
Dynamic Roles of Insect Carboxyl/Cholinesterases in Chemical Adaptation

Abstract
:Simple Summary
Abstract
1. Introduction
2. CCE Classification and Structural Characteristics
2.1. Classification of Insect CCEs
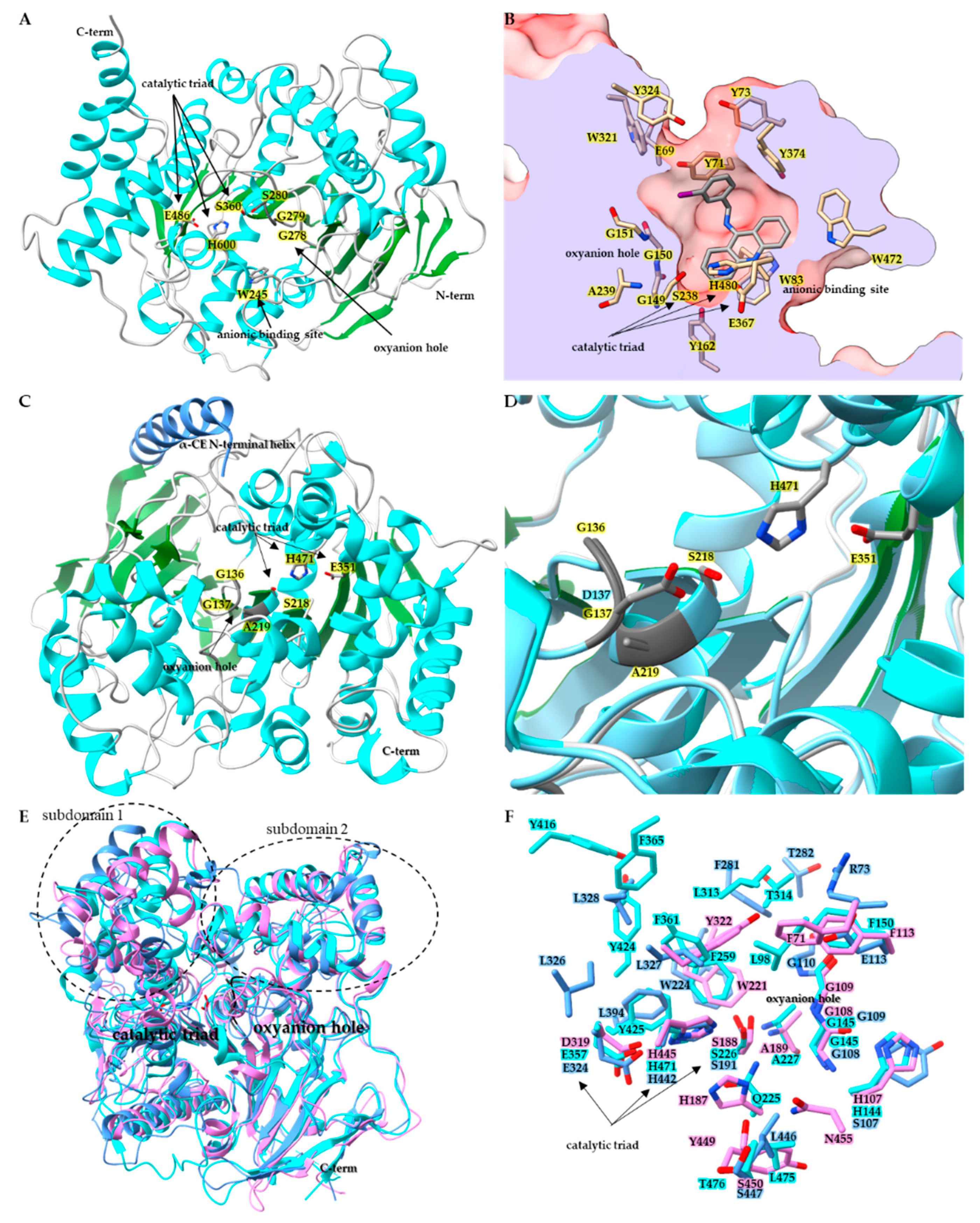
2.2. Structural Characteristics of Insect CCEs

{kind=link}
| Order | Species | Dietary/ Detoxification | Hormone/Semiochemical Process | Neurodevelopmental Process | Total | Reference | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| α-Esterase and Others (Clades A–C) | Integument Esterase (Clade D) | β-Esterase (Clade E) | JHE (Clades F and G) | Glutactin and Like Enzymes (Clade H) | Uncharacterized (Clade I) | AChE (Clade J) | Uncharacterized Neuroreceptors (Clade K) | Neuroligin (Clade L) | Gliotactin (Clade M) | Neurotactin (Clade N) | ||||
| Coleoptera | Leptinotarsa decemlineata | 52 | 8 | 3 | 1 | 1 | 1 | 2 | 1 | 2 | 1 | 0 | 72 | [25] |
| Tribolium castenaum | 26 | 3 | 4 | 2 | 1 | 1 | 2 | 2 | 5 | 1 | 0 | 47 | [25] | |
| Diptera | Aedes aegypti | 22 | 1 | 2 | 12 | 10 | 0 | 2 | 0 | 0 | 0 | 0 | 49 | [57] |
| Anopheles gambiae | 16 | 0 | 4 | 10 | 10 | 1 | 2 | 1 | 5 | 2 | 0 | 51 | [20] | |
| Culex quinquefasciatus | 30 | 1 | 3 | 22 | 6 | 0 | 2 | 1 | 3 | 1 | 2 | 61 | [40] | |
| Drosophila melanogaster | 13 | 3 | 2 | 3 | 5 | 1 | 1 | 1 | 4 | 2 | 0 | 35 | [20,58] | |
| Musca domestica | 17 | 7 | 2 | 1 | 5 | 0 | 1 | 1 | 3 | 2 | 0 | 39 | [14] | |
| Sphaerophoria rueppellii | 15 | 0 | 9 | 4 | 4 | 0 | 1 | 0 | 5 | 1 | 1 | 40 | [59] | |
| Hemiptera | Bemisia tabaci | 6 | 0 | 15 | 3 | 1 | 1 | 4 | 1 | 10 | 1 | 0 | 42 | [58] |
| Diaphorina citri | 2 | 0 | 2 | 4 | 3 | 0 | 2 | 1 | 6 | 0 | 0 | 20 | [60] | |
| Myzus persicae | 5 | 0 | 12 | 0 | 0 | 1 | 3 | 1 | 0 | 0 | 0 | 22 | [61] | |
| Nilaparvata lugens | 3 | 1 | 19 | 0 | 2 | 0 | 2 | 0 | 1 | 1 | 0 | 29 | [62,63] | |
| Orius laevigatus | 0 | 0 | 6 | 9 | 1 | 1 | 2 | 1 | 8 | 3 | 1 | 32 | [64] | |
| Pediculus humanus | 3 | 0 | 1 | 0 | 1 | 0 | 2 | 1 | 6 | 3 | 0 | 17 | [65,66] | |
| Rhodnius prolixus | 0 | 0 | 40 | 0 | 2 | 1 | 2 | 1 | 13 | 2 | 0 | 61 | [67] | |
| Triatoma dimidiata * | 0 | 0 | 25 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 27 | ||
| Triatoma infestans * | 0 | 0 | 18 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 19 | ||
| Triatoma pallidipennis * | 0 | 0 | 17 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 18 | ||
| Homoptera | Acyrthosiphon pisum | 5 | 0 | 18 | 0 | 0 | 1 | 2 | 1 | 3 | 0 | 0 | 30 | [61] |
| Hymenoptera | Apis mellifera | 8 | 1 | 3 | 1 | 0 | 2 | 2 | 1 | 5 | 1 | 0 | 24 | [68] |
| Nasonia vitripennis | 13 | 4 | 11 | 2 | 1 | 1 | 2 | 1 | 5 | 1 | 0 | 41 | [20] | |
| Lepidoptera | Bombyx mori | 55 | 0 | 2 | 4 | 0 | 1 | 2 | 1 | 6 | 1 | 2 | 74 | [69] |
| Plutella xylostella | 20 | 1 | 1 | 6 | 1 | 4 | 2 | 6 | 4 | 3 | 0 | 48 | [70,71] | |
| Spodoptera litura | 83 | 4 | 2 | 8 | 0 | 0 | 2 | 3 | 7 | 1 | 1 | 111 | [72] | |
| Class | Clade | Species Name | Protein Name | Catalytic Triad | Reference |
