
Effects of Traditional Orchard Abandonment and Landscape Context on the Beneficial Arthropod Community in a Mediterranean Agroecosystem


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling Design
2.3. Statistical Analyses
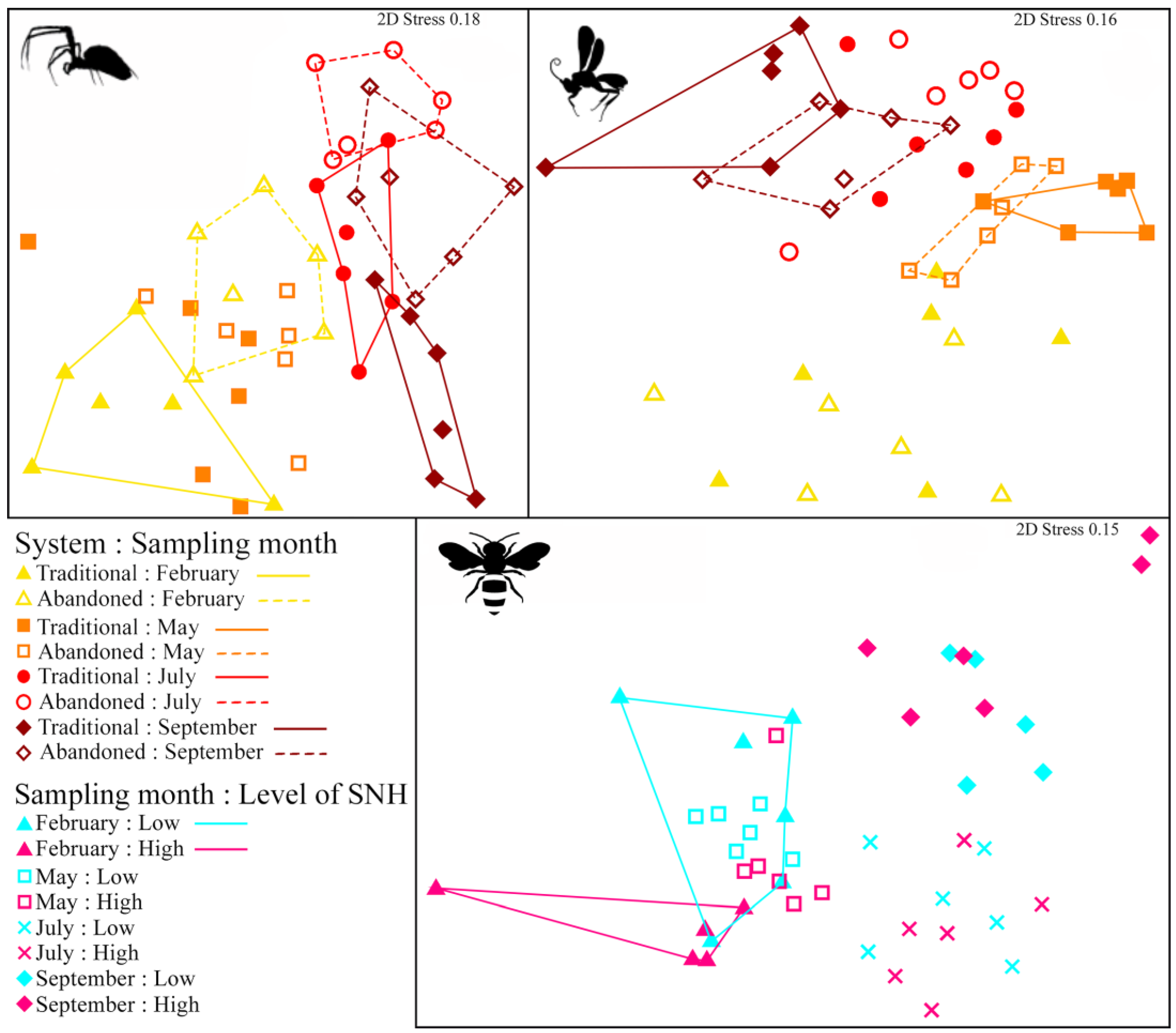
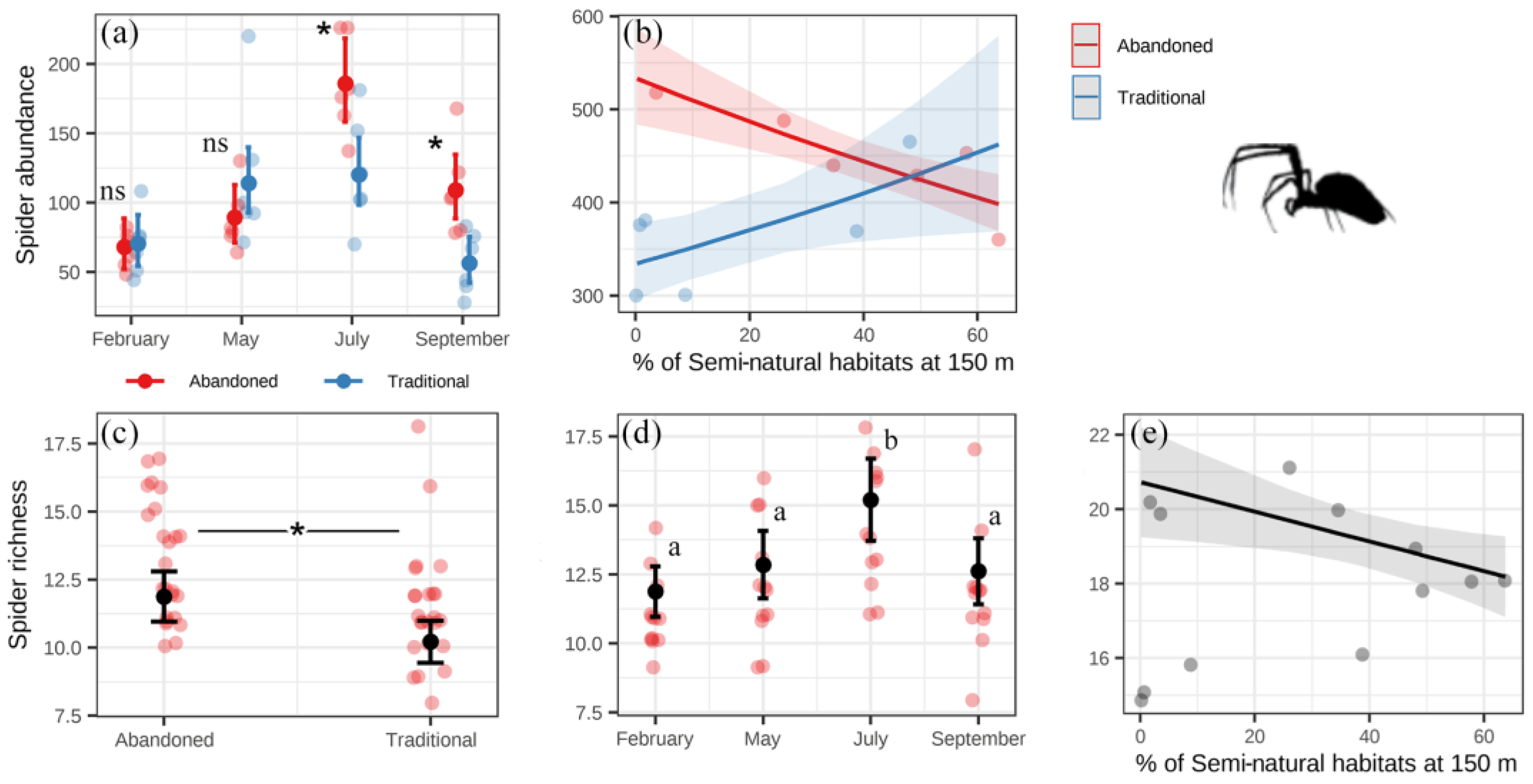
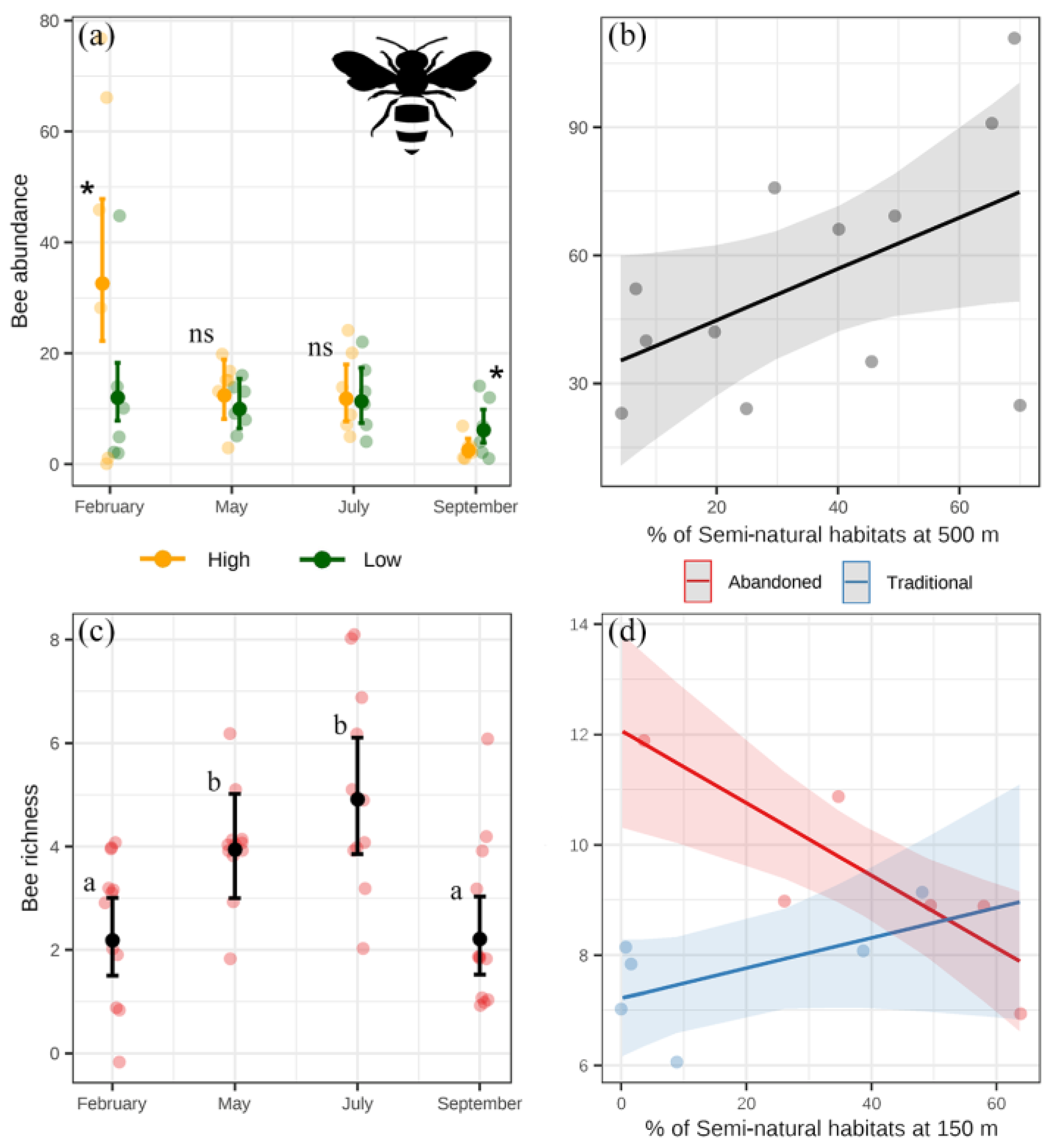
3. Results
4. Discussion
4.1. System Effects
4.2. Landscape Effects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural Sustainability and Intensive Production Practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- IPBES. Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. IPBES 2019, 44, 22–47. [Google Scholar]
- Raven, P.H.; Wagner, D.L. Agricultural Intensification and Climate Change Are Rapidly Decreasing Insect Biodiversity. Proc. Natl. Acad. Sci. USA 2021, 118, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide Decline of the Entomofauna: A Review of Its Drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Bengtsson, J.; Ahnström, J.; Weibull, A.C. The Effects of Organic Agriculture on Biodiversity and Abundance: A Meta-Analysis. J. Appl. Ecol. 2005, 42, 261–269. [Google Scholar] [CrossRef]
- Tscharntke, T.; Clough, Y.; Wanger, T.C.; Jackson, L.; Motzke, I.; Perfecto, I.; Vandermeer, J.; Whitbread, A. Global Food Security, Biodiversity Conservation and the Future of Agricultural Intensification. Biol. Conserv. 2012, 151, 53–59. [Google Scholar] [CrossRef]
- Rusch, A.; Chaplin-Kramer, R.; Gardiner, M.M.; Hawro, V.; Holland, J.; Landis, D.; Thies, C.; Tscharntke, T.; Weisser, W.W.; Winqvist, C.; et al. Agricultural Landscape Simplification Reduces Natural Pest Control: A Quantitative Synthesis. Agric. Ecosyst. Environ. 2016, 221, 198–204. [Google Scholar] [CrossRef] [Green Version]
- Batáry, P.; Dicks, L.V.; Kleijn, D.; Sutherland, W.J. The Role of Agri-Environment Schemes in Conservation and Environmental Management. Conserv. Biol. 2015, 29, 1006–1016. [Google Scholar] [CrossRef] [Green Version]
- Tscharntke, T.; Grass, I.; Wanger, T.C.; Westphal, C.; Batáry, P. Beyond Organic Farming—Harnessing Biodiversity-Friendly Landscapes. Trends Ecol. Evol. 2021, 36, 919–930. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agriculture Data. Available online: https://www.fao.org/faostat/en/#data (accessed on 5 September 2022).
- Plieninger, T.; Hui, C.; Gaertner, M.; Huntsinger, L. The Impact of Land Abandonment on Species Richness and Abundance in the Mediterranean Basin: A Meta-Analysis. PLoS ONE 2014, 9, e98355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, H.M.; Navarro, L.M. Rewilding European Landscapes; Pereira, H.M., Navarro, L.M., Eds.; Springer International Publishing: Cham, Switzerland, 2012; ISBN 978-3-319-12038-6. [Google Scholar]
- Oppermann, R.; Beaufoy, G.; Jones, G. High Nature Value Farming in Europe [35 European Countries—Experiences and Perspectives]; Verlag Regionalkultur: Ubstadt-Weiher, Germany, 2012; ISBN 9783897356573. [Google Scholar]
