
The Structure of the Female Genital System of the Diving Beetle Scarodytes halensis (Fabricius, 1787) (Hydroporinae, Dytiscidae), and the Organization of the Spermatheca and the Spermathecal Gland Complex

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
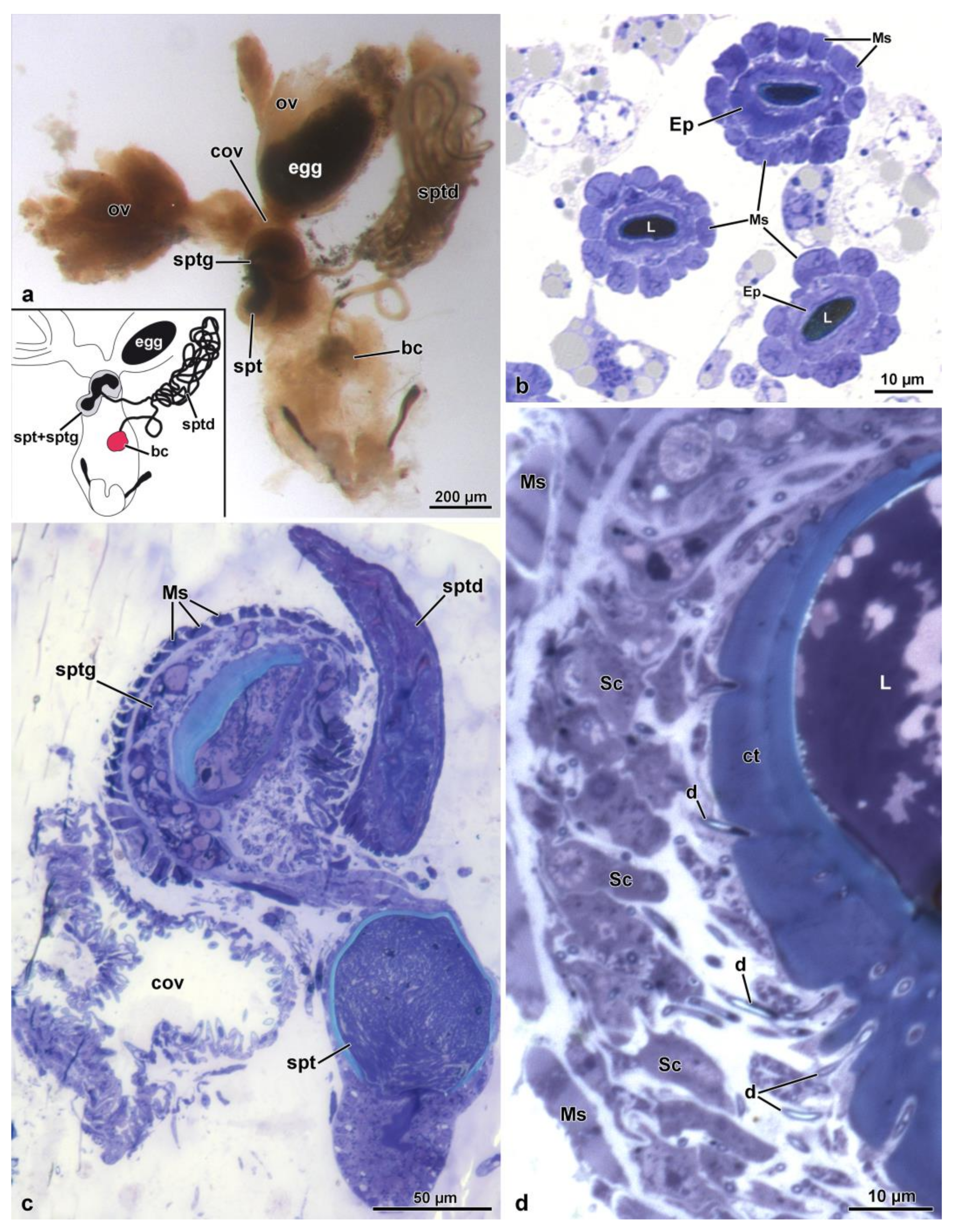
3.1. The General Organization of the Female Genital Apparatus
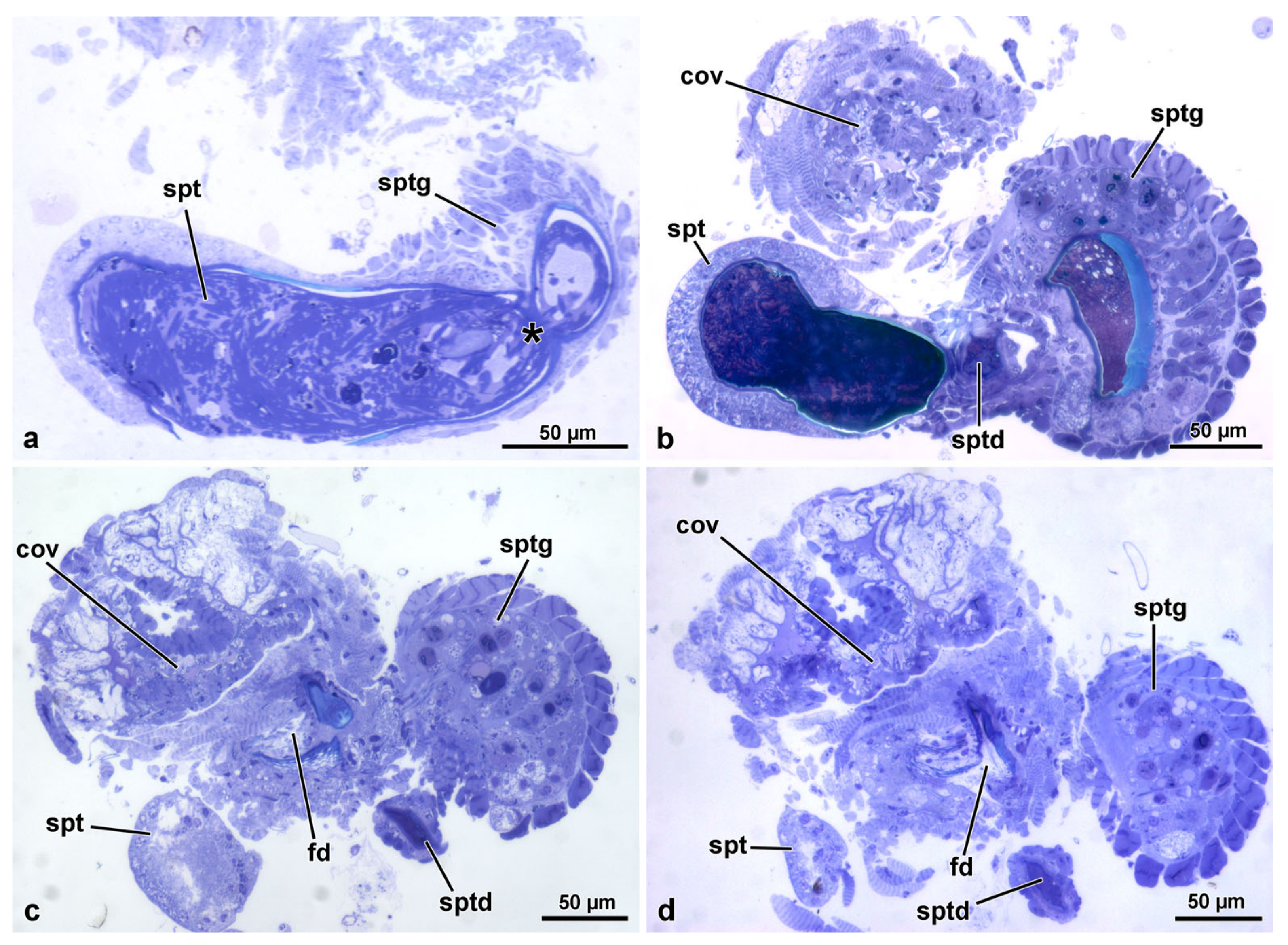
3.2. The Spermathecal Duct
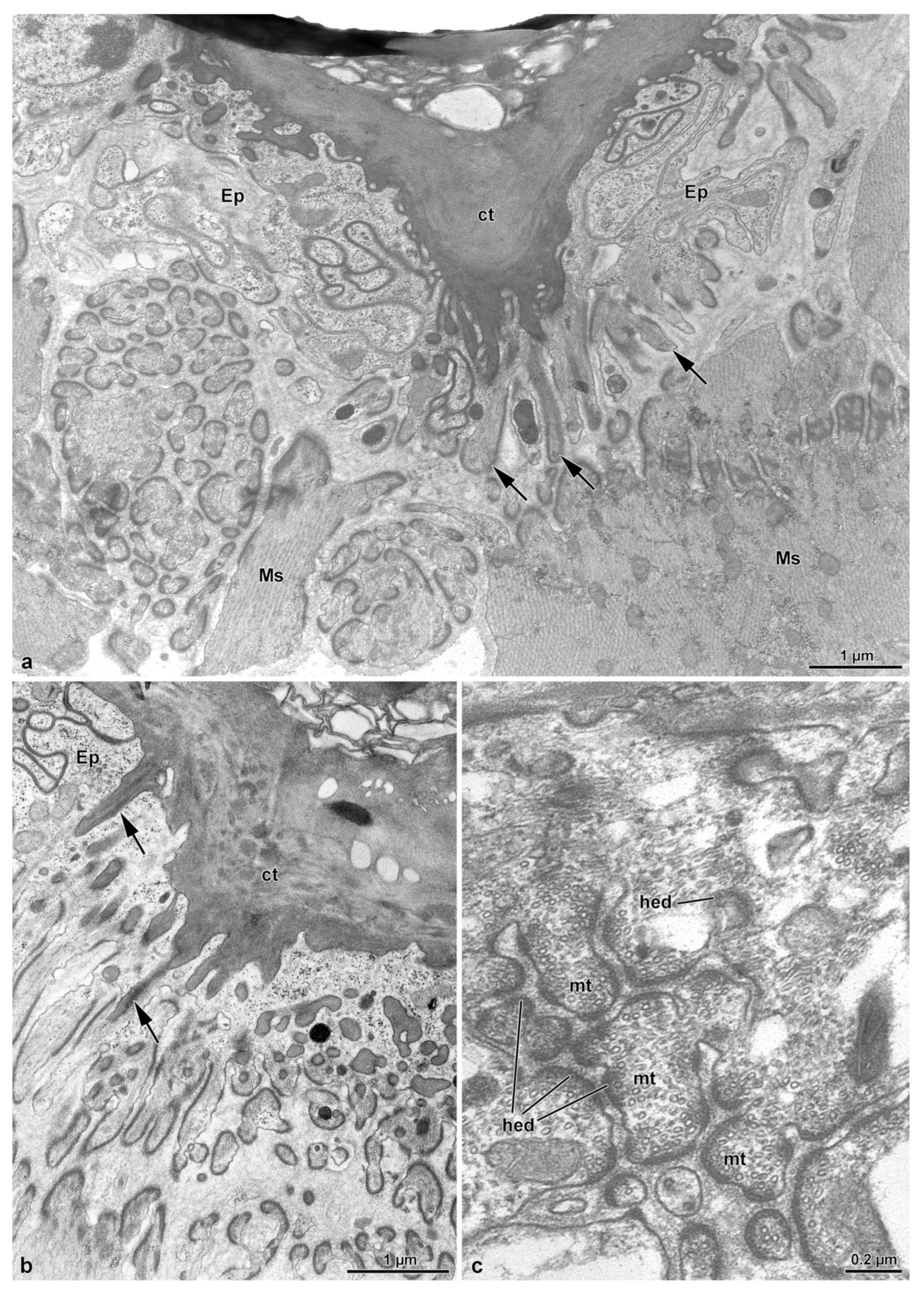
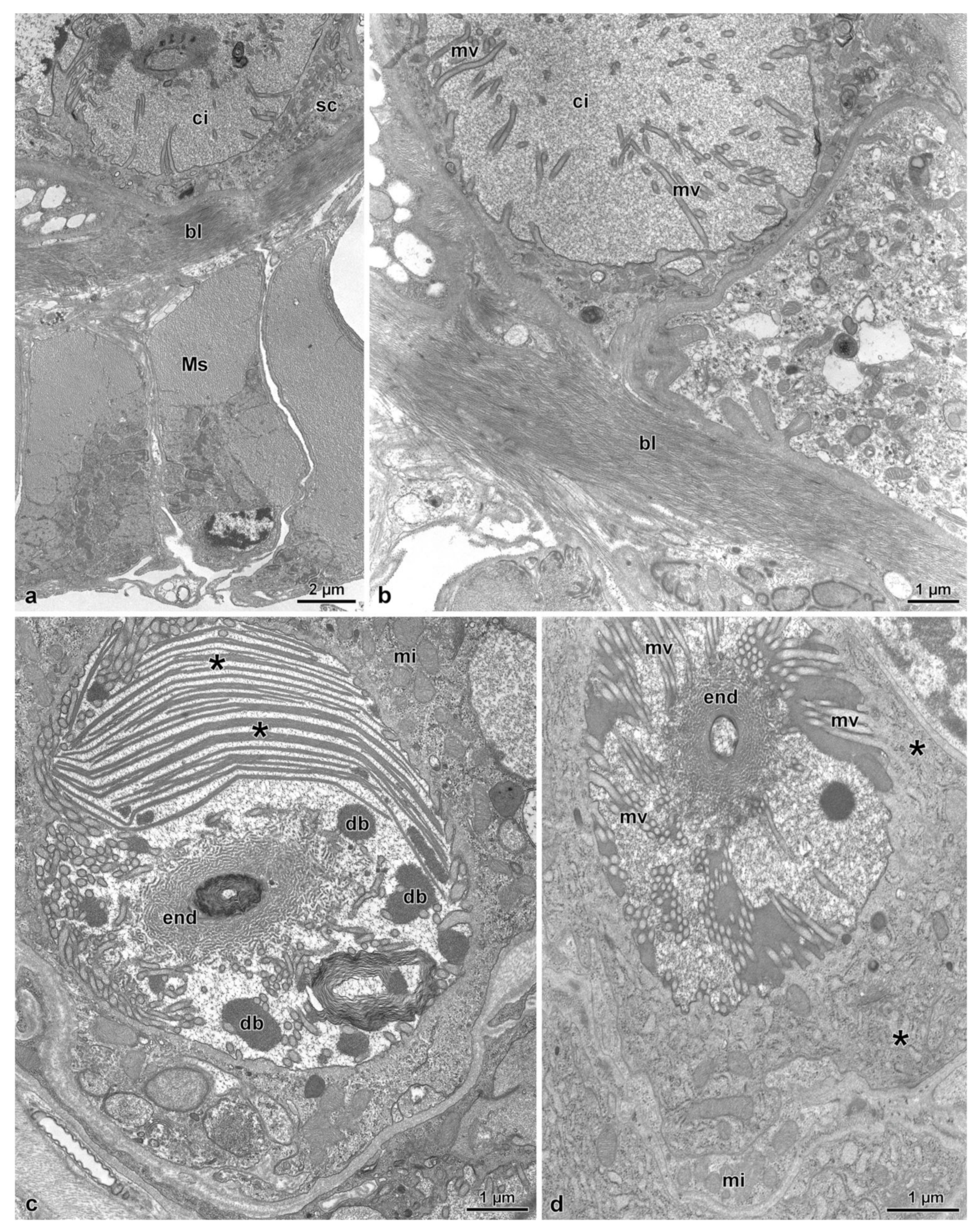
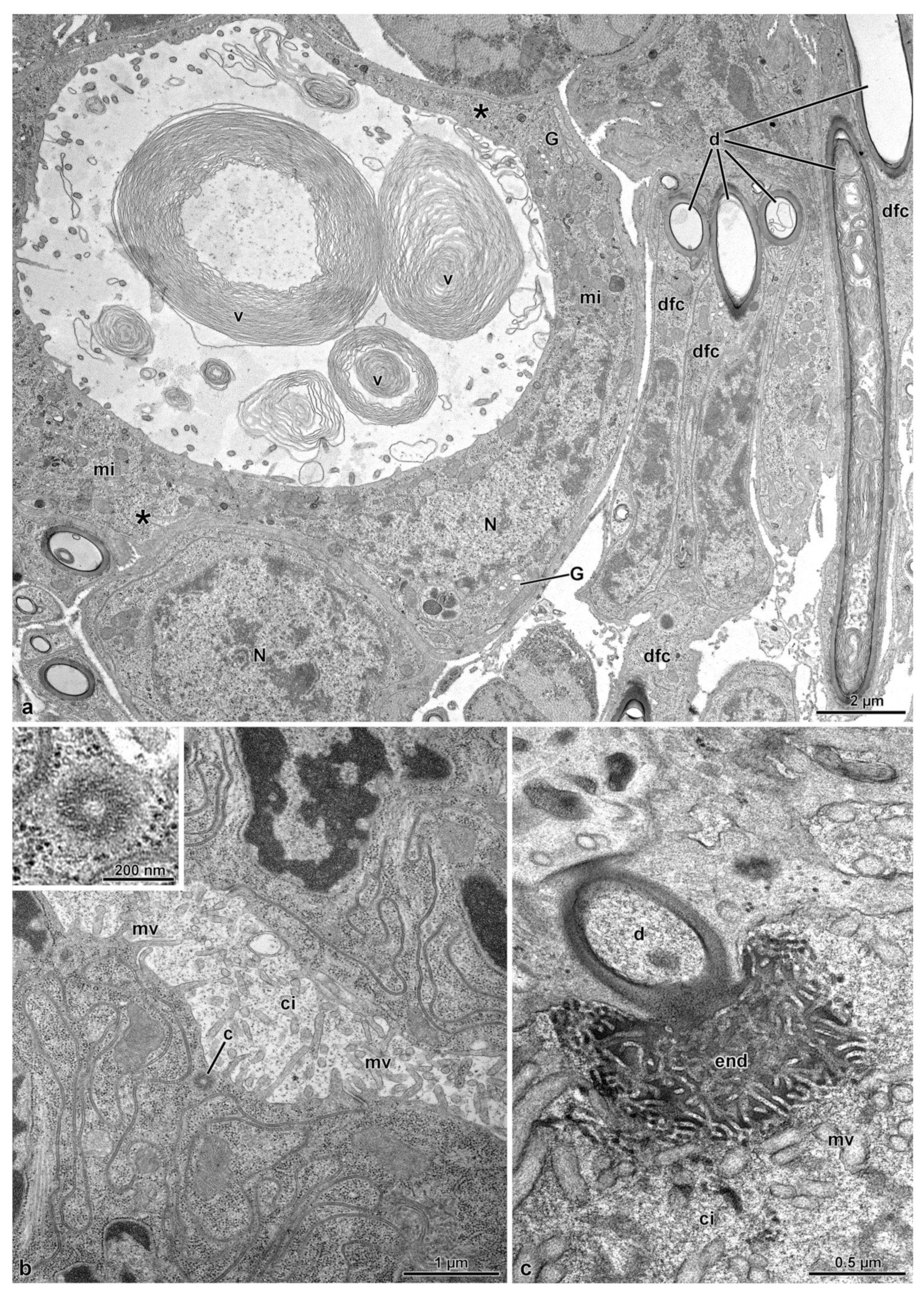
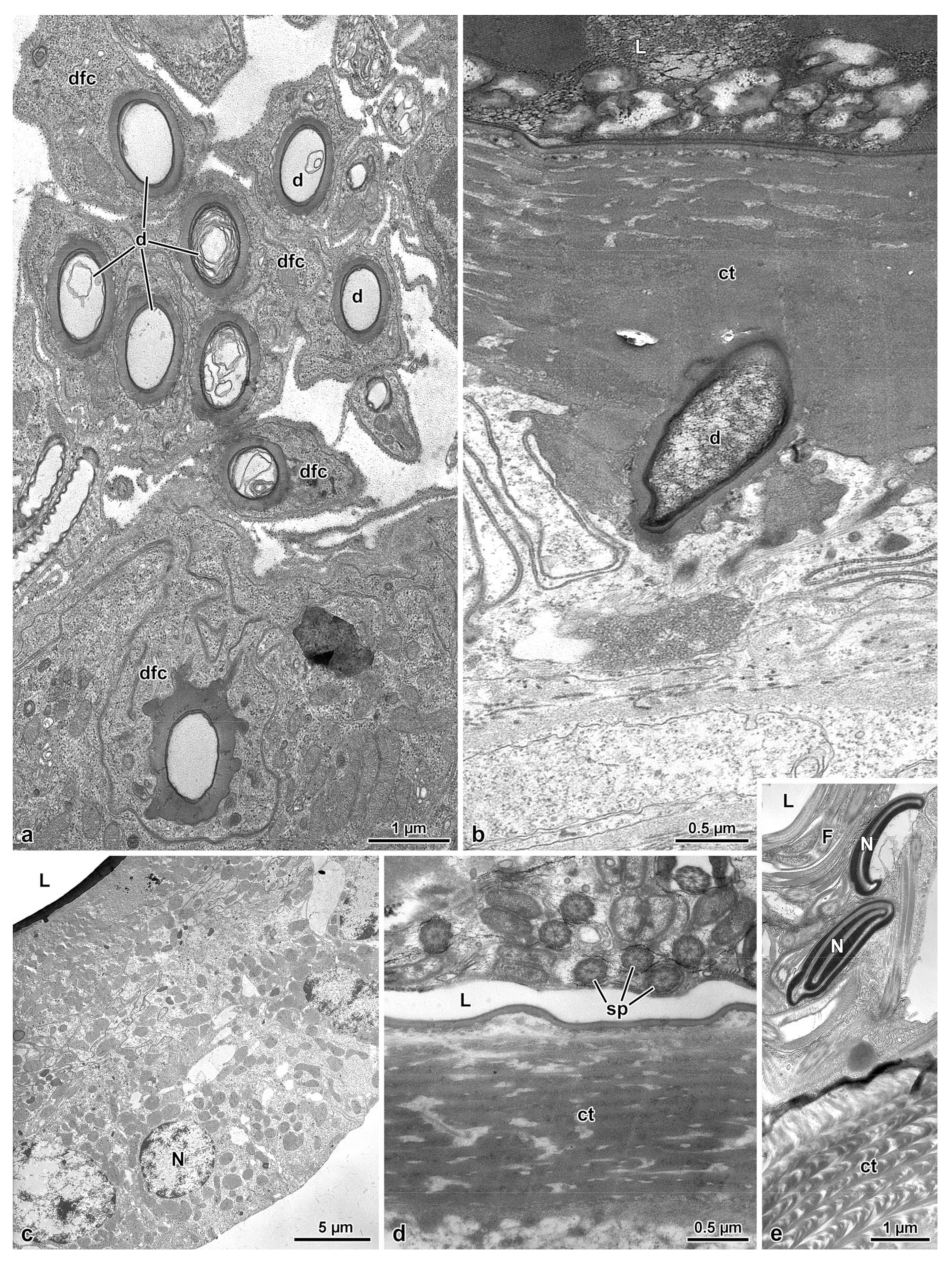
3.3. The Spermatheca and Spermathecal Gland Complex
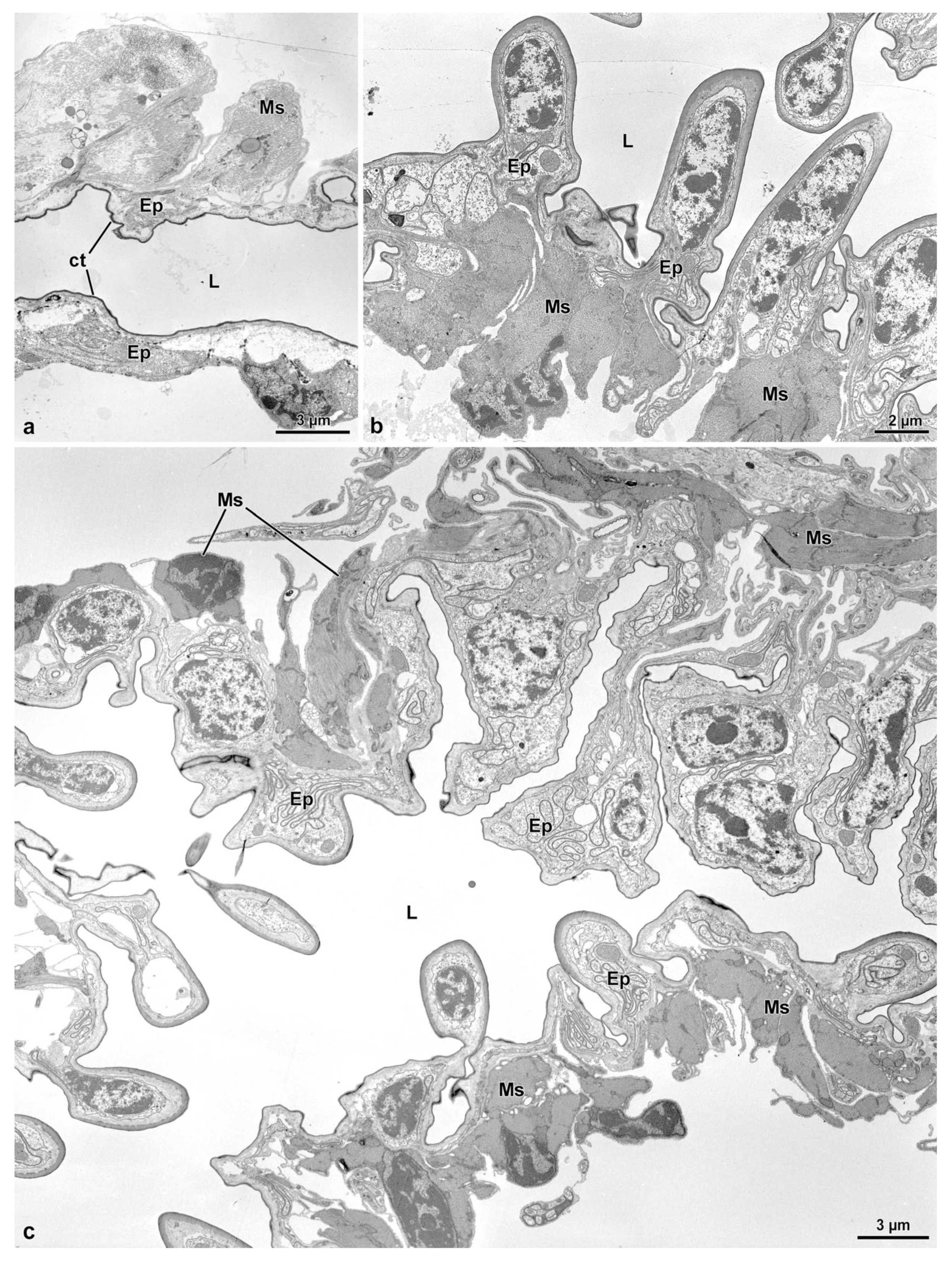
3.4. The Fertilization Duct and the Common Oviduct
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, G.T.; Pitnick, S. Sperm-female coevolution in Drosophila. Science 2002, 298, 1230–1233. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.T.; Pitnick, S. Functional significance of seminal receptacle length in Drosophila melanogaster. J. Evol. Biol. 2003, 16, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Pattarini, J.M.; Starmer, W.T.; Bjork, A.; Pitnick, S. Mechanisms underlying the sperm quality advantage in Drosophila melanogaster. Evolution 2006, 60, 2064–2080. [Google Scholar] [PubMed]
