
Observation of Genetic Gain with Instrumental Insemination of Honeybee Queens
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
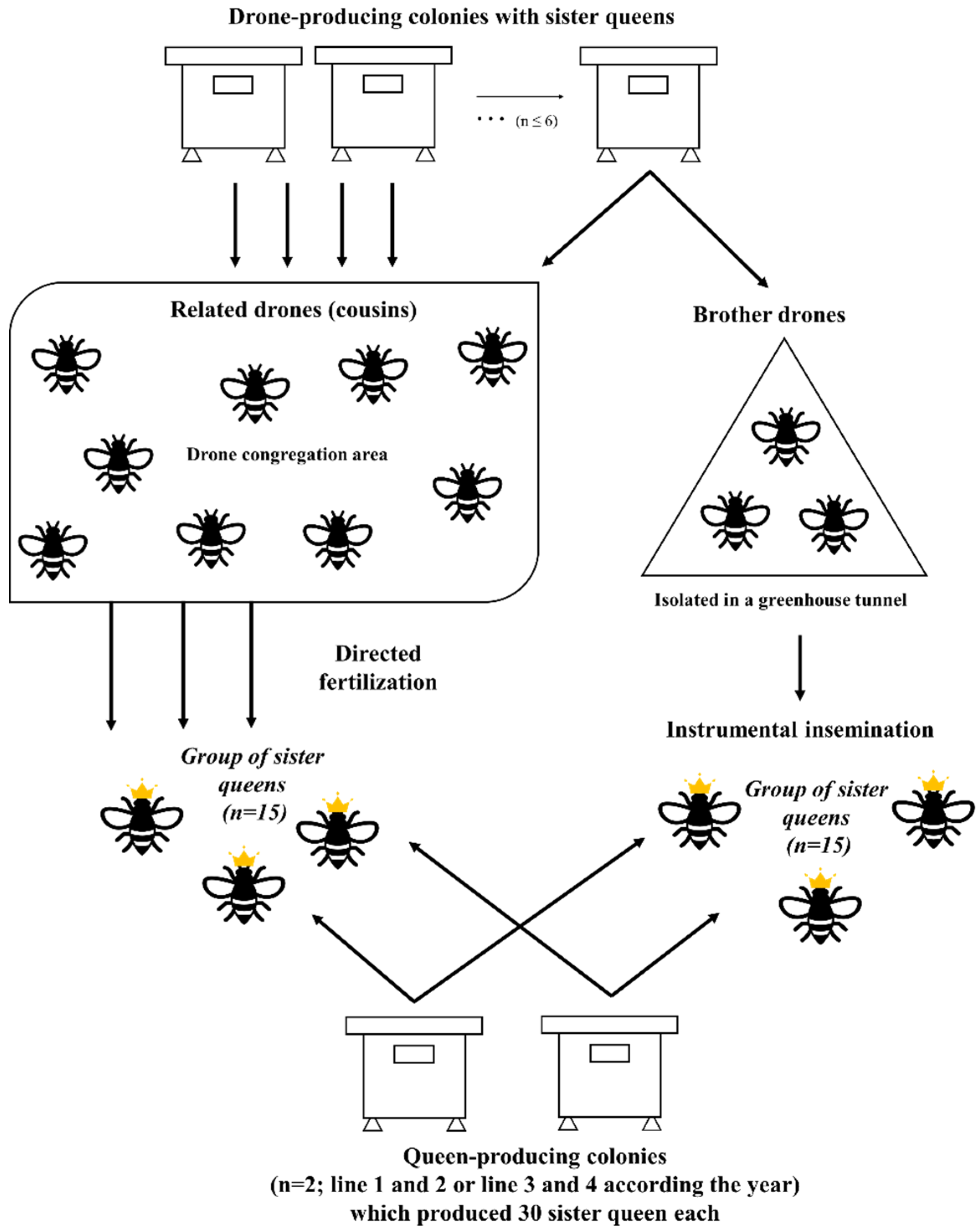
2. Materials and Methods
2.1. Honeybee Colonies
2.2. The Choice of Colonies for Breeding
2.3. Instrumental Insemination
2.3.1. Semen Collection
2.3.2. Insemination of Virgin Queen
2.3.3. Natural Mating
2.3.4. Colony Management
2.3.5. Colony Performance Criteria
- Honey production: honey supers of each colony were weighed before and after summer on a platform scale (CAS-131 USA, East-Rutherford, NY, USA; CAS CI-2001BS) to calculate the colony’s total seasonal honey production.