- Plieninger, T.; Bieling, C. Resilience-Based Perspectives to Guiding High-Nature-Value Farmland through Socioeconomic Change. Ecol. Soc. 2013, 18, 420. [Google Scholar] [CrossRef] [Green Version]
- Quintas-Soriano, C.; Buerkert, A.; Plieninger, T. Effects of Land Abandonment on Nature Contributions to People and Good Quality of Life Components in the Mediterranean Region: A Review. Land Use Policy 2022, 116, 106053. [Google Scholar] [CrossRef]
- Queiroz, C.; Beilin, R.; Folke, C.; Lindborg, R. Farmland Abandonment: Threat or Opportunity for Biodiversity Conservation? A Global Review. Front. Ecol. Environ. 2014, 12, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Benayas, J.M.R.; Martins, A.; Nicolau, J.M.; Schulz, J.J. Abandonment of Agricultural Land: An Overview of Drivers and Consequences. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2007, 2, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Penado, A.; Rebelo, H.; Goulson, D.; Wood, T.J.; Porto, M.; Rotheray, E.L.; Beja, P. From Pastures to Forests: Changes in Mediterranean Wild Bee Communities after Rural Land Abandonment. Insect. Conserv. Divers. 2022, 15, 325–336. [Google Scholar] [CrossRef]
- Walcher, R.; Karrer, J.; Sachslehner, L.; Bohner, A.; Pachinger, B.; Brandl, D.; Zaller, J.G.; Arnberger, A.; Frank, T. Diversity of Bumblebees, Heteropteran Bugs and Grasshoppers Maintained by Both: Abandonment and Extensive Management of Mountain Meadows in Three Regions across the Austrian and Swiss Alps. Landsc. Ecol. 2017, 32, 1937–1951. [Google Scholar] [CrossRef]
- Wersebeckmann, V.; Kolb, S.; Entling, M.H.; Leyer, I. Maintaining Steep Slope Viticulture for Spider Diversity. Glob. Ecol. Conserv. 2021, 29, e01727. [Google Scholar] [CrossRef]
- Hussain, R.I.; Walcher, R.; Brandl, D.; Jernej, I.; Arnberger, A.; Zaller, J.G.; Frank, T. Influence of Abandonment on Syrphid Assemblages in Mountainous Meadows. J. Appl. Entomol. 2018, 142, 450–456. [Google Scholar] [CrossRef]
- Walcher, R.; Hussain, R.I.; Karrer, J.; Bohner, A.; Brandl, D.; Zaller, J.G.; Arnberger, A.; Frank, T. Effects of Management Cessation on Hoverflies (Diptera: Syrphidae) across Austrian and Swiss Mountain Meadows. Web. Ecol. 2020, 20, 143–152. [Google Scholar] [CrossRef]
- Katayama, N.; Bouam, I.; Koshida, C.; Baba, Y.G. Biodiversity and Yield under Different Land-Use Types in Orchard/Vineyard Landscapes: A Meta-Analysis. Biol. Conserv. 2019, 229, 125–133. [Google Scholar] [CrossRef]
- Kajtoch, Ł. The Importance of Traditional Orchards for Breeding Birds: The Preliminary Study on Central European Example. Acta Oecologica 2017, 78, 53–60. [Google Scholar] [CrossRef]
- de Paz, V.; Tobajas, E.; Rosas-Ramos, N.; Tormos, J.; Asís, J.D.; Baños-Picón, L. Effect of Organic Farming and Agricultural Abandonment on Beneficial Arthropod Communities Associated with Olive Groves in Western Spain: Implications for Bactrocera Oleae Management. Insects 2022, 13, 48. [Google Scholar] [CrossRef]
