

Using MaxEnt Model to Predict the Potential Distribution of Three Potentially Invasive Scarab Beetles in China
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Source
2.2. Occurrence Data
2.3. Bioclimatic Variables
2.4. Modeling Methods
3. Results
3.1. Modeling Results Validation
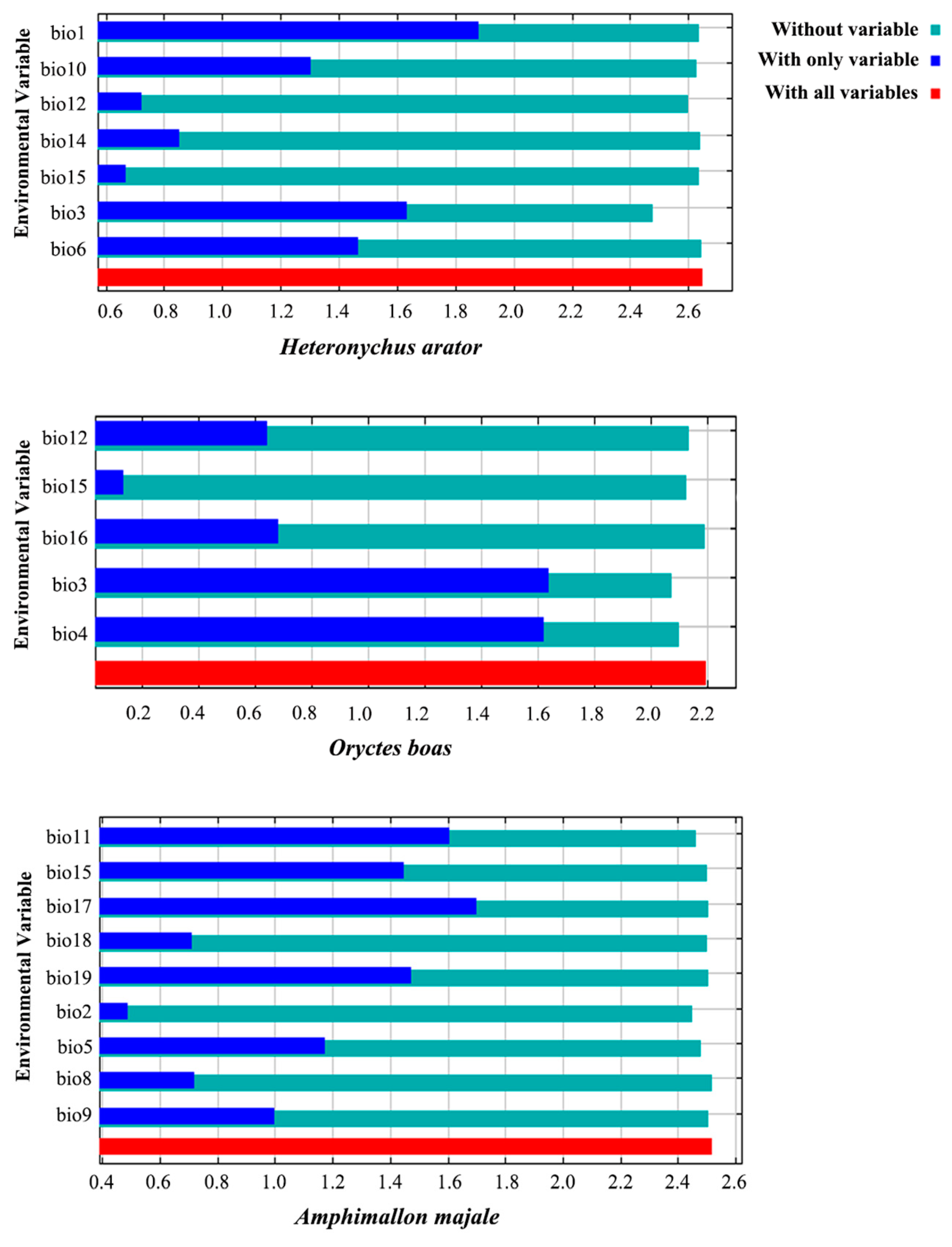
3.2. Key Bioclimatic Variables
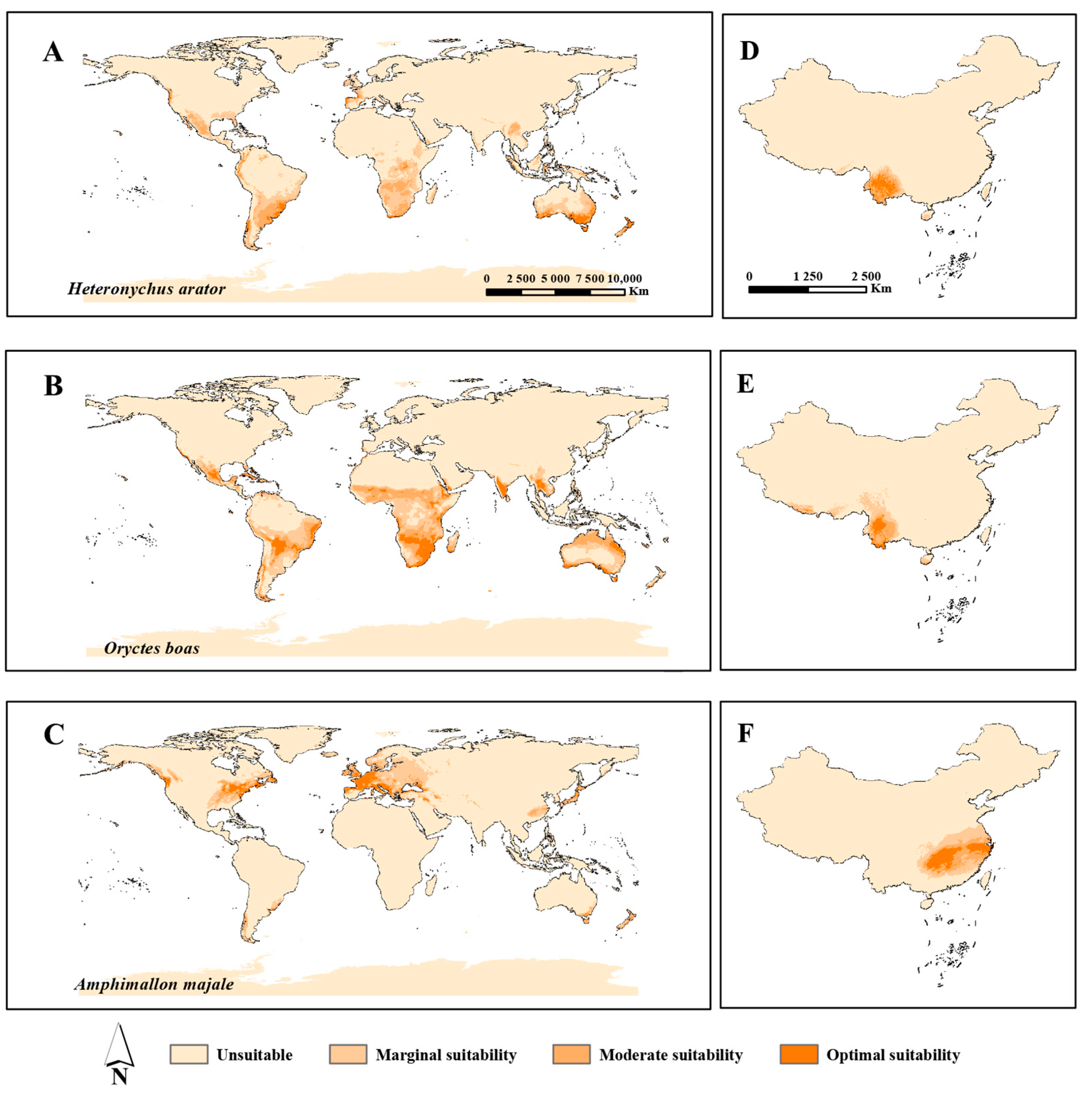
3.3. Potential Geographical Distribution
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef]
- Lv, F.; Du, Y.Z.; Zhou, Y.; Yang, W.Y. Research summary of pest risk analysis. Plant Quar. 2016, 30, 7–12. [Google Scholar]
- Peng, L.; Wan, F.H.; Hou, Y.M. Research progress on prevention and control of invasive insects in China. Chin. J. Appl. Entomol. 2020, 57, 244–258. [Google Scholar]
- Jin, S.; Li, R.F.; Chu, W.; Fan, G.H. Analysis on pests intercepted from imported commodity during 2013–2014 and the proposal for plant quarantine. Plant Quar. 2016, 30, 72–75. [Google Scholar]
- Kolar, C.S.; Lodge, D.M. Progress in invasion biology: Predicting invaders. Trends Ecol. Evol. 2001, 16, 199–204. [Google Scholar] [CrossRef]
- Waage, J.K.; Reaser, J.K. A global strategy to defeat invasive species. Science 2001, 292, 1477–1486. [Google Scholar] [CrossRef]