- Higginson, D.M.; Miller, K.B.; Segraves, K.A.; Pitnick, S. Convergence, recurrence and diversification of complex sperm traits in diving beetles (Dytiscidae). Evolution 2012, 66, 1650–1661. [Google Scholar] [CrossRef] [Green Version]
- Lüpold, S.; Manier, M.K.; Berben, K.S.; Smith, B.D.; Daley, S.H.; Buckley, S.H.; Belote, J.M.; Pitnick, S. How multivariate ejaculate traits determine competitive fertilization success in Drosophila melanogaster. Curr. Biol. 2012, 22, 1667–1672. [Google Scholar] [CrossRef] [Green Version]
- Lüpold, S.; Pitnick, S. Sperm form and function: What do we know about the role of sexual selection? Reproduction 2018, 155, R229–R243. [Google Scholar] [CrossRef] [Green Version]
- Dallai, R.; Gottardo, M.; Mercati, D.; Machida, R.; Mashimo, Y.; Matsumura, Y.; Beutel, R.G. Giant spermatozoa and a huge spermatheca: A case of coevolution of male and female reproductive organs in the ground louse Zorotypus impolitus (Insecta, Zoraptera). Arthropod Struct. Dev. 2014, 43, 135–151. [Google Scholar] [CrossRef]
- Dallai, R.; Fanciulli, P.P.; Lupetti, P.; Mercati, D. The ultrastructure of sperm and female sperm storage organs in the water strider Gerris lacustris L. (Heteroptera) and a possible example of genital coevolution. Arthropod Struct. Dev. 2021, 61, 101043. [Google Scholar] [CrossRef]
