRef]
- Gurr, G.M.; Wratten, S.D.; Barbosa, P. Success in Conservation Biological Control of Arthropods. In Biological Control: Measures of Success; Gurr, G., Wratten, S., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 105–132. ISBN 978-1-4020-1052-1. [Google Scholar]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat Management to Conserve Natural Enemies of Arthropod Pests in Agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Eilenberg, J.; Hajek, A.; Lomer, C. Suggestions for Unifying the Terminology in Biological Control. BioControl 2001, 46, 387–400. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Booij, C.J.H.; Tscharntke, T. Sustainable Pest Regulation in Agricultural Landscapes: A Review on Landscape Composition, Biodiversity and Natural Pest Control. Proc. R. Soc. B Biol. Sci. 2006, 273, 1715–1727. [Google Scholar] [CrossRef] [Green Version]
- Chaplin-Kramer, R.; O’Rourke, M.E.; Blitzer, E.J.; Kremen, C. A Meta-Analysis of Crop Pest and Natural Enemy Response to Landscape Complexity. Ecol. Lett. 2011, 14, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Veres, A.; Petit, S.; Conord, C.; Lavigne, C. Does Landscape Composition Affect Pest Abundance and Their Control by Natural Enemies? A Review. Agric. Ecosyst. Environ. 2013, 166, 110–117. [Google Scholar] [CrossRef]
- Haan, N.L.; Zhang, Y.; Landis, D.A. Predicting Landscape Configuration Effects on Agricultural Pest Suppression. Trends Ecol. Evol. 2020, 35, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tscharntke, T.; Karp, D.S.; Chaplin-Kramer, R.; Batáry, P.; DeClerck, F.; Gratton, C.; Hunt, L.; Ives, A.; Jonsson, M.; Larsen, A.; et al. When Natural Habitat Fails to Enhance Biological Pest Control—Five Hypotheses. Biol. Conserv. 2016, 204, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, L.M.; Otero, J.T.; Manzano, M.R. Biological Control of the Greenhouse Whitefly by Amitus Fuscipennis: Understanding the Role of Extrafloral Nectaries from Crop and Non-Crop Vegetation. Biol. Control 2013, 67, 227–234. [Google Scholar] [CrossRef]
- Pollier, A.; Tricault, Y.; Plantegenest, M.; Bischoff, A. Sowing of Margin Strips Rich in Floral Resources Improves Herbivore Control in Adjacent Crop Fields: Floral Resources Improve Herbivore Control. Agric. For. Entomol. 2019, 21, 119–129. [Google Scholar] [CrossRef]
- Karp, D.S.; Chaplin-Kramer, R.; Meehan, T.D.; Martin, E.A.; DeClerck, F.; Grab, H.; Gratton, C.; Hunt, L.; Larsen, A.E.; Martínez-Salinas, A.; et al. Crop Pests and Predators Exhibit Inconsistent Responses to Surrounding Landscape Composition. Proc. Natl. Acad. Sci. USA 2018, 115, E7863–E7870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messelink, G.J.; Lambion, J.; Janssen, A.; van Rijn, P.C.J. Biodiversity in and around Greenhouses: Benefits and Potential Risks for Pest Management. Insects 2021, 12, 933. [Google Scholar] [CrossRef]
- Postic, E.; Le Ralec, A.; Buchard, C.; Granado, C.; Outreman, Y. Variations in Community Assemblages and Trophic Networks of Aphids and Parasitoids in Protected Crops. Ecosphere 2020, 11, e03126. [Google Scholar] [CrossRef]
- Agreste, La Statistique Agricole. Available online: https://agreste.agriculture.gouv.fr/agreste-web/ (accessed on 14 February 2023).