- Scholtz, C.H.; Grebennikov, V.V. 12 Scarabaeiformia Crowson, 1960. In Handbook of Zoology; Beutel, R.G., Kristensen, N.P., Eds.; De Gruyter: Berlin, Germany, 2016; Part 38 Volume IV, pp. 345–425. [Google Scholar]
- Fleming, W.E. Biology of the Japanese Beetle; USDA Technical Bulletins: Washington, DC, USA, 1972; Volume 1449, pp. 1–129. [Google Scholar]
- Potter, D.A.; Held, D.W. Biology and management of the Japanese beetle. Annu. Rev. Entomol. 2002, 47, 175–205. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.F.; Xu, Y.; Huang, X.K.; Han, X.M.; Xu, R.M. Prediction of suitable areas for the Japanses Beetle in China. Plant Quar. 2002, 2, 73–77. [Google Scholar]
- United States Department of Agricultural (USDA) Managing the Japanese Beetle: A Homeowner’s Handbook. APHIS 81-25-003. Available online: https://www.aphis.usda.gov/plant_health/plant_pest_info/jb/downloads/JBhandbook.pdf (accessed on 24 March 2018).
- Kistner-Thomas, E.J. The potential global distribution and voltinism of the Japanese beetle (Coleoptera: Scarabaeidae) under current and future climates. J. Insect Sci. 2019, 19, 16. [Google Scholar] [CrossRef]