2.3.6. Statistical Analysis
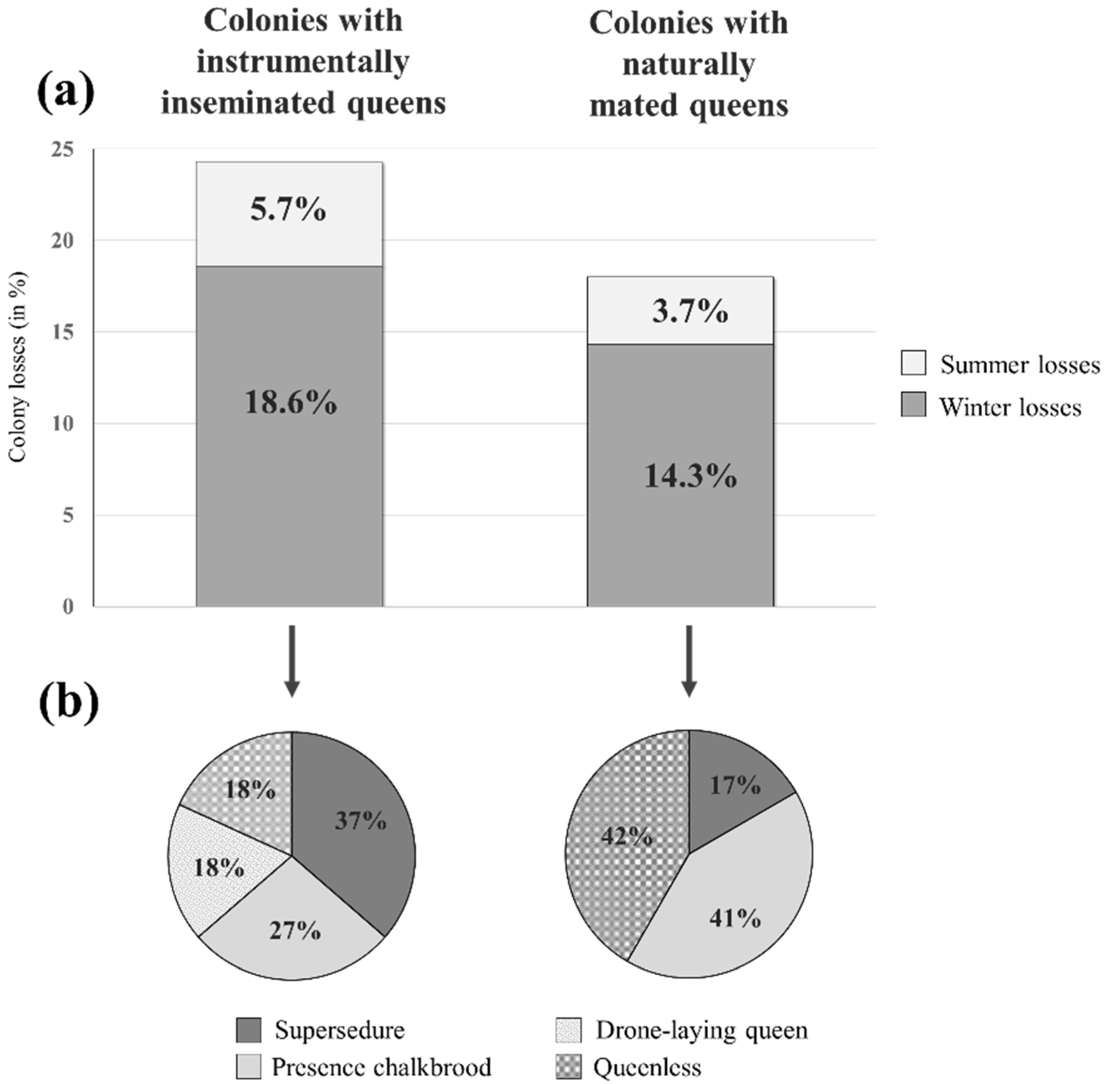
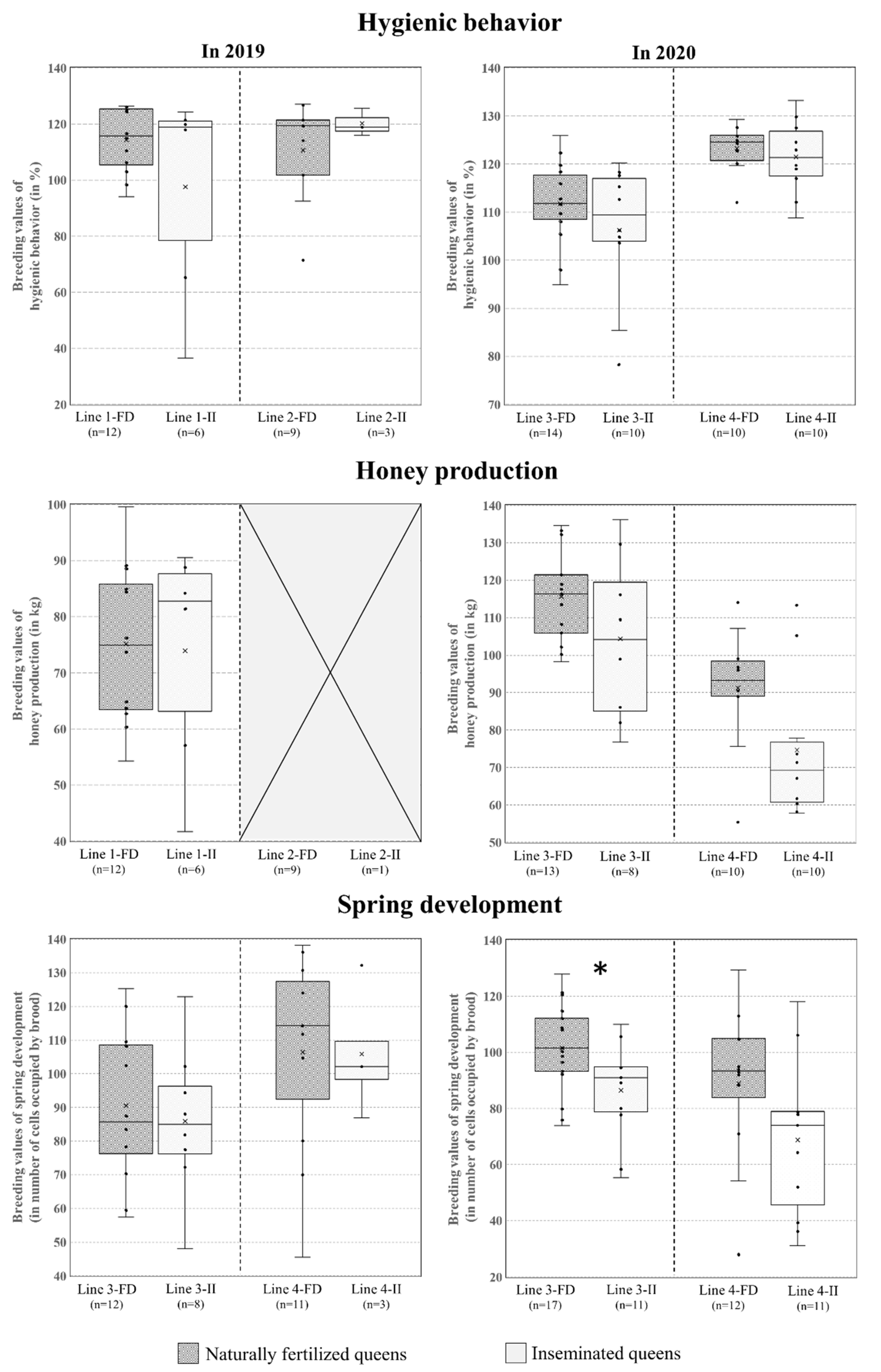
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cobey, S.W.; Sheppard, W.S.; Tarpy, D.R. Chapter 4: Status of Breeding Practices and Genetic Diversity in Domestic U.S. Honey Bees. In Honey Bee Colony Health: Challenges and Sustainable Solutions; CRC Press: Boca Raton, FL, USA, 2009; pp. 39–53. [Google Scholar]
- Bixby, M.; Mcafee, A. Queens Rule! Growing Canada’s Queen Breeding Industry: Results from the 2016-2017 Queen Breeder Survey. Am. Bee J. 2017, 157, 893–895. [Google Scholar]
- Petersen, G.E.L.; Fennessy, P.F.; Amer, P.R.; Dearden, P.K. Designing and implementing a genetic improvement program in commercial beekeeping operations. J. Apic. Res. 2020, 59, 638–647. [Google Scholar] [CrossRef]
- Bienefeld, K.; Ehrhardt, K.; Reinhardt, F. Genetic Evaluation in the Honey Bee Considering Queen and Worker Effects—A BLUP-Animal Model Approach. Apidologie 2007, 38, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Minvielle, F. Principes D’amélioration Génétique Des Animaux Domestiques, 1st ed.; Les presses de l’Université Laval: Québec, QC, Canada, 1990; pp. 15–68. [Google Scholar]
- Kerr, R.J.; Hammond, K.; Kinghorn, B.P. Effects of multiple-sire matings on genetic evaluations, selection response and rates of inbreeding. Livest. Prod. Sci. 1994, 38, 161–168. [Google Scholar] [CrossRef]
- Bienefeld, K.; Ehrhardt, K.; Reinhardt, F. Bee Breeding around the World—Noticeable Success in Honey Bee Selection after the Introduction of Genetic Evaluation Using BLUP. Am. Bee J. 2008, 148, 739–742. [Google Scholar]