- Dallai, R.; Fanciulli, P.P.; Mercati, D.; Lupetti, P. Coevolution between female seminal receptacle and sperm morphology in the semiaquatic measurer bug Hydrometra stagnorum L. (Heteroptera, Hydrometridae). Arthropod Struct. Dev. 2021, 60, 101001. [Google Scholar] [CrossRef]
- Mercati, D.; Fanciulli, P.P.; Lupetti, P.; Dallai, R. The sperm structure of the diving beetles Stictonectes optatus (Seidlitz, 1887) and Scarodytes halensis (Fabricius, 1787) (Dytiscidae, Hydroporinae) with evidence of a spermatostyle in the sperm conjugation. Micron 2023, 166, 103412. [Google Scholar] [CrossRef]
- Dallai, R.; Mercati, D.; Fanciulli, P.P.; Lupetti, P. Fine structure of the female genital system of diving beetle Stictonectes optatus (Seidlitz, 1887) (Dytiscidae-Hydroporinae) and evidence of mating plug formation. Arthropod Struct. Dev. 2023, in press. [Google Scholar]
- Angus, R.B. Suphrodytes Des Gozis a valid genus, not a subgenus of Hydroporus Clairville (Coleoptera: Dytiscidae). Ent. Scand. 1985, 16, 269–275. [Google Scholar] [CrossRef]