- Aidoo, O.F.; Hao, M.; Ding, F.; Wang, D.; Jiang, D.; Ma, T.; Qian, Y.; Tettey, E.; Yankey, N.; Dadzie Ninsin, K.; et al. The Impact of Climate Change on Potential Invasion Risk of Oryctes monoceros Worldwide. Front. Ecol. Evol. 2022, 10, 895906. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, J.S.; Ren, G.; Zhao, K.X.; Wang, X.F. Global potential distribution prediction of Xanthium italicum based on Maxent model. Sci. Rep. 2021, 11, 16545. [Google Scholar] [CrossRef]
- Abdallah, M.; Mwatawala, M.W.; Kudra, A.B.; Urio, N.A.; Mtakwa, P.W. Damage and control of the invasive African black beetle Heteronychus arator F.(Coleoptera: Scarabaeidae) in Southern highlands of Tanzania. Int. J. Pest Manag. 2018, 64, 88–93. [Google Scholar] [CrossRef]
- CABI. Invasive Species Compendium; CAB International: Wallingford, UK. Available online: https://www.cabi.org/isc/ (accessed on 10 October 2022).
- Matthiessen, J.N.; Ridsdill-Smith, T.J. Populations of African black beetle, Heteronychus arator (Coleoptera: Scarabaeidae) in a Mediterranean climate region of Australia. Bull. Entomol. Res. 1991, 81, 85–91. [Google Scholar] [CrossRef]
- Ross, K.M. Resistance to African Black Beetle (Heteronychus arator) in Perennial Ryegrass (Lolium perenne L.) Infected with AR1 Endophyte; University of Waikato: Hamilton, New Zealand, 2016. [Google Scholar]
- EPPO. EPPO Global Database. Available online: https://gd.eppo.int (accessed on 10 October 2022).