- Bienefeld, K. Breeding Success or Genetic Diversity in Honey Bees? Bee World 2016, 93, 40–44. [Google Scholar] [CrossRef]
- Hoppe, A.; Du, M.; Bernstein, R.; Tiesler, F.K.; Kärcher, M.; Bienefeld, K. Substantial genetic progress in the international Apis mellifera carnica population since the implementation of genetic evaluation. Insects 2020, 11, 768. [Google Scholar] [CrossRef]
- Maucourt, S.; Fortin, F.; Robert, C.; Giovenazzo, P. Genetic progress achieved during 10 years of selective breeding for honeybee traits of interest to the beekeeping industry. Agriculture 2021, 11, 535. [Google Scholar] [CrossRef]
- Willam, A. Parameter- und Zuchtwertschätzung für Die Honigbiene (Apis mellifera carnica). Ph.D. Thesis, University of Agricultural Sciences, Vienna, Austria, 1991. [Google Scholar]
- Tarpy, D.R.; Nielsen, D.I. Structure of Honey Bee Colonies (Hymenoptera: Apidae). Ann. Entomol. Soc. Am. 2002, 95, 513–528. [Google Scholar] [CrossRef] [Green Version]
- Oxley, P.R.; Oldroyd, B.P. The genetic architecture of honeybee breeding. In Advances in Insect Physiology; Academic Press: Burlington, VT, USA, 2010; pp. 83–118. [Google Scholar]
- Plate, M.; Bernstein, R.; Hoppe, A.; Bienefeld, K. The importance of controlled mating in honeybee breeding. Genet. Sel. Evol. 2019, 51, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Büchler, R.; Andonov, S.; Bienefeld, K.; Costa, C.; Kezic, N.; Kryger, P.; Spivak, M.; Uzunov, A.; Wilde, J.; Hatjina, F. Standard methods for rearing and selection of Apis mellifera Queens. J. Apic. Res. 2013, 52, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Scharpenberg, H.; Neumann, P.; van Praagh, J.P.; Moritz, R.F.A. Reliability of an island mating apiary under routine management. J. Apic. Res. 2006, 45, 153–154. [Google Scholar] [CrossRef]