- Deuve, T. Études Morphologiques et Phylogénétiques sur L’abdomen et les Genitalia Ectodermiques Femelles des Coleopteres Adephaga. Ph.D. Thesis, Université de Paris 6, Paris, France, 1988. [Google Scholar]
- Deuve, T. I’abdomen et les genitalia des femelles de Coleopteres Adephaga. Mem. Mus. Nat. Hist. Nat. 1993, 155, 184. [Google Scholar]
- Burmeister, E.G. Der Ovipositor der Hydradephaga (Coleoptera) und seine phylogenetische Bedeutung unter besonderer Berücksichtigung der Dytiscidae. Zoomorphologie 1976, 85, 165–257. [Google Scholar] [CrossRef]
- Burmeister, E.G. Funktionsmorphologie und Evolution des Ovipositor der Adephaga (Coleoptera). Verh. Naturwiss. Ver. Hambg. 1980, 24, 89–184. [Google Scholar]
- Burmeister, E.G. On the systematic position of Amphizoidae, emphasizing features of the female genital organs (Insecta: Coleoptera: Adephaga). Quaest. Entomol. 1990, 26, 245–272. [Google Scholar]
- Miller, K.B. On the phylogeny of the Dytiscidae (Insecta: Coleoptera) with emphasis on the morphology of the female reproductive system. Insect Syst. Evol. 2001, 32, 45–89. [Google Scholar] [CrossRef]
- De Marzo, L. Anatomical revision of the spermatheca in dytiscids. Entomologica 1997, 31, 207–219. [Google Scholar]