|---|---|---|---|---|---|
| Dietary/xenobiotic detoxification | α-esterases | Lucilia cuprina | LcαE7 (PDB: 4FNG) | Serine, glutamate, and histidine | [51] |
| Hormone/pheromone processing | β-esterases | Drosophila melanogaster | DmEsterase-6 (PDB: 5THM) | Serine, aspartate, and histidine | [73] |
| Culex qunquefasciatus | Cqestβ2 (PDB: 5W1U) | Serine, glutamate, and histidine | [53] | ||
| JHEs | Manduca sexta | MsJHE (PDB: 2FJ0) | Serine, glutamate, and histidine | [37] | |
| Neurodevelopmental | AChEs | Drosophila melanogaster | DmAChE (PDB: 1QO9) | Serine, glutamate, and histidine | [52] |
| Anopheles gambiae | AgAChE (PDB: 5X61) | Serine, glutamate, and histidine | [55,56] |
3. Dynamic Rules of Insect CCEs in Chemical Adaptation
3.1. CCE-Mediated Insecticide Resistance
3.2. Metabolism of Plant Allelochemicals
3.3. Odorant Degradation in the Olfactory System
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhu, F.; Gujar, H.; Gordon, J.R.; Haynes, K.F.; Potter, M.F.; Palli, S.R. Bed bugs evolved unique adaptive strategy to resist pyrethroid insecticides. Sci. Rep. 2013, 3, 1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Despres, L.; David, J.P.; Gallet, C. The evolutionary ecology of insect resistance to plant chemicals. Trends Ecol. Evol. 2007, 22, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Koirala, B.K.S.; Moural, T.; Zhu, F. Functional and structural diversity of insect glutathione S-transferases in xenobiotic adaptation. Int. J. Biol. Sci. 2022, 18, 5713–5723. [Google Scholar] [CrossRef] [PubMed]
- Brattsten, L.B. Biochemical defense mechansims in herbivores against plant allelochemicals. In Herbivores-Their Interaction with Secondary Plant Metabolites; Rosenthal, G.A., Janzen, D.H., Eds.; Academic Press: New York, NY, USA, 1979; pp. 199–270. [Google Scholar]
- Berenbaum, M.R.; Johnson, R.M. Xenobiotic detoxification pathways in honey bees. Curr. Opin. Insect. Sci. 2015, 10, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Cui, Y.; Walsh, D.B.; Lavine, L.C. Application of RNAi towards insecticide resistance management. In Short Views on Insect Biochemistry and Molecular Biology; Chandrasekar, R., Tyagi, B.K., Gui, Z., Reeck, G.R., Eds.; Academic Publisher: Manhattan, New York, NY, USA, 2014; Volume 2, pp. 595–619. [Google Scholar]
- Hamby, K.A.; Kwok, R.S.; Zalom, F.G.; Chiu, J.C. Integrating circadian activity and gene expression profiles to predict chronotoxicity of Drosophila suzukii response to insecticides. PLoS ONE 2013, 8, e68472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amezian, D.; Nauen, R.; Le Goff, G. Transcriptional regulation of xenobiotic detoxification genes in insects—An overview. Pestic. Biochem. Physiol. 2021, 174, 104822. [Google Scholar] [CrossRef]
- Montella, I.R.; Schama, R.; Valle, D. The classification of esterases: An important gene family involved in insecticide resistance—A review. Mem. Inst. Oswaldo Cruz 2012, 107, 437–449. [Google Scholar] [CrossRef] [Green Version]