- Bernardi, D.; Andreazza, F.; Botton, M.; Baronio, C.A.; Nava, D.E. Susceptibility and Interactions of Drosophila suzukii and Zaprionus indianus (Diptera: Drosophilidae) in Damaging Strawberry. Neotrop. Entomol. 2017, 46, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Jamieson, A.R.; Moreau, D.L.; Fillmore, S.A.E. Evaluation of Strawberry Genotypes for Chaetosiphon fragaefolii (Strawberry Aphid) Preference. Int. J. Fruit Sci. 2016, 16, 188–193. [Google Scholar] [CrossRef]
- Van Lenteren, J.C. A Greenhouse without Pesticides: Fact or Fantasy? Crop Prot. 2000, 19, 375–384. [Google Scholar] [CrossRef]
- Arnó, J.; Castañé, C.; Alomar, O.; Riudavets, J.; Agustí, N.; Gabarra, R.; Albajes, R. Forty Years of Biological Control in Mediterranean Tomato Greenhouses: The Story of Success. Isr. J. Entomol. 2018, 48, 209–226. [Google Scholar] [CrossRef]
- Turquet, M.; Geny, A.; Ginez, A.; Souriau, R.; Carmagnat, C. DEPHY Fraise: Réduire l’emploi des produits phytosanitaires en culture de fraisiers hors sol. Innov. Agron. 2019, 76, 3–16. [Google Scholar] [CrossRef]
- Postic, E.; Outreman, Y.; Derocles, S.; Granado, C.; Le Ralec, A. Genetics of Wild and Mass-Reared Populations of a Generalist Aphid Parasitoid and Improvement of Biological Control. PLoS ONE 2021, 16, e0249893. [Google Scholar] [CrossRef]
- Bienvenue Sur Le Projet QGIS! Available online: https://qgis.org/fr/site/ (accessed on 14 February 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.r-project.org/ (accessed on 3 October 2022).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Naimi, B.; Hamm, N.A.S.; Groen, T.A.; Skidmore, A.K.; Toxopeus, A.G. Where Is Positional Uncertainty a Problem for Species Distribution Modelling? Ecography 2014, 37, 191–203. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Statistics for Biology and Health; Springer: New York, NY, USA, 2009; ISBN 978-0-387-87457-9. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H. A General and Simple Method for Obtaining R2 from Generalized Linear Mixed-Effects Models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Rathee, M.; Singh, N.V.; Dalal, P.K. Integrated Pest Management under Protected Cultivation: A Review. J. Entomol. Zool. Stud. 2018, 6, 1201–1208. [Google Scholar]
- Alignier, A.; Raymond, L.; Deconchat, M.; Menozzi, P.; Monteil, C.; Sarthou, J.-P.; Vialatte, A.; Ouin, A. The Effect of Semi-Natural Habitats on Aphids and Their Natural Enemies across Spatial and Temporal Scales. Biol. Control 2014, 77, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Weisser, W.W.; Völkl, W. Dispersal in the Aphid Parasitoid, Lysiphlebus Cardui (Marshall) (Hym., Aphidiidae). J. Appl. Entomol. 1997, 121, 23–28. [Google Scholar] [CrossRef]
- Ratsimba, N.; Therond, O.; Parry, H.; Monteil, C.; Vialatte, A. Inconsistent Responses of Conservation Biocontrol to Landscape Structure: New Insights from a Network-Based Review. Ecol. Appl. 2022, 32, e2456. [Google Scholar] [CrossRef]
- Fok, E.J.; Petersen, J.D.; Nault, B.A. Relationships between Insect Predator Populations and Their Prey, Thrips Tabaci, in Onion Fields Grown in Large-Scale and Small-Scale Cropping Systems. BioControl 2014, 59, 739–748. [Google Scholar] [CrossRef] [Green Version]
- Picault, S. Protecting leek crops from Thrips tabaci: Using sanitizing plants to attract natural enemies. Infos-Ctifl 2017, 334, 26–39. [Google Scholar]
- Den Belder, E.; Elderson, J.; van den Brink, W.J.; Schelling, G. Effect of Woodlots on Thrips Density in Leek Fields: A Landscape Analysis. Agric. Ecosyst. Environ. 2002, 91, 139–145. [Google Scholar] [CrossRef]
- Aviron, S.; Poggi, S.; Varennes, Y.-D.; Lefèvre, A. Local Landscape Heterogeneity Affects Crop Colonization by Natural Enemies of Pests in Protected Horticultural Cropping Systems. Agric. Ecosyst. Environ. 2016, 227, 1–10. [Google Scholar] [CrossRef]
- Ardanuy, A.; Figueras, M.; Matas, M.; Arnó, J.; Agustí, N.; Alomar, Ò.; Albajes, R.; Gabarra, R. Banker Plants and Landscape Composition Influence Colonisation Precocity of Tomato Greenhouses by Mirid Predators. J. Pest Sci. 2022, 95, 447–459. [Google Scholar] [CrossRef]
- Baguette, M.; Blanchet, S.; Legrand, D.; Stevens, V.M.; Turlure, C. Individual Dispersal, Landscape Connectivity and Ecological Networks. Biol. Rev. 2013, 88, 310–326. [Google Scholar] [CrossRef]
- May, R.M.; Hassell, M.P.; Anderson, R.M.; Tonkyn, D.W. Density Dependence in Host-Parasitoid Models. J. Anim. Ecol. 1981, 50, 855–865. [Google Scholar] [CrossRef]