- Miller, K.B.; Bergsten, J. Predaceous diving beetle sexual system. In Ecology, Systematics, and Natural History of Predaceous Diving Beetles (Coleoptera: Dytiscidae); Yee, D.A., Ed.; Springer: New York, NY, USA, 2014; pp. 199–234. [Google Scholar]
- Quennedey, A. Insect epidermal gland cells: Ultrastructure and morphogenesis. In Microscopic Anatomy of Invertebrates; Harrison, F.W., Locke, M., Eds.; Wiley-Liss, Inc.: New York, NY, USA, 1998; Volume 11A, pp. 177–207. [Google Scholar]
- Sreng, L.; Quennedey, A. Role of a temporary ciliary structure in the morphogenesis of insect glands. An electron microscope study of the tergal glands of male Blattella germanica L. (Dictyoptera, Blattellidae). J. Ultrastruct. Res. 1976, 56, 78–95. [Google Scholar] [CrossRef] [PubMed]
- Quennedey, A. The moulting process of perennial class 3 gland cells during the postembryonic development of two heterometabolous insects: Blaberus (Dictyoptera) and Dysdercus (Heteroptera). Ann. Soc. Entomol. 1991, 27, 143–161. [Google Scholar]
- Lai-Fook, J.; Beaton, C. Muscle Insertions. In Microscopic Anatomy of Invertebrates; Harrison, F.W., Locke, M., Eds.; Wiley-Liss, Inc.: New York, NY, USA, 1998; Volume 11B, pp. 573–580. [Google Scholar]