- Marshall, S.D.; Putterill, J.J.; Plummer, K.M.; Newcomb, R.D. The carboxylesterase gene family from Arabidopsis thaliana. J. Mol. Evol. 2003, 57, 487–500. [Google Scholar]
- Satoh, T.; Hosokawa, M. The mammalian carboxylesterases: From molecules to functions. Annu. Rev. Pharmacol. Toxicol. 1998, 38, 257–288. [Google Scholar] [CrossRef]
- Bornscheuer, U.T. Microbial carboxyl esterases: Classification, properties and application in biocatalysis. FEMS Microbiol. Revs. 2002, 26, 73–81. [Google Scholar] [CrossRef]
- Oakeshott, J.G.; Claudianos, C.; Russell, R.J.; Robin, G.C. Carboxyl/cholinesterases: A case study of the evolution of a successful multigene family. BioEssays 1999, 21, 1031–1042. [Google Scholar] [CrossRef]
- Feng, X.; Li, M.; Liu, N. Carboxylesterase genes in pyrethroid resistant house flies, Musca domestica. Insect Biochem. Mol. Biol. 2018, 92, 30–39. [Google Scholar] [CrossRef]
- Wei, H.; Tan, S.; Li, Z.; Li, J.; Moural, T.W.; Zhu, F.; Liu, X. Odorant degrading carboxylesterases modulate foraging and mating behaviors of Grapholita molesta. Chemosphere 2021, 270, 128647. [Google Scholar] [CrossRef]
- Vogt, R.G. Molecular basis of pheromone detection in insects. In Comprehensive Molecular Insect Science; Gilbert, L.I., Iatro, K., Gill, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; Volume 3, pp. 753–804. [Google Scholar]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef]
- Oakeshott, J.G.; Claudianos, C.; Campbell, P.M.; Newcomb, R.D.; Russell, R.J. Biochemical genetics and genomics of insect esterases. In Comprehensive Molecular Insect Science; Gilbert, L.I., Ed.; Elsevier: Amsterdam, The Netherlands, 2005; Volume 5, pp. 309–381. [Google Scholar]
- Oakeshott, J.G.; Johnson, R.M.; Berenbaum, M.R.; Ranson, H.; Cristino, A.S.; Claudianos, C. Metabolic enzymes associated with xenobiotic and chemosensory responses in Nasonia vitripennis. Insect Mol. Biol. 2010, 19 (Suppl. 1), 147–163. [Google Scholar] [CrossRef] [Green Version]
- Vogt, R.G.; Riddiford, L.M. Pheromone binding and inactivation by moth antennae. Nature 1981, 293, 161–163. [Google Scholar] [CrossRef]
- Ishida, Y.; Leal, W.S. Cloning of putative odorant-degrading enzyme and integumental esterase cDNAs from the wild silkmoth, Antheraea polyphemus. Insect Biochem. Mol. Biol. 2002, 32, 1775–1780. [Google Scholar] [CrossRef]
- Klein, U. Sensillum-lymph proteins from antennal olfactory hairs of the moth Antheraea polyphemus (Saturniidae). Insect Biochem. 1987, 17, 1193–1204. [Google Scholar] [CrossRef]
- Vogt, R.G.; Riddiford, L.M. Scale esterase: A pheromone-degrading enzyme from scales of silk moth Antheraea polyphemus. J. Chem. Ecol. 1986, 12, 469–482. [Google Scholar] [CrossRef]