- Bruggisser, O.T.; Schmidt-Entling, M.H.; Bacher, S. Effects of Vineyard Management on Biodiversity at Three Trophic Levels. Biol. Conserv. 2010, 143, 1521–1528. [Google Scholar] [CrossRef] [Green Version]
- Connell, J.H. Diversity in Tropical Rain Forests and Coral Reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grime, J.P. Competitive Exclusion in Herbaceous Vegetation. Nature 1973, 242, 344–347. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Penas, Á.; del Río, S.; Díaz González, T.E.; Rivas-Sáenz, S. Bioclimatology of the Iberian Peninsula and the Balearic Islands. In The Vegetation of the Iberian Peninsula; Loidi, J., Ed.; Springer International Publishing AG: Cham, Switzerland, 2017; Volume 1, pp. 29–80. [Google Scholar]
- Rosas-Ramos, N.; Asís, J.D.; Tobajas, E.; de Paz, V.; Baños-Picón, L. Disentangling the Benefits of Organic Farming for Beetle Communities (Insecta: Coleoptera) in Traditional Fruit Orchards. Agriculture 2022, 12, 243. [Google Scholar] [CrossRef]
- Zamora, J.; Verdú, J.R.; Galante, E. Species Richness in Mediterranean Agroecosystems: Spatial and Temporal Analysis for Biodiversity Conservation. Biol. Conserv. 2007, 134, 113–121. [Google Scholar] [CrossRef]
- Tscharntke, T.; Tylianakis, J.M.; Rand, T.A.; Didham, R.K.; Fahrig, L.; Batáry, P.; Bengtsson, J.; Clough, Y.; Crist, T.O.; Dormann, C.F.; et al. Landscape Moderation of Biodiversity Patterns and Processes—Eight Hypotheses. Biol. Rev. 2012, 87, 661–685. [Google Scholar] [CrossRef]
- Langellotto, G.A.; Denno, R.F. Responses of Invertebrate Natural Enemies to Complex-Structured Habitats: A Meta-Analytical Synthesis. Oecologia 2004, 139, 1–10. [Google Scholar] [CrossRef]
- Chaplin-Kramer, R.; Kremen, C. Pest Control Experiments Show Benefits of Complexity at Landscape and Local Scales. Ecol. Appl. 2012, 22, 1936–1948. [Google Scholar] [CrossRef]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat Management to Conserve Natural Enemies of Arthropod Pests in Agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Holland, J.M.; Bianchi, F.J.; Entling, M.H.; Moonen, A.-C.; Smith, B.M.; Jeanneret, P. Structure, Function and Management of Semi-Natural Habitats for Conservation Biological Control: A Review of European Studies. Pest Manag. Sci. 2016, 72, 1638–1651. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, C.M.; Lonsdorf, E.; Neel, M.C.; Williams, N.M.; Ricketts, T.H.; Winfree, R.; Bommarco, R.; Brittain, C.; Burley, A.L.; Cariveau, D.; et al. A Global Quantitative Synthesis of Local and Landscape Effects on Wild Bee Pollinators in Agroecosystems. Ecol. Lett. 2013, 16, 584–599. [Google Scholar] [CrossRef]
- Maurer, C.; Sutter, L.; Martínez-Núñez, C.; Pellissier, L.; Albrecht, M. Different Types of Semi-natural Habitat Are Required to Sustain Diverse Wild Bee Communities across Agricultural Landscapes. J. Appl. Ecol. 2022, 59, 2604–2615. [Google Scholar] [CrossRef]
- Westrich, P. Habitat Requirements of Central European Bees and the Problems of Partial Habitats. In The Conservation of Bees; Matheson, A., Buchmann, S.L., O’Toole, C., Westrich, P., Williams, I.H., Eds.; Academic Press: London, UK, 1996; pp. 1–16. [Google Scholar]