- Guzmán-Novoa, E. Elemental Genetics and Breeding for the Honeybee; Ontario Beekeepers Association: Ontario, ON, Canada, 2007. [Google Scholar]
- Fert, G. L’élevage Des Reines; Editions Rustica: Paris, France, 2014. [Google Scholar]
- Hellmich, R.L.; Waller, G.D. Preparing for africanized honey bees: Evaluating control in mating apiaries. Am. Bee J. 1990, 130, 537–542. [Google Scholar]
- Jensen, A.; Palmer, K.; Chaline, N.; Raine, N.; Tofilski, A.; Martin, S.; Pedersen, B.; Boomsma, J.; Ratnieks, F. Quantifying honey bee mating range and isolation in semi-isolated valleys by DNA microsatellite paternity analysis. Conserv. Genet. 2005, 6, 527–537. [Google Scholar] [CrossRef]
- Chapleau, J.-P. Apiculture: La Sélection Chez Les Abeilles; MAPAQ: Québec, QC, Canada, 1988. [Google Scholar]
- Cobey, S.W. Comparison studies of instrumentally inseminated and naturally mated honey bee queens and factors affecting their performance. Apidologie 2007, 38, 390–410. [Google Scholar] [CrossRef] [Green Version]
- Cobey, S.W.; Tarpy, D.R.; Woyke, J. Standard methods for instrumental insemination of Apis mellifera queens. J. Apic. Res. 2013, 52, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Currie, R.W.; Pernal, S.F.; Guzmán-Novoa, E. Honey Bee Colony Losses in Canada. J. Apic. Res 2010, 49, 104–106. [Google Scholar] [CrossRef]
- Statement on Honey bee Wintering Losses in Canada 2019—Canadian Association of Professional Apiculturists. Available online: https://capabees.com/shared/2018-2019-CAPA-Statement-on-Colony-Losses.pdf (accessed on 13 September 2021).