- Lu, F.G.; Fu, K.Y.; Li, Q.; Guo, W.C.; Ahmat, T.; Li, G.Q. Identification of carboxylesterase genes and their expression profiles in the Colorado potato beetle Leptinotarsa decemlineata treated with fipronil and cyhalothrin. Pestic. Biochem. Physiol. 2015, 122, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Field, L.M.; Devonshire, A.L. Structure and organization of amplicons containing the E4 esterase genes responsible for insecticide resistance in the aphid Myzus persicae (Sulzer). Biochem. J. 1997, 322, 867–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, L.M.; Williamson, M.S.; Moores, G.D.; Devonshire, A.L. Cloning and analysis of the esterase genes conferring insecticide resistance in the peach-potato aphid, Myzus persicae (Sulzer). Biochem. J. 1993, 294, 569–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida, Y.; Leal, W.S. Rapid inactivation of a moth pheromone. Proc. Natl. Acad. Sci. USA 2005, 102, 14075–14079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida, Y.; Leal, W.S. Chiral discrimination of the Japanese beetle sex pheromone and a behavioral antagonist by a pheromone-degrading enzyme. Proc. Natl. Acad. Sci. USA 2008, 105, 9076–9080. [Google Scholar] [CrossRef] [Green Version]
- Chertemps, T.; François, A.; Durand, N.; Rosell, G.; Dekker, T.; Lucas, P.; Maïbèche-Coisne, M. A carboxylesterase, Esterase-6, modulates sensory physiological and behavioral response dynamics to pheromone in Drosophila. BMC Biol. 2012, 10, 56. [Google Scholar] [CrossRef] [Green Version]
- He, P.; Zhang, Y.N.; Li, Z.Q.; Yang, K.; Zhu, J.Y.; Liu, S.J.; Dong, S.L. An antennae-enriched carboxylesterase from Spodoptera exigua displays degradation activity in both plant volatiles and female sex pheromones. Insect Mol. Biol. 2014, 23, 475–486. [Google Scholar] [CrossRef]
- Mackert, A.; do Nascimento, A.M.; Bitondi, M.M.; Hartfelder, K.; Simões, Z.L. Identification of a juvenile hormone esterase-like gene in the honey bee, Apis mellifera L.—Expression analysis and functional assays. Comp. Biochem. Physiol. B, Biochem. 2008, 150, 33–44. [Google Scholar] [CrossRef]
- Ward, V.K.; Bonning, B.C.; Huang, T.; Shiotsuki, T.; Griffeth, V.N.; Hammock, B.D. Analysis of the catalytic mechanism of juvenile hormone esterase by site-directed mutagenesis. Int. J. Biochem. 1992, 24, 1933–1941. [Google Scholar] [CrossRef]
- Kamita, S.G.; Hammock, B.D. Juvenile hormone esterase: Biochemistry and structure. J. Pestic Sci. 2010, 35, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Bai, H.; Ramaseshadri, P.; Palli, S.R. Identification and characterization of juvenile hormone esterase gene from the yellow fever mosquito, Aedes aegypti. Insect Biochem. Mol. Biol. 2007, 37, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Kamita, S.G.; Hinton, A.C.; Wheelock, C.E.; Wogulis, M.D.; Wilson, D.K.; Wolf, N.M.; Stok, J.E.; Hock, B.; Hammock, B.D. Juvenile hormone (JH) esterase: Why are you so JH specific? Insect Biochem. Mol. Biol. 2003, 33, 1261–1273. [Google Scholar] [CrossRef]