- Noirot, C.; Noirot-Timothée, C. Cell Association. In Microscopic Anatomy of Invertebrates; Harrison, F.W., Locke, M., Eds.; Wiley-Liss, Inc.: New York, NY, USA, 1998; Volume 11A, pp. 27–49. [Google Scholar]
- Bitsch, C.; Bitsch, J. The endoskeletal structures in arthropods: Cytology, morphology and evolution. Arthropod Struct. Dev. 2002, 30, 159–177. [Google Scholar] [CrossRef]
- Joly, D.; Bressac, C. Sperm length in Drosophilidae (Diptera): Estimation by testis and receptacle lengths. Int. J. Insect Morph. Embryol. 1994, 23, 85–92. [Google Scholar] [CrossRef]
- Rice, W. Sexually antagonistic male adaptation triggered by experimental arrest of female evolution. Nature 1996, 381, 232–234. [Google Scholar] [CrossRef]
- Pitnick, S.; Markow, T.A.; Spicer, G.S. Evolution of multiple kinds of female sperm-storage organs in Drosophila. Evolution 1999, 53, 1804–1822. [Google Scholar]
- Pitnick, S.; Miller, G.T.; Reagan, J.; Holland, B. Males’ evolutionary responses to experimental removal of sexual selection. Proc. R. Soc. Lond. B. 2001, 268, 1071–1080. [Google Scholar] [CrossRef] [Green Version]
- Pitnick, S.; Hosken, D.J.; Birkhead, T.R. Sperm morphological diversity. In Sperm Biology, an Evolutionary Perspective; Birkhead, T.R., Hosken, D.J., Pitnick, S., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 69–149. [Google Scholar]