- Randlkofer, B.; Obermaier, E.; Hilker, M.; Meiners, T. Vegetation Complexity-The Influence of Plant Species Diversity and Plant Structures on Plant Chemical Complexity and Arthropods. Basic Appl. Ecol. 2010, 11, 383–395. [Google Scholar] [CrossRef]
- Öckinger, E.; Eriksson, A.K.; Smith, H.G. Effects of Grassland Abandonment, Restoration and Management on Butterflies and Vascular Plants. Biol. Conserv. 2006, 133, 291–300. [Google Scholar] [CrossRef]
- Tscharntke, T.; Rand, T.A.; Bianchi, F.J.J.A. The Landscape Context of Trophic Interactions: Insect Spillover across the Crop-Noncrop Interface. Ann. Zool. Fenn. 2005, 42, 421–432. [Google Scholar]
- Carrié, R.; Andrieu, E.; Cunningham, S.A.; Lentini, P.E.; Loreau, M.; Ouin, A. Relationships among Ecological Traits of Wild Bee Communities along Gradients of Habitat Amount and Fragmentation. Ecography 2017, 40, 85–97. [Google Scholar] [CrossRef]
- Rusch, A.; Valantin-Morison, M.; Sarthou, J.-P.; Roger-Estrade, J. Biological Control of Insect Pests in Agroecosystems. In Advances in Agronomy; Elsevier Ltd.: Amsterdam, The Netherlands, 2010; Volume 109, pp. 219–259. [Google Scholar]
- Holland, J.M.; Douma, J.C.; Crowley, L.; James, L.; Kor, L.; Stevenson, D.R.W.; Smith, B.M. Semi-Natural Habitats Support Biological Control, Pollination and Soil Conservation in Europe. A Review. Agron. Sustain. Dev. 2017, 37, 31. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Xiao, H.; Luo, S.; Hodgson, J.A.; Bianchi, F.J.J.A.; He, H.; van der Werf, W.; Zou, Y. Can Landscape Level Semi-Natural Habitat Compensate for Pollinator Biodiversity Loss Due to Farmland Consolidation? Agric. Ecosyst. Environ. 2021, 319, 107519. [Google Scholar] [CrossRef]
- Duelli, P.; Obrist, M.K. Regional Biodiversity in an Agricultural Landscape: The Contribution of Semi-Natural Habitat Islands. Basic Appl. Ecol. 2003, 4, 129–138. [Google Scholar] [CrossRef]
- Picchi, M.S.; Bocci, G.; Petacchi, R.; Entling, M.H. Taxonomic and Functional Differentiation of Spiders in Habitats in a Traditional Olive Producing Landscape in Italy. Eur. J. Entomol. 2020, 117, 18–26. [Google Scholar] [CrossRef]
- Rey, P.J.; Manzaneda, A.J.; Valera, F.; Alcántara, J.M.; Tarifa, R.; Isla, J.; Molina-Pardo, J.L.; Calvo, G.; Salido, T.; Gutiérrez, J.E.; et al. Landscape-Moderated Biodiversity Effects of Ground Herb Cover in Olive Groves: Implications for Regional Biodiversity Conservation. Agric. Ecosyst. Environ. 2019, 277, 61–73. [Google Scholar] [CrossRef]
- Shackelford, G.; Steward, P.R.; Benton, T.G.; Kunin, W.E.; Potts, S.G.; Biesmeijer, J.C.; Sait, S.M. Comparison of Pollinators and Natural Enemies: A Meta-Analysis of Landscape and Local Effects on Abundance and Richness in Crops. Biol. Rev. 2013, 88, 1002–1021. [Google Scholar] [CrossRef]