- Wogulis, M.; Wheelock, C.E.; Kamita, S.G.; Hinton, A.C.; Whetstone, P.A.; Hammock, B.D.; Wilson, D.K. Structural studies of a potent insect maturation inhibitor bound to the juvenile hormone esterase of Manduca sexta. Biochem. J. 2006, 45, 4045–4057. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, D.H.; Rane, R.V.; Younus, F.; Coppin, C.W.; Pandey, G.; Jackson, C.J.; Oakeshott, J.G. The molecular basis for the neofunctionalization of the juvenile hormone esterase duplication in Drosophila. Insect Biochem. Mol. Biol. 2019, 106, 10–18. [Google Scholar] [CrossRef]
- Crone, E.J.; Sutherland, T.D.; Campbell, P.M.; Coppin, C.W.; Russell, R.J.; Oakeshott, J.G. Only one esterase of Drosophila melanogaster is likely to degrade juvenile hormone in vivo. Insect Biochem. Mol. Biol. 2007, 37, 540–549. [Google Scholar] [CrossRef]
- Yan, L.; Yang, P.; Jiang, F.; Cui, N.; Ma, E.; Qiao, C.; Cui, F. Transcriptomic and phylogenetic analysis of Culex pipiens quinquefasciatus for three detoxification gene families. BMC Genom. 2012, 13, 609. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, M.M.; Auld, V.J. Evolution of clams (cholinesterase-like adhesion molecules): Structure and function during development. Front. Biosci. 2005, 10, 2177–2192. [Google Scholar] [CrossRef] [Green Version]
- Durand, N.; Chertemps, T.; Bozzolan, F.; Maïbèche, M. Expression and modulation of neuroligin and neurexin in the olfactory organ of the cotton leaf worm Spodoptera littoralis. Insect Sci. 2017, 24, 210–221. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.Y.; Lee, S.H.; Clark, J.M. A point mutation of acetylcholinesterase associated with azinphosmethyl resistance and reduced fitness in Colorado potato beetle. Pestic. Biochem. Physiol. 1996, 55, 100–108. [Google Scholar] [CrossRef]
- Baek, J.H.; Kim, J.I.; Lee, D.-W.; Chung, B.K.; Miyata, T.; Lee, S.H. Identification and characterization of ace1-type acetylcholinesterase likely associated with organophosphate resistance in Plutella xylostella. Pestic. Biochem. Physiol. 2005, 81, 164–175. [Google Scholar] [CrossRef]
- Fournier, D. Mutations of acetylcholinesterase which confer insecticide resistance in insect populations. Chem. Biol. Interact. 2005, 157–158, 257–261. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, Y.H.; Kwon, D.H.; Cha, D.J.; Kim, J.H. Mutation and duplication of arthropod acetylcholinesterase: Implications for pesticide resistance and tolerance. Pestic. Biochem. Physiol. 2015, 120, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, B.; Li, J.; Liu, M.; Liu, Z. Point mutations in acetylcholinesterase 1 associated with chlorpyrifos resistance in the brown planthopper, Nilaparvata lugens Stål. Insect Mol. Biol. 2017, 26, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.; Smith, J.; Roskams, A.J.; Auld, V.J. Neuroligin 3 is a vertebrate gliotactin expressed in the olfactory ensheathing glia, a growth-promoting class of macroglia. Glia 2001, 34, 151–164. [Google Scholar] [CrossRef] [PubMed]
- de la Escalera, S.; Bockamp, E.O.; Moya, F.; Piovant, M.; Jiménez, F. Characterization and gene cloning of neurotactin, a Drosophila transmembrane protein related to cholinesterases. EMBO J. 1990, 9, 3593–3601. [Google Scholar] [CrossRef]