- Presgraves, D.C.; Baker, R.H.; Wilkinson, G.S. Coevolution of sperm and female reproductive tract morphology in stalk-eyed flies. Proc. R. Soc. Lond. B Biol. Sci. 1999, 266, 1041–1047. [Google Scholar] [CrossRef]
- Kotrba, M.; Heß, M.; Dallai, R. Giant spermatozoa of Diasemopsis (Diopsidae, Diptera)—Structural, ultrastructural and functional aspects. Arthropod Struct. Dev. 2016, 45, 42–56. [Google Scholar] [CrossRef]
- Minder, A.M.; Hosken, D.J.; Ward, P.I. Co-evolution of male and female reproductive characters across the Scathophagidae (Diptera). J. Evol. Biol. 2005, 18, 60–69. [Google Scholar] [CrossRef]
- Dybas, L.K.; Dybas, H.S. Coadaptation and taxonomic differentiation of sperm and spermathecae in featherwing beetles. Evolution 1981, 35, 168–174. [Google Scholar] [CrossRef]
- García-González, F.; Simmons, L.W. Paternal indirect genetic effects on offspring viability and the benefits of polyandry. Curr. Biol. 2007, 17, 32–36. [Google Scholar]
- Bergsten, J.; Miller, K.B. Phylogeny of diving beetles reveals a coevolutionary arms race between the sexes. PLoS ONE 2007, 2, e522. [Google Scholar] [CrossRef] [PubMed]
- Dallai, R. Overview on spermatogenesis and sperm structure of Hexapoda. Arthropod Struct. Dev. 2014, 43, 257–290. [Google Scholar] [CrossRef] [PubMed]