- Baldock, D.; Beaufoy, G.; Bennett, G.; Clark, J. Nature Conservation and New Directions in the Common Agricultural Policy: The Potential Role of EC Policies in Maintaining Farming and Management Systems of High Nature Value in the Community; Institute for European Environmental Policy (IEEP): London, UK, 1993; ISBN 90-74095-03-8. [Google Scholar]
- Fischer, J.; Hartel, T.; Kuemmerle, T. Conservation Policy in Traditional Farming Landscapes. Conserv. Lett. 2012, 5, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Keenleyside, C.; Beaufoy, G.; Tucker, G.; Jones, G. High Nature Value Farming throughout EU-27 and Its Financial Support under the CAP; Institute for European Environmental Policy (IEEP): London, UK, 2014; ISBN 978-9-27937-958-1. [Google Scholar]
- Horak, J.; Peltanova, A.; Podavkova, A.; Safarova, L.; Bogusch, P.; Romportl, D.; Zasadil, P. Biodiversity Responses to Land Use in Traditional Fruit Orchards of a Rural Agricultural Landscape. Agric. Ecosyst. Environ. 2013, 178, 71–77. [Google Scholar] [CrossRef]
- Gradziel, T.M. Almonds: Botany, Production and Uses; CABI: Wallingford, CT, USA, 2017; ISBN 978-1-78064-354-0. [Google Scholar]
- Eilers, E.J.; Klein, A.-M. Landscape Context and Management Effects on an Important Insect Pest and Its Natural Enemies in Almond. Biol. Control. 2009, 51, 388–394. [Google Scholar] [CrossRef]
- Henselek, Y.; Eilers, E.J.; Kremen, C.; Hendrix, S.D.; Klein, A.M. Pollination Requirements of Almond (Prunus Dulcis): Combining Laboratory and Field Experiments. J. Econ. Entomol. 2018, 111, 1006–1013. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of Pollinators in Changing Landscapes for World Crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding Pollinators and Their Values to Human Well-Being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benhadi-Marin, J.; Pereira, J.A.; Barrientos, J.A.; Bento, A.; Santos, S.A.P. Diversity of Predaceous Arthropods in the Almond Tree Canopy in Northeastern Portugal: A Methodological Approach. Entomol. Sci. 2011, 14, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Michalko, R.; Pekár, S.; Dul’a, M.; Entling, M.H. Global Patterns in the Biocontrol Efficacy of Spiders: A Meta-analysis. Glob. Ecol. Biogeogr. 2019, 28, 1366–1378. [Google Scholar] [CrossRef]
- Birkhofer, K.; Entling, M.H.; Lubin, Y. Agroecology: Trait Composition, Spatial Relationships, Trophic Interactions. In Spider Research in the 21st Century: Trends & Perspectives; Penney, D., Ed.; SIRI Scientific Press: Rochdale, UK, 2013; pp. 220–228. [Google Scholar]
- Schäckermann, J.; Pufal, G.; Mandelik, Y.; Klein, A.M. Agro-Ecosystem Services and Dis-Services in Almond Orchards Are Differentially Influenced by the Surrounding Landscape. Ecol. Entomol. 2015, 40, 12–21. [Google Scholar] [CrossRef]
- Sánchez-Ramos, I.; Marcotegui, A.; Pascual, S.; Fernández, C.E.; Cobos, G.; González-Núñez, M. Compatibility of Organic Farming Treatments against Monosteira Unicostata with Non-Target Arthropod Fauna of Almond Trees Canopy. Span. J. Agric. Res. 2017, 15, e1004. [Google Scholar] [CrossRef] [Green Version]