- Bedford, G.O. Biology and management of palm dynastid beetles: Recent advances. Annu. Rev. Entomol 2013, 58, 353–372. [Google Scholar] [CrossRef]
- Gries, G.; Gries, R.; Perez, A.L.; Oehlschlager, A.C.; Gonzales, L.M.; Pierce, H.D. Aggregation Pheromone of the African Rhinoceros Beetle, Oryctes monoceros (Olivier) (Coleoptera: Scarabaeidae). Z. Für Nat. C 1994, 49, 363–366. [Google Scholar] [CrossRef]
- Mansfield-Aders, W. Insects injurious to economic crops in the Zanzibar Protectorate. Bull. Entomol. Res. 1920, 10, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Cappaert, D.L.; Koppenhöfer, A.M. Steinernema scarabaei, an entomopathogenic nematode for control of the European chafer. Biol. Control 2003, 28, 379–386. [Google Scholar] [CrossRef]
- Renkema, J.M.; Nyrop, J.P.; Difonzo, C.; Sears, M.K.; Schaafsma, A.W. Control decision rule for European chafer (Coleoptera: Scarabaeidae) larvae in field corn. J. Econ. Entomol. 2006, 99, 76–84. [Google Scholar] [CrossRef]
- Bughrara, S.S.; Smitley, D.R.; Cappaert, D. European chafer grub feeding on warm-season and cool-season turfgrasses, native prairie grasses, and Pennsylvania sedge. HortTechnology 2008, 18, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Peterson, A.T.; Soberon, J.; Pearson, R.G.; Anderson, R.P.; Martınez-Meyer, E.; Nakamura, M.; Araujo, M.B. Ecological Niches and Geographic Distributions; Princeton University Press: Princeton, NJ, USA, 2011; p. 314. [Google Scholar]
- Yi, Y.; Cheng, X.; Yang, Z.; Wieprecht, S.; Zhang, S.; Wu, Y. Evaluating the ecological influence of hydraulic projects: A review of aquatic habitat suitability models. Renew. Sustain. Energy Rev. 2017, 68, 748–762. [Google Scholar] [CrossRef]
- Sunil, K.; Thomas, J.S. Maxent modeling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia. J. Ecol. Nat. Environ. 2009, 1, 094–098. [Google Scholar]
- Hou, N.; Dai, Q.; Ran, J.; Jiao, Y.; Cheng, Y.; Zhao, C. A corridor design for the giant panda in the Niba Mountain of China. Chin. J. App. Environ. Biol. 2014, 20, 1039–1045. [Google Scholar]
- Zhang, Q.; Zhao, L.; He, P.; Zheng, K.M.; Wu, P.X.; Fan, J.Y. The potential distribution analysis and economic loss assessmentof Spodoptera frugiperda (Smith) in Qujing, Yunnan Province. J. Yunnan Univ. Nat. Sci. Ed. 2020, 42, 1224–1229. [Google Scholar] [CrossRef]
- Shi, W.; Zhu, E.J.; Wang, Y.C.; Ma, F.Z.; He, Q.J.; Yi, C.H. Prediction of potentially suitable distribution area of Propomacrus davidi Deyrolle in China based on MaxEnt model. Chin. J. Ecol. 2021, 40, 2936–2944. [Google Scholar] [CrossRef]
- Dutrillaux, A.M.; Mamuris, Z.; Dutrillaux, B. Chromosome analyses challenge the taxonomic position of Augosoma centaurus Fabricius, 1775 (Coleoptera: Scarabaeidae: Dynastinae) and the separation of Dynastini and Oryctini. Zoosystema 2013, 35, 537–549. [Google Scholar] [CrossRef]
- Mohan, K.; Padmanaban, A.M. Diversity and Abundance of Coleopteran Insects in Bhavani Taluk Erode District, Tamil Nadu, India. Int. J. Innov. Bio-Sci. 2013, 3, 57–63. [Google Scholar]
- Ba-Angood, S. Date palm status and perspective in Yemen. In Date Palm Genetic Resources and Utilization; Al-Khayri, J., Jain, S., Johnson, D., Eds.; Springer: Dordrecht, The Netherlands, 2015; Volume 2, pp. 241–263. [Google Scholar] [CrossRef]