- Morrow, E.H.; Gage, M.J.G. The evolution of sperm length in moths. Proc. R. Soc. Lond. B Biol. Sci. 2000, 267, 307–313. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dallai, R.; Mercati, D.; Fanciulli, P.P.; Lupetti, P. The Structure of the Female Genital System of the Diving Beetle Scarodytes halensis (Fabricius, 1787) (Hydroporinae, Dytiscidae), and the Organization of the Spermatheca and the Spermathecal Gland Complex. Insects 2023, 14, 282. https://doi.org/10.3390/insects14030282
Dallai R, Mercati D, Fanciulli PP, Lupetti P. The Structure of the Female Genital System of the Diving Beetle Scarodytes halensis (Fabricius, 1787) (Hydroporinae, Dytiscidae), and the Organization of the Spermatheca and the Spermathecal Gland Complex. Insects. 2023; 14(3):282. https://doi.org/10.3390/insects14030282
Chicago/Turabian StyleDallai, Romano, David Mercati, Pietro P. Fanciulli, and Pietro Lupetti. 2023. "The Structure of the Female Genital System of the Diving Beetle Scarodytes halensis (Fabricius, 1787) (Hydroporinae, Dytiscidae), and the Organization of the Spermatheca and the Spermathecal Gland Complex" Insects 14, no. 3: 282. https://doi.org/10.3390/insects14030282