- Legner, E.F.; Gordh, G. Lower Navel Orange Worm (Lepidoptera: Phycitidae) Population Densities Following Establishment of Goniozus Legneri (Hymenoptera: Bethylidae) in California. J. Econ. Entomol. 1992, 85, 2153–2160. [Google Scholar] [CrossRef]
- Spears, L.R.; MacMahon, J.A. An Experimental Study of Spiders in a Shrub-Steppe Ecosystem: The Effects of Prey Availability and Shrub Architecture. J. Arachnol. 2012, 40, 218–227. [Google Scholar] [CrossRef]
- Rosas-Ramos, N.; Baños-Picón, L.; Tobajas, E.; de Paz, V.; Tormos, J.; Asís, J.D. Value of Ecological Infrastructure Diversity in the Maintenance of Spider Assemblages: A Case Study of Mediterranean Vineyard Agroecosystems. Agric. Ecosyst. Env. 2018, 265, 244–253. [Google Scholar] [CrossRef]
- Sanz, C.; Mata, R.; Gómez, J.; Allende, F.; López, N.; Molina, P.; Galiana, L. Atlas de Los Paisajes de España; Ministerio de Medio Ambiente: Madrid, Spain, 2003.
- ITACYL. AEMET Inicio—Atlas Agroclimático—ITACyL Portal Web. Available online: http://www.atlas.itacyl.es/ (accessed on 20 February 2023).
- IUSS. Working Group WRB World Reference Base for Soil Resources. In International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 4th ed.; International Union of Soil Sciences (IUSS): Vienna, Austria, 2022. [Google Scholar]
- Cardenas, M.; Pascual, F.; Campos, M.; Pekar, S. The Spider Assemblage of Olive Groves Under Three Management Systems. Environ. Entomol. 2015, 44, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Avinent, L.; Llacer, G. Adaptación de Un Aspirador de Jardín Para La Captura de Insectos. Bol. Sanid. Veg. Plagas 1995, 21, 329–335. [Google Scholar]
- Westphal, C.; Bommarco, R.; Carré, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyörgyi, H.; Tscheulin, T.; et al. Measuring Bee Diversity in Different European Habitats and Biogeographical Regions. Ecol. Monogr. 2008, 78, 653–671. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, S.S.; Ortega, J.C.G.; dos Santos Ribas, L.G.; Lopes, V.G.; Bini, L.M. Higher Taxa Are Sufficient to Represent Biodiversity Patterns. Ecol. Indic. 2020, 111, 105994. [Google Scholar] [CrossRef]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. User’s Guide and Application. 2013. Available online: https://www.robertkcolwell.org/pages/1407-estimates (accessed on 2 February 2023).
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Use R! 2nd ed.; Springer International Publishing: New York, NY, USA, 2018; ISBN 978-3-319-71403-5. [Google Scholar]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science & Business Media: New York, NY, USA, 2009; ISBN 978-0-387-87457-9. [Google Scholar]
- Marrec, R.; le Roux, V.; Martin, L.; Lenoir, J.; Brunet, J.; Cousins, S.A.O.; de Smedt, P.; Deconchat, M.; Diekmann, M.; Ehrmann, S.; et al. Multiscale Drivers of Carabid Beetle (Coleoptera: Carabidae) Assemblages in Small European Woodlands. Glob. Ecol. Biogeogr. 2021, 30, 165–182. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R Package Version 0.4.6, 2022. Available online: https://CRAN.R-project.org/package=DHARMa (accessed on 2 February 2023).