- Nardini, M.; Dijkstra, B.W. α/β Hydrolase fold enzymes: The family keeps growing. Curr. Opin. Struct. Biol. 1999, 9, 732–737. [Google Scholar] [CrossRef]
- Jackson, C.J.; Liu, J.W.; Carr, P.D.; Younus, F.; Coppin, C.; Meirelles, T.; Lethier, M.; Pandey, G.; Ollis, D.L.; Russell, R.J.; et al. Structure and function of an insect α-carboxylesterase (αEsterase7) associated with insecticide resistance. Proc. Natl. Acad. Sci. USA 2013, 110, 10177–10182. [Google Scholar] [CrossRef] [Green Version]
- Harel, M.; Kryger, G.; Rosenberry, T.L.; Mallender, W.D.; Lewis, T.; Fletcher, R.J.; Guss, J.M.; Silman, I.; Sussman, J.L. Three-dimensional structures of Drosophila melanogaster acetylcholinesterase and of its complexes with two potent inhibitors. Preotein Sci. 2000, 9, 1063–1072. [Google Scholar] [CrossRef]
- Hopkins, D.H.; Fraser, N.J.; Mabbitt, P.D.; Carr, P.D.; Oakeshott, J.G.; Jackson, C.J. Structure of an insecticide sequestering carboxylesterase from the disease vector Culex quinquefasciatus: What makes an enzyme a good insecticide sponge? Biochem. J. 2017, 56, 5512–5525. [Google Scholar] [CrossRef]
- Kumar, K.; Mhetre, A.; Ratnaparkhi, G.S.; Kamat, S.S. A superfamily-wide activity atlas of Serine hydrolases in Drosophila melanogaster. Biochemistry 2021, 60, 1312–1324. [Google Scholar] [CrossRef]
- Han, Q.; Wong, D.M.; Robinson, H.; Ding, H.; Lam, P.C.H.; Totrov, M.M.; Carlier, P.R.; Li, J. Crystal structure of acetylcholinesterase catalytic subunits of the malaria vector Anopheles gambiae. Insect Sci. 2018, 25, 721–724. [Google Scholar] [CrossRef]
- Cheung, J.; Mahmood, A.; Kalathur, R.; Liu, L.; Carlier, P.R. Structure of the G119S mutant acetylcholinesterase of the malaria vector Anopheles gambiae reveals basis of insecticide resistance. Structure 2018, 26, 130–136.e2. [Google Scholar] [CrossRef]
- Strode, C.; Wondji, C.S.; David, J.P.; Hawkes, N.J.; Lumjuan, N.; Nelson, D.R.; Drane, D.R.; Karunaratne, S.H.; Hemingway, J.; Black, W.C.t.; et al. Genomic analysis of detoxification genes in the mosquito Aedes aegypti. Insect Biochem. Mol. Biol. 2008, 38, 113–123. [Google Scholar] [CrossRef]
- Xia, J.; Xu, H.; Yang, Z.; Pan, H.; Yang, X.; Guo, Z.; Yang, F.; Guo, L.; Sun, X.; Wang, S.; et al. Genome-wide analysis of carboxylesterases (COEs) in the whitefly, Bemisia tabaci (Gennadius). Int. J. Mol. Sci. 2019, 20, 4973. [Google Scholar] [CrossRef] [Green Version]
- Bailey, E.; Field, L.; Rawlings, C.; King, R.; Mohareb, F.; Pak, K.H.; Hughes, D.; Williamson, M.; Ganko, E.; Buer, B.; et al. A near-chromosome level genome assembly of the European hoverfly, Sphaerophoria rueppellii (Diptera: Syrphidae), provides comparative insights into insecticide resistance-related gene family evolution. BMC Genom. 2022, 23, 198. [Google Scholar] [CrossRef]
- Wu, Z.; Pu, X.; Shu, B.; Bin, S.; Lin, J. Transcriptome analysis of putative detoxification genes in the Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2020, 76, 3857–3870. [Google Scholar] [CrossRef]