- Kevan, P.; Guzmán, E.; Skinner, A.; van Englesdorp, D. Colony Collapse Disorder in Canada: Do we have a problem? HiveLights 2007, 20, 14–16. [Google Scholar]
- Statement Honey Bee Wintering Losses in Canada (2020)—Canadian Association of Professional Apiculturists. Available online: https://capabees.com/shared/CAPA-Statement-on-Colony-Losses-2020.pdf (accessed on 29 April 2021).
- Bixby, M. Canadian Beekeeping Perspective on Colony Health and Growing Our Local Queen Supply. HiveLights 2017, 30, 13–15. [Google Scholar]
- Seeley, T.D.; Tarpy, D.R. Queen promiscuity lowers disease within honeybee colonies. Proc. R. Soc. B 2007, 274, 67–72. [Google Scholar] [CrossRef]
- Parker, R.; Melathopoulos, A.P.; White, R.; Pernal, S.F.; Guarna, M.; Foster, L. Ecological Adaptation of Diverse Honey Bee (Apis Mellifera) Populations. PLoS ONE 2010, 5, e11096. [Google Scholar] [CrossRef]
- Meixner, M.D.; Büchler, R.; Costa, C.; Roy, M.F.; Hatjina, F.; Kryger, P.; Uzunov, A.; Carreck, N.L. Honey Bee Genotypes and the Environment. J. Apic. Res. 2014, 53, 183–187. [Google Scholar] [CrossRef] [Green Version]
- Canadian Honey Council. Honey promotions through partnerships. HiveLights 2015, 28, 6–8. [Google Scholar]
- Bixby, M.; Polinsky, M.; Scarlett, R.; Higo, H.; Common, J.; Hoover, S.E.; Foster, L.J.; Zayed, A.; Cunningham, M.; Guarna, M.M. Impacts of COVID-19 on Canadian Beekeeping: Survey Results and a Profitability Analysis. J. Econ. Entomol. 2021, 114, 2245–2254. [Google Scholar] [CrossRef]
- Maucourt, S.; Fortin, F.; Robert, C.; Giovenazzo, P. Genetic parameters of honey bee colonies traits in a Canadian selection program. Insects 2020, 11, 587. [Google Scholar] [CrossRef] [PubMed]
- Connor, L.J. Bee Sex Essentials; Wicwas Press: Michigan, MI, USA, 2008; pp. 15–26. [Google Scholar]
- Stoian, R.O.; Mălinaş, C.; Botha, M.; Petrescu-Mag, I.V. Technical, sanitary and environmental sequences to improve artificial insemination of honey bee, Apis mellifera. Part I. Experimental results. ABAH Bioflux 2018, 10, 122–132. [Google Scholar]
- Rousseau, A.; Giovenazzo, P. Optimizing Drone Fertility With Spring Nutritional Supplements to Honey Bee (Hymenoptera: Apidae) Colonies. J. Econ. Entomol. 2016, 109, 1009–1014. [Google Scholar] [CrossRef]
- Locke, S.; Peng, Y.-S. The effects of drone age, semen storage and contamination on semen quality in the honey bee (Apis mellifera). Physiol. Entomol. 1993, 18, 144–148. [Google Scholar] [CrossRef]
- Rousseau, A.; Fournier, V.; Giovenazzo, P. Apis mellifera (Hymenoptera: Apidae) drone sperm quality in relation to age, genetic line, and time of breeding. Can. Entomol. 2015, 147, 702–711. [Google Scholar] [CrossRef]
- Paillard, M.; Rousseau, A.; Giovenazzo, P.; Bailey, J.L. Preservation of domesticated honey bee (Hymenoptera: Apidae) drone semen. J. Econ. Entomol. 2017, 110, 1412–1418. [Google Scholar] [CrossRef]
- Mackensen, O. Effect of carbon dioxide on initial oviposition of artificially inseminated and virgin queen bees. J. Econ. Entomol. 1947, 40, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Fischer, F. External influences on the filling of the spermatheca with sperm. Apidologie 1990, 21, 359–360. [Google Scholar]
- Finding DCA’s. Available online: https://www.beeculture.com/finding-dcas/ (accessed on 29 April 2021).