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual; Plymouth Marine Laboratory: Plymouth, MA, USA, 2006. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Barton, P.S.; Evans, M.J.; Foster, C.N.; Cunningham, S.A.; Manning, A.D. Environmental and Spatial Drivers of Spider Diversity at Contrasting Microhabitats. Austral. Ecol. 2017, 42, 700–710. [Google Scholar] [CrossRef] [Green Version]
- Theron, K.J.; Gaigher, R.; Pryke, J.S.; Samways, M.J. High Quality Remnant Patches in a Complex Agricultural Landscape Sustain High Spider Diversity. Biol. Conserv. 2020, 243, 108480. [Google Scholar] [CrossRef]
- Markó, V.; Keresztes, B.; Fountain, M.T.; Cross, J.V. Prey Availability, Pesticides and the Abundance of Orchard Spider Communities. Biol. Control. 2009, 48, 115–124. [Google Scholar] [CrossRef]
- Rosas-Ramos, N.; Baños-Picón, L.; Tormos, J.; Asís, J.D. The Complementarity between Ecological Infrastructure Types Benefits Natural Enemies and Pollinators in a Mediterranean Vineyard Agroecosystem. Ann. Appl. Biol. 2019, 175, 193–201. [Google Scholar] [CrossRef]
- Benhadi-Marín, J.; Pereira, J.A.; Sousa, J.P.; Santos, S.A.P. Distribution of the Spider Community in the Olive Grove Agroecosystem (Portugal): Potential Bioindicators. Agric. For. Entomol. 2020, 22, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, P.; Pekár, S.; Jocqué, R.; Coddington, J.A. Global Patterns of Guild Composition and Functional Diversity of Spiders. PLoS ONE 2011, 6, e21710. [Google Scholar] [CrossRef] [Green Version]
- Rosas-Ramos, N.; Baños-Picón, L.; Tormos, J.; Asís, J.D. Farming System Shapes Traits and Composition of Spider Assemblages in Mediterranean Cherry Orchards. PeerJ 2020, 8, e8856. [Google Scholar] [CrossRef] [Green Version]
- Cardenas, M.; Castro, J.; Campos, M. Short-Term Response of Soil Spiders to Cover-Crop Removal in an Organic Olive Orchard in a Mediterranean Setting. J. Insect Sci. 2012, 12, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, M.A.K.; Gurr, G.M.; Wratten, S.D. Beyond Nectar Provision: The Other Resource Requirements of Parasitoid Biological Control Agents. Entomol. Exp. Appl. 2016, 159, 207–221. [Google Scholar] [CrossRef]
- Gómez, J.E.; Lohmiller, J.; Joern, A. Importance of Vegetation Structure to the Assembly of an Aerial Web-Building Spider Community in North American Open Grassland. J. Arachnol. 2016, 44, 28. [Google Scholar] [CrossRef]
- Petanidou, T.; Lamborn, E. A Land for Flowers and Bees: Studying Pollination Ecology in Mediterranean Communities. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2005, 139, 279–294. [Google Scholar] [CrossRef]
- Roulston, T.H.; Goodell, K. The Role of Resources and Risks in Regulating Wild Bee Populations. Annu. Rev. Entomol. 2011, 56, 293–312. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Williams, N.; Kremen, C. Effects of Cultivation and Proximity to Natural Habitat on Ground-Nesting Native Bees in California Sunflower Fields. J. Kans Entomol. Soc. 2006, 79, 309–320. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Booij, C.J.H.; Tscharntke, T. Sustainable Pest Regulation in Agricultural Landscapes: A Review on Landscape Composition, Biodiversity and Natural Pest Control. Proc. R. Soc. B Biol. Sci. 2006, 273, 1715–1727. [Google Scholar] [CrossRef] [Green Version]
- Chaplin-Kramer, R.; O’Rourke, M.E.; Blitzer, E.J.; Kremen, C. A Meta-Analysis of Crop Pest and Natural Enemy Response to Landscape Complexity. Ecol. Lett. 2011, 14, 922–932. [Google Scholar] [CrossRef]
- Marino, P.C.; Landis, D.A. Effect of Landscape Structure on Parasitoid Diversity and Parasitism in Agroecosystems. Ecol. Appl. 1996, 6, 276–284. [Google Scholar] [CrossRef]
- Thies, C.; Tscharntke, T. Landscape Structure and Biological Control in Agroecosystems. Science 1999, 285, 893–895. [Google Scholar] [CrossRef]
- Thies, C.; Roschewitz, I.; Tscharntke, T. The Landscape Context of Cereal Aphid–Parasitoid Interactions. Proc. R. Soc. B Biol. Sci. 2005, 272, 203–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, F.J.J.A.; van Wingerden, W.K.R.E.; Griffioen, A.J.; van der Veen, M.; van der Straten, M.J.J.; Wegman, R.M.A.; Meeuwsen, H.A.M. Landscape Factors Affecting the Control of Mamestra Brassicae by Natural Enemies in Brussels Sprout. Agric. Ecosyst. Environ. 2005, 107, 145–150. [Google Scholar] [CrossRef]
- Kruess, A.; Tscharntke, T. Habitat Fragmentation, Species Loss, and Biological Control. Science 1994, 264, 1581–1584. [Google Scholar] [CrossRef] [PubMed]