- Ramsey, J.S.; Rider, D.S.; Walsh, T.K.; De Vos, M.; Gordon, K.H.; Ponnala, L.; Macmil, S.L.; Roe, B.A.; Jander, G. Comparative analysis of detoxification enzymes in Acyrthosiphon pisum and Myzus persicae. Insect Mol. Biol. 2010, 19 (Suppl. 2), 155–164. [Google Scholar] [CrossRef]
- Xue, J.; Zhou, X.; Zhang, C.-X.; Yu, L.-L.; Fan, H.-W.; Wang, Z.; Xu, H.-J.; Xi, Y.; Zhu, Z.-R.; Zhou, W.-W.; et al. Genomes of the rice pest brown planthopper and its endosymbionts reveal complex complementary contributions for host adaptation. Genom. Biol. 2014, 15, 521. [Google Scholar] [CrossRef] [Green Version]
- Mao, K.; Ren, Z.; Li, W.; Cai, T.; Qin, X.; Wan, H.; Jin, B.R.; He, S.; Li, J. Carboxylesterase genes in nitenpyram-resistant brown planthoppers, Nilaparvata lugens. Insect Sci. 2021, 28, 1049–1060. [Google Scholar] [CrossRef]
- Bailey, E.; Field, L.; Rawlings, C.; King, R.; Mohareb, F.; Pak, K.H.; Hughes, D.; Williamson, M.; Ganko, E.; Buer, B.; et al. A scaffold-level genome assembly of a minute pirate bug, Orius laevigatus (Hemiptera: Anthocoridae), and a comparative analysis of insecticide resistance-related gene families with hemipteran crop pests. BMC Genom. 2022, 23, 45. [Google Scholar] [CrossRef]
- Kwon, D.H.; Kim, J.H.; Kim, Y.H.; Yoon, K.S.; Clark, J.M.; Lee, S.H. Identification and characterization of an esterase involved in malathion resistance in the head louse Pediculus humanus capitis. Pestic. Biochem. Physiol. 2014, 112, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kang, J.S.; Min, J.S.; Yoon, K.S.; Strycharz, J.P.; Johnson, R.; Mittapalli, O.; Margam, V.M.; Sun, W.; Li, H.M.; et al. Decreased detoxification genes and genome size make the human body louse an efficient model to study xenobiotic metabolism. Insect Mol. Biol. 2010, 19, 599–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traverso, L.; Lavore, A.; Sierra, I.; Palacio, V.; Martinez-Barnetche, J.; Latorre-Estivalis, J.M.; Mougabure-Cueto, G.; Francini, F.; Lorenzo, M.G.; Rodríguez, M.H.; et al. Comparative and functional triatomine genomics reveals reductions and expansions in insecticide resistance-related gene families. PLoS. Negl. Trop. Dis. 2017, 11, e0005313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claudianos, C.; Ranson, H.; Johnson, R.M.; Biswas, S.; Schuler, M.A.; Berenbaum, M.R.; Feyereisen, R.; Oakeshott, J.G. A deficit of detoxification enzymes: Pesticide sensitivity and environmental response in the honeybee. Insect Mol. Biol. 2006, 15, 615–636. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.-Y.; Lu, C.; Li, W.-L.; Xiang, Z.-H.; Zhang, Z. Annotation and expression of carboxylesterases in the silkworm, Bombyx mori. BMC Genom. 2009, 10, 553. [Google Scholar] [CrossRef] [Green Version]
- You, M.; Yue, Z.; He, W.; Yang, X.; Yang, G.; Xie, M.; Zhan, D.; Baxter, S.W.; Vasseur, L.; Gurr, G.M.; et al. A heterozygous moth genome provides insights into herbivory and detoxification. Nat. Genet. 2013, 45, 220–225. [Google Scholar] [CrossRef] [Green Version]