- Spivak, M.; Downey, D.L. Field assays for hygienic behavior in honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 1998, 91, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Spivak, M.; Reuter, G.S. Honey bee hygienic behavior. Am. Bee J. 1998, 138, 283–286. [Google Scholar]
- Giovenazzo, P.; Dubreuil, P. Evaluation of Spring Organic Treatments against Varroa Destructor (Acari: Varroidae) in Honey Bee Apis Mellifera (Hymenoptera: Apidae) Colonies in Eastern Canada. Exp. Appl. Acarol. 2011, 55, 65–76. [Google Scholar] [CrossRef]
- Delaplane, K.S.; Van der Steen, J.; Guzmán-Novoa, E. Standard methods for estimating strength parameters of Apis mellifera colonies. J. Apic. Res. 2013, 52, 1–12. [Google Scholar] [CrossRef]
- Chevalet, C.; Cornuet, J.-M. Etude theorique sur la selection du caractère production de miel chez l’abeille I. Modèle génétique et statistique. Apidologie 1982, 13, 39–65. [Google Scholar] [CrossRef] [Green Version]
- Application of Linear Models in Animal Breeding. Available online: http://cgil.uoguelph.ca/pub/ (accessed on 20 October 2021).
- Ibrahim, A.; Reuter, G.S.; Spivak, M. Field trial of honey bee colonies bred for mechanisms of resistance against Varroa destructor. Apidologie 2007, 38, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Rinderer, T.E.; Harris, J.W.; Hunt, G.J.; de Guzmán, L.I. Breeding for resistance to Varroa destructor in North America. Apidologie 2010, 41, 409–424. [Google Scholar] [CrossRef] [Green Version]
- Richard, F.J.; Tarpy, D.R.; Grozinger, C.M. Effects of insemination quantity on honey bee queen physiology. PLoS ONE 2007, 2, e980. [Google Scholar] [CrossRef]
- Richard, F.J.; Schal, C.; Tarpy, D.R.; Grozinger, C.M. Effects of Instrumental Insemination and Insemination Quantity on Dufour’s Gland Chemical Profiles and Vitellogenin Expression in Honey Bee Queens (Apis mellifera). J. Chem. Ecol. 2011, 37, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Fresnaye, J. L’insémination artificielle des reines d’abeilles. Ann. De L’abeille 1966, 9, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Kouas, H.; Adjlane, N. Les méthodes de l’insémination artificielle des reines d’abeille. Maghrebian J. Pure Appl. Sci. 2021, 7, 50–62. [Google Scholar]
- Oldroyd, B.P.; Rinderer, T.E.; Harbo, J.R.; Buco, S.M. Effects of intracolonial genetic diversity on honey bee (Hymenoptera: Apidae) colony performance. Ann. Entomol. Soc. Am. 1992, 85, 335–343. [Google Scholar] [CrossRef]
- Mattila, H.R.; Seeley, T.D. Genetic diversity in honey bee colonies enhances productivity and fitness. Science 2007, 317, 362–364. [Google Scholar] [CrossRef] [Green Version]
- Tarpy, D.R.; Vanengelsdorp, D.; Pettis, J.S. Genetic diversity affects colony survivorship in commercial honey bee colonies. Naturwissenschaften 2013, 100, 723–728. [Google Scholar] [CrossRef]
- Gerula, D.; Węgrzynowicz, P.; Panasiuk, B.; Bieńkowska, M.; Skowronek, W. Performance of bee colonies headed by queens instrumentally inseminated with semen of drones who come from a single colony or many colonies. J. Apic. Sci. 2014, 58, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Woyke, J. Effect of sex allele homo-heterozygosity on honeybee colony populations and on their honey production. Unfavourable development conditions and restricted queens. J. Apic. Res. 1981, 20, 148–155. [Google Scholar] [CrossRef]