- Abdulla, B.S.; Ahmmed, S.H.; Mawlood, N.A.; Omar, Z.Z. A new record of European Chafer beetles, Amphimallon majale (Razoumowsky, 1789) (Coleoptera: Melolonthidae) From Iraq. Plant Arch. 2020, 20, 6357–6361. [Google Scholar]
- Baloch, M.N.; Fan, J.; Haseeb, M.; Zhang, R.Z. Mapping potential distribution of Spodoptera frugiperda (Lepidoptera: Noctuidae) in central Asia. Insects 2020, 11, 172. [Google Scholar] [CrossRef] [Green Version]
- Verbruggen, H.; Tyberghein, L.; Belton, G.S.; Mineur, F.; Jueterbock, A.; Hoarau, G.; Gurgel, C.F.; Clerk, O.D. Improving transferability of introduced species’distribution models: New tools to forecast the spread of a highly invasive seaweed. PLoS ONE 2013, 8, e63337. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. Available online: https://rmets.onlinelibrary.wiley.com/doi/abs/10.1002/joc.1276 (accessed on 28 September 2022). [CrossRef]
- Heikkinen, R.K.; Araújo, M.B.; Virkkala, R.; Thuiller, W.; Sykes, M.T. Methods and uncertainties in bioclimatic envelop modeling under climate change. Prog. Phys. Geogr. 2006, 30, 751–777. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.; Qi, G.J.; Ma, j.; Ren, Y.L.; Wang, R.; Mckirdy, S. Predicting the potential geographic distribution of Bactrocera bryoniae and Bactrocera neohumeralis (Diptera: Tephritidae) in China using MaxEnt ecological niche modeling. J. Integr. Agric. 2020, 19, 2072–2082. [Google Scholar] [CrossRef]
- Zhu, G.P.; Bu, W.J.; Gao, Y.B.; Liu, G.Q. Potential geographic distribution of brown marmorated stink bug invasion (Halyomorpha halys). PLoS ONE 2012, 7, e31246. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Qi, G.J.; Chen, T.; Gao, Y.; Lei, Y.Y.; Lv, L.H. Potential geographic distribution of Planococcus minor and P. lilacinus in China based on Maxent. J. Environ. Entomol. 2015, 37, 219–223. [Google Scholar]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Ge, Y.H.; Ye, J.R.; Liang, Z.W.; Lin, S.X.; Zhang, Y.X. Prediction of potential distribution of Xylotrechus rusticus in China based on MaxEnt model. Plant Quar. 2021, 4, 65–70. [Google Scholar]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jimenez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Mao, M.; Chen, S.; Ke, Z.; Qian, Z.; Xu, Y. Using MaxEnt to Predict the Potential Distribution of the Little Fire Ant (Wasmannia auropunctata) in China. Insects 2022, 13, 1008. [Google Scholar] [CrossRef]
- Gong, Z.; Ma, G.C.; Wen, H.B.; Peng, Z.Q. List of alien invasive insects in tropical regions of China. Chin. J. Appl. Entomol. 2021, 58, 27–48. [Google Scholar]
- Xian, X.Q.; Wang, R.; Guo, J.Y.; Liu, W.X.; Zhang, G.F.; Sun, Y.F.; Wan, F.H. Analysis of new invasive alien species in China’s agricultural and forestry ecosystems in recent 20 years. Plant Prot. 2018, 44, 168–175. [Google Scholar]