- He, P.; Zhang, Y.-F.; Hong, D.-Y.; Wang, J.; Wang, X.-L.; Zuo, L.-H.; Tang, X.-F.; Xu, W.-M.; He, M. A reference gene set for sex pheromone biosynthesis and degradation genes from the diamondback moth, Plutella xylostella, based on genome and transcriptome digital gene expression analyses. BMC Genom. 2017, 18, 219. [Google Scholar] [CrossRef] [Green Version]
- Cheng, T.; Wu, J.; Wu, Y.; Chilukuri, R.V.; Huang, L.; Yamamoto, K.; Feng, L.; Li, W.; Chen, Z.; Guo, H.; et al. Genomic adaptation to polyphagy and insecticides in a major East Asian noctuid pest. Nat. Ecol. Evol. 2017, 1, 1747–1756. [Google Scholar] [CrossRef] [Green Version]
- Younus, F.; Fraser, N.J.; Coppin, C.W.; Liu, J.-W.; Correy, G.J.; Chertemps, T.; Pandey, G.; Maïbèche, M.; Jackson, C.J.; Oakeshott, J.G. Molecular basis for the behavioral effects of the odorant degrading enzyme Esterase 6 in Drosophila. Sci. Rep. 2017, 7, 46188. [Google Scholar] [CrossRef] [Green Version]
- Newcomb, R.D.; Campbell, P.M.; Ollis, D.L.; Cheah, E.; Russell, R.J.; Oakeshott, J.G. A single amino acid substitution converts a carboxylesterase to an organophosphorus hydrolase and confers insecticide resistance on a blowfly. Proc. Natl. Acad. Sci. USA 1997, 94, 7464–7468. [Google Scholar] [CrossRef] [Green Version]
- Comoletti, D.; Trobiani, L.; Chatonnet, A.; Bourne, Y.; Marchot, P. Comparative mapping of selected structural determinants on the extracellular domains of cholinesterase-like cell-adhesion molecules. Neuropharmacology 2021, 184, 108381. [Google Scholar] [CrossRef]
- Moural, T.W.; White, D.S.; Choy, C.J.; Kang, C.; Berkman, C.E. Crystal structure of phosphoserine blaC from mycobacterium tuberculosis inactivated by bis(benzoyl) phosphate. Int. J. Mol. Sci. 2019, 20, 3247. [Google Scholar] [CrossRef] [Green Version]
- Mabbitt, P.D.; Correy, G.J.; Meirelles, T.; Fraser, N.J.; Coote, M.L.; Jackson, C.J. Conformational disorganization within the active site of a recently evolved organophosphate hydrolase limits its catalytic efficiency. Biochem. J. 2016, 55, 1408–1417. [Google Scholar] [CrossRef]
- Newcomb, R.D.; Gleeson, D.M.; Yong, C.G.; Russell, R.J.; Oakeshott, J.G. Multiple mutations and gene duplications conferring organophosphorus insecticide resistance have been selected at the Rop-1 locus of the sheep blowfly, Lucilia cuprina. J. Mol. Evol. 2005, 60, 207–220. [Google Scholar] [CrossRef]
- Levot, G.W. Resistance and the control of sheep ectoparasites. Int. J. Parisitol. 1995, 25, 1355–1362. [Google Scholar] [CrossRef]
- Toutant, J.P. Insect acetylcholinesterase: Catalytic properties, tissue distribution and molecular forms. Prog. Neurobiol. 1989, 32, 423–446. [Google Scholar] [CrossRef]
- Vogt, R.G.; Riddiford, L.M.; Prestwich, G.D. Kinetic properties of a sex pheromone-degrading enzyme: The sensillar esterase of Antheraea polyphemus. Proc. Natl. Acad. Sci. USA 1985, 82, 8827–8831. [Google Scholar] [CrossRef] [Green Version]
- Ranson, H.; Claudianos, C.