- Baer, B.; Schmid-Hempel, P. Experimental variation in polyandry affects parasite loads and fitness in a bumble-bee. Nature 1999, 397, 151–154. [Google Scholar] [CrossRef]
- Tarpy, D.R.; Page, R.E. Sex determination and the evolution of polyandry in honey bees (Apis mellifera). Behav. Ecol. Sociobiol. 2002, 52, 143–150. [Google Scholar] [CrossRef]
- Beye, M.; Hasselmann, M.; Fondrk, M.K.; Page, R.E.; Omholt, S.W. The Gene Csd Is the Primary Signal for Sexual Development in the Honeybee and Encodes an SR-Type Protein. Cell 2003, 114, 419–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarpy, D.R. Genetic diversity within honeybee colonies prevents severe infections and promotes colony growth. Proc. R. Soc. B 2003, 270, 99–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.C.; Myerscough, M.R.; Graham, S.; Oldroyd, B.P. Honey bee nest thermoregulation: Diversity promotes stability. Science 2004, 305, 402–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zayed, A. Bee genetics and conservation. Apidologie 2009, 40, 237–262. [Google Scholar] [CrossRef] [Green Version]
- Desai, S.D.; Currie, R.W. Genetic diversity within honey bee colonies affects pathogen load and relative virus levels in honey bees, Apis mellifera L. Behav. Ecol. Sociobiol. 2015, 69, 1527–1541. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Queen-Producing Colonies | Selection Index | Breeding Value of Hygienic Behavior | Breeding Value of Honey Production | Breeding Value of Spring Development | |
|---|---|---|---|---|---|
| 2018 | Mother colony for line 1 | 178.5 | 185.9 | 116.3 | 133.9 |
| Mother colony for line 2 | 130.2 | 156.5 | 87.8 | 81.9 | |
| 2019 | Mother colony for line 3 | 154.7 | 132.8 | 156.3 | 144.7 |
| Mother colony for line 4 | 153.4 | 132.8 | 163.0 | 131.5 | |
| Drone-Producing Colonies for Natural Mating | Selection Index of Mother Colony of Sister Colonies Producing Drones | Average Selection Index of Sister Colonies Producing Drones | Breeding Value of Hygienic Behavior | Breeding Value of Honey Production | Breeding Value of Spring Development | |
|---|---|---|---|---|---|---|
| 2018 | 6 sister colonies | 104.3 | 93.0 ± 5.0 | 93.8 ± 5.0 | 97.2 ± 4.6 | 95.3 ± 11.1 |
| 2019 | 8 sister colonies | 148.2 | 119.3 ± 4.0 | 123.4 ± 3.9 | 108.5 ± 5.2 | 100.1 ± 4.9 |
| Drone-Producing Colonies for Instrumental Insemination | Selection Index of Mother Colony of Drone-Producing Colony | Selection Index of Drone-Producing Colony | Breeding Value of Hygienic Behavior | Breeding Value of Honey Production | Breeding Value of Spring Development | |
|---|---|---|---|---|---|---|
| 2018 | 1 drone-producing colony | 104.3 | 106.4 | 116.6 | 94.8 | 88.6 |
| 2019 | 1 drone-producing colony | 148.2 | 136.0 | 127.8 | 138.1 | 125.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maucourt, S.; Rousseau, A.; Fortin, F.; Robert, C.; Giovenazzo, P. Observation of Genetic Gain with Instrumental Insemination of Honeybee Queens. Insects 2023, 14, 301. https://doi.org/10.3390/insects14030301
Maucourt S, Rousseau A, Fortin F, Robert C, Giovenazzo P. Observation of Genetic Gain with Instrumental Insemination of Honeybee Queens. Insects. 2023; 14(3):301. https://doi.org/10.3390/insects14030301
Chicago/Turabian StyleMaucourt, Ségolène, Andrée Rousseau, Frédéric Fortin, Claude Robert, and Pierre Giovenazzo. 2023. "Observation of Genetic Gain with Instrumental Insemination of Honeybee Queens" Insects 14, no. 3: 301. https://doi.org/10.3390/insects14030301