- Wei, J.F.; Zhao, Q.; Zhao, W.Q.; Zhang, H.F. Predicting the potential distributions of the invasive cycad scale Aulacaspis yasumatsui (Hemiptera: Diaspididae) under different climate change scenarios and the implications for management. PeerJ 2018, 6, e64832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinclair, S.J.; White, M.D.; Newell, G.R. How useful are species distribution models for managing biodiversity under future climates? Ecol. Soc. 2010, 15, 175–183. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P.; Araújo, M. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Pan, X.B.; Chen, K.; Huang, J.; Liang, B.Y. Data requirement for pest risk analysis and development and prospect of information system. Plant Quar. 2019, 33, 6–9. [Google Scholar] [CrossRef]
- Xian, X.Q.; Chen, H.; Zhao, J.; Wan, F.H. Introduction to the database of invasive alien species in China. Plant Prot. 2013, 39, 103–109. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Variable | Variable Description | Heteronychus arator | Oryctes boas | Amphimallon majale |
|---|---|---|---|---|
| Bio1 | Annual mean temperature | √ | ||
| Bio2 | Mean diurnal range | √ | ||
| Bio3 | Isothermality (Bio2/Bio7) (×100) | √ | √ | |
| Bio4 | Temperature seasonality | √ | √ | |
| Bio5 | Max temperature of the warmest month | √ | ||
| Bio6 | Min temperature of coldest month | |||
| Bio8 | Mean temperature of the wettest quarter | √ | ||
| Bio9 | Mean temperature of the driest quarter | √ | √ | |
| Bio10 | Mean temperature of warmest quarter | |||
| Bio11 | Mean temperature of the coldest quarter | √ | ||
| Bio12 | Annual precipitation | √ | ||
| Bio14 | Precipitation of driest month | √ | ||
| Bio15 | Precipitation seasonality | √ | √ | |
| Bio16 | Precipitation of wettest quarter | √ | ||
| Bio17 | Precipitation of driest quarter | √ | ||
| Bio18 | Precipitation of the warmest quarter | √ | √ | |
| Bio19 | Precipitation of the coldest quarter | √ |
| Variable | Percent Contribution | ||
|---|---|---|---|
| Heteronychus arator | Oryctes boas | Amphimallon majale | |
| Bio1 | 21.4 | ||
| Bio2 | 3.2 | ||
| Bio3 | 47.9 | 67.3 | |
| Bio4 | 4.5 | 12.7 | |
| Bio5 | 8.1 | ||
| Bio9 | 5.1 | 0.7 | |
| Bio11 | 16.6 | ||
| Bio12 | 13.8 | ||
| Bio14 | 17.7 | ||
| Bio15 | 0.4 | 3.2 | 4 |
| Bio16 | 3 | ||
| Bio17 | 52 | ||
| Bio18 | 3 | 2.6 | |
| Bio19 | 12.8 | ||
| Percentage | Species (World/China) | ||
|---|---|---|---|
| Heteronychus arator | Oryctes boas | Amphimallon majale | |
| Potential distribution area | 10.5%/3.8% | 14.8%/5.6% | 7.9%/9.5% |
| Optimal suitability area | 0.8%/1.1% | 2.3%/0.8% | 1.2%/2.4% |
| Moderate suitability area | 3.0%/1.4% | 5.3%/1.6% | 2.0%/2.9% |
| Margin suitability area | 6.7%/1.3% | 7.2%/3.2% | 4.7%/4.2% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Lu, Y.; Han, M.; Li, L.; He, P.; Shi, A.; Bai, M. Using MaxEnt Model to Predict the Potential Distribution of Three Potentially Invasive Scarab Beetles in China. Insects 2023, 14, 239. https://doi.org/10.3390/insects14030239
Wang S, Lu Y, Han M, Li L, He P, Shi A, Bai M. Using MaxEnt Model to Predict the Potential Distribution of Three Potentially Invasive Scarab Beetles in China. Insects. 2023; 14(3):239. https://doi.org/10.3390/insects14030239
Chicago/Turabian StyleWang, Shuangyi, Yuanyuan Lu, Mengyang Han, Lulu Li, Ping He, Aimin Shi, and Ming Bai. 2023. "Using MaxEnt Model to Predict the Potential Distribution of Three Potentially Invasive Scarab Beetles in China" Insects 14, no. 3: 239. https://doi.org/10.3390/insects14030239