- Mates, S.G.; Perfecto, I.; Badgley, C. Parasitoid Wasp Diversity in Apple Orchards along a Pest-Management Gradient. Agric. Ecosyst. Environ. 2012, 156, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, M.; Buckley, H.L.; Case, B.S.; Wratten, S.D.; Hale, R.J.; Didham, R.K. Agricultural Intensification Drives Landscape-Context Effects on Host-Parasitoid Interactions in Agroecosystems. J. Appl. Ecol. 2012, 49, 706–714. [Google Scholar] [CrossRef]
- Eeraerts, M.; van den Berge, S.; Proesmans, W.; Verheyen, K.; Smagghe, G.; Meeus, I. Fruit Orchards and Woody Semi-Natural Habitat Provide Complementary Resources for Pollinators in Agricultural Landscapes. Landsc. Ecol. 2021, 36, 1377–1390. [Google Scholar] [CrossRef]
- Holzschuh, A.; Steffan-Dewenter, I.; Tscharntke, T. How Do Landscape Composition and Configuration, Organic Farming and Fallow Strips Affect the Diversity of Bees, Wasps and Their Parasitoids? J. Anim. Ecol. 2010, 79, 491–500. [Google Scholar] [CrossRef]
- Steffan-Dewenter, I.; Leschke, K. Effects of Habitat Management on Vegetation and Above-Ground Nesting Bees and Wasps of Orchard Meadows in Central Europe. Biodivers. Conserv. 2003, 12, 1953–1968. [Google Scholar] [CrossRef]
- Priyadarshana, T.S.; Lee, M.B.; Ascher, J.S.; Qiu, L.; Goodale, E. Crop Heterogeneity Is Positively Associated with Beneficial Insect Diversity in Subtropical Farmlands. J. Appl. Ecol. 2021, 58, 2747–2759. [Google Scholar] [CrossRef]
- Inclán, D.J.; Cerretti, P.; Marini, L. Landscape Composition Affects Parasitoid Spillover. Agric. Ecosyst. Environ. 2015, 208, 48–54. [Google Scholar] [CrossRef]
- Marja, R.; Tscharntke, T.; Batáry, P. Increasing Landscape Complexity Enhances Species Richness of Farmland Arthropods, Agri-Environment Schemes Also Abundance—A Meta-Analysis. Agric. Ecosyst. Environ. 2022, 326, 107822. [Google Scholar] [CrossRef]
- Rischen, T.; Kaffenberger, M.; Plath, E.; Wolff, J.; Fischer, K. Configurational Landscape Heterogeneity: Crop-Fallow Boundaries Enhance the Taxonomic Diversity of Carabid Beetles and Spiders. Agric. Ecosyst. Environ. 2023, 341, 108194. [Google Scholar] [CrossRef]
- EEA (European Environmental Agency). Distribution and Targeting of the CAP Budget from a Biodiversity Perspective; Technical Report No 12/2009; European Environment Agency: Luxembourg, 2009.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response Variable | Explanatory Variables | d.f. | Pseudo-F | p-Value |
|---|---|---|---|---|
| Spiders | System | 1 | 5.761 | 0.001 |
| Sampling month | 3 | 10.754 | 0.001 | |
| System: Sampling month | 3 | 2.114 | 0.003 | |
| Parasitoids | System | 1 | 1.549 | 0.177 |
| Sampling month | 3 | 12.696 | 0.001 | |
| System: Sampling month | 3 | 2.351 | 0.018 | |
| Bees | Sampling month | 3 | 13.263 | 0.001 |
| Level of SNH | 1 | 2.242 | 0.041 | |
| Sampling month: Level of SNH | 3 | 1.466 | 0.103 | |
| Pairwise comparisons | Pseudo-t | p-value | ||
| Spiders | Traditional—Abandoned, February = 0 | 2.102 | 0.002 | |
| Traditional—Abandoned, May = 0 | 1.295 | 0.100 | ||
| Traditional—Abandoned, July = 0 | 1.773 | 0.023 | ||
| Traditional—Abandoned, September = 0 | 1.987 | 0.004 | ||
| Parasitoids | Traditional—Abandoned, February = 0 | 0.835 | 0.641 | |
| Traditional—Abandoned, May = 0 | 1.830 | 0.025 | ||
| Traditional—Abandoned, July = 0 | 1.221 | 0.171 | ||
| Traditional—Abandoned, September = 0 | 2.009 | 0.001 | ||
| Bees | Low—High, February = 0 | 1.998 | 0.018 | |
| Low—High, May = 0 | 1.143 | 0.283 | ||
| Low—High, July = 0 | 0.974 | 0.477 | ||
| Low—High, September = 0 | 0.653 | 0.664 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Paz, V.; Asís, J.D.; Holzschuh, A.; Baños-Picón, L. Effects of Traditional Orchard Abandonment and Landscape Context on the Beneficial Arthropod Community in a Mediterranean Agroecosystem. Insects 2023, 14, 277. https://doi.org/10.3390/insects14030277
de Paz V, Asís JD, Holzschuh A, Baños-Picón L. Effects of Traditional Orchard Abandonment and Landscape Context on the Beneficial Arthropod Community in a Mediterranean Agroecosystem. Insects. 2023; 14(3):277. https://doi.org/10.3390/insects14030277
Chicago/Turabian Stylede Paz, Víctor, Josep D. Asís, Andrea Holzschuh, and Laura Baños-Picón. 2023. "Effects of Traditional Orchard Abandonment and Landscape Context on the Beneficial Arthropod Community in a Mediterranean Agroecosystem" Insects 14, no. 3: 277. https://doi.org/10.3390/insects14030277