- Gabarra, R.; Alomar, Ò.; Castañé, C.; Goula, M.; Albajes, R. Movement of Greenhouse Whitefly and Its Predators between in- and outside of Mediterranean Greenhouses. Agric. Ecosyst. Environ. 2004, 102, 341–348. [Google Scholar] [CrossRef]
- Garratt, M.P.D.; Wright, D.J.; Leather, S.R. The Effects of Farming System and Fertilisers on Pests and Natural Enemies: A Synthesis of Current Research. Agric. Ecosyst. Environ. 2011, 141, 261–270. [Google Scholar] [CrossRef]
- Bommarco, R.; Miranda, F.; Bylund, H.; Björkman, C. Insecticides Suppress Natural Enemies and Increase Pest Damage in Cabbage. J. Econ. Entomol. 2011, 104, 782–791. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Landscape Properties | Landscape Metric | Metric Components | Maintained after VIF 1 Analysis |
|---|---|---|---|
| Landscape composition | % Cereal crops | Corn, Wheat, Oats, Barley | NO |
| % Oleaginous crops | Sunflower, Rapeseed | yes | |
| % Vegetable and red fruit crops | Potato, Beetroot, Sweet Pepper/Chili, Tomato, Eggplant, Raspberry, Blueberry, Red fruit, Zucchini, Salad, Strawberry | yes | |
| % Orchards | Walnut, Hazelnut, Chestnut, Orchard, Kiwi, Vine, Forestry, Tree nurseries | yes | |
| % Semi-natural habitats and grassland | Grassland, Weedy area, Wasteland, Flower strips | yes | |
| % Woodland | Wood | yes | |
| % Water | Rivers, Reservoirs, Ponds, Basin | yes | |
| % Bare ground | yes | ||
| % Urban | Residential areas, Urban environment, Industrial zones | yes | |
| Landscape heterogeneity | Shannon diversity index | H’= −Σ pi ln pi; pi = the percentage of the land cover category i | yes |
| Number of land cover types | yes | ||
| Landscape connectivity | log (Length of hedges) | yes | |
| Landscape fragmentation | Number of patches | yes | |
| Mean patch surface | NO |
| Sampling Session | Aphids | Phytophagous Bugs | Thrips | Whiteflies | Aphid Parasitoids | Aphid Predators |
|---|---|---|---|---|---|---|
| 1st Session | 0.317 ± 0.054 | 0.014 ± 0.005 | 0.129 ± 0.031 | 0.052 ± 0.012 | 0.021 ± 0.009 | 0.127 ± 0.048 |
| 2nd Session | 0.516 ± 0.062 | 0.049 ± 0.021 | 0.337 ± 0.048 | 0.198 ± 0.046 | 0.085 ± 0.019 | 0.494 ± 0.058 |
| Response | Significant Explanatory Variables | β | se (β) | p-Value of β | Conditional R2 | Marginal R2 |
|---|---|---|---|---|---|---|
| % Plants infested with aphids | % Semi-natural habitat and grassland | 0.061 | 0.020 | 0.003 | 0.379 | 0.258 |
| Session 1st | 0 | 0.000 | ||||
| Session 2nd | 1.638 | 0.126 | <0.001 | |||
| Closed greenhouse | 0 | 0.000 | ||||
| Open greenhouse | −1.158 | 0.318 | <0.001 | |||
| No release of aphid predators | 0 | 0.000 | ||||
| Release of aphid predators | 0.950 | 0.381 | 0.012 | |||
| % Plants infested with phytophagous bugs | Session 1st | 0 | 0.000 | 0.637 | 0.067 | |
| Session 2nd | 1.555 | 0.388 | <0.001 | |||
| % Plants infested with thrips | % Wooded area | 0.055 | 0.024 | 0.030 | 0.503 | 0.293 |
| Number of land use types | −0.421 | 0.121 | 0.001 | |||
| Session 1st | 0 | 0.000 | ||||
| Session 2nd | 1.503 | 0.150 | <0.001 | |||
| % Plants infested with whiteflies | Number of patches | 0.081 | 0.001 | 0.020 | 0.499 | 0.223 |
| % Orchards | −0.078 | 0.001 | 0.028 | |||
| Session 1st | 0 | 0.000 | ||||
| Session 2nd | 1.712 | 0.001 | <0.001 | |||
| % Plants infested with aphid predators | Session 1st | 0 | 0.000 | 0.282 | 0.136 | |
| Session 2nd | 1.582 | 0.001 | <0.001 | |||
| % Plants presenting mummified aphids | % Urban area | −0.130 | 0.061 | 0.037 | 0.648 | 0.377 |
| Session 1st | 0 | 0.000 | ||||
| Session 2nd | 3.044 | 0.188 | <0.001 | |||
| Closed greenhouse | 0 | 0.000 | ||||
| Open greenhouse | −2.325 | 0.631 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doehler, M.; Chauvin, D.; Le Ralec, A.; Vanespen, É.; Outreman, Y. Effect of the Landscape on Insect Pests and Associated Natural Enemies in Greenhouses Crops: The Strawberry Study Case. Insects 2023, 14, 302. https://doi.org/10.3390/insects14030302
Doehler M, Chauvin D, Le Ralec A, Vanespen É, Outreman Y. Effect of the Landscape on Insect Pests and Associated Natural Enemies in Greenhouses Crops: The Strawberry Study Case. Insects. 2023; 14(3):302. https://doi.org/10.3390/insects14030302
Chicago/Turabian StyleDoehler, Marianne, Delphine Chauvin, Anne Le Ralec, Émeline Vanespen, and Yannick Outreman. 2023. "Effect of the Landscape on Insect Pests and Associated Natural Enemies in Greenhouses Crops: The Strawberry Study Case" Insects 14, no. 3: 302. https://doi.org/10.3390/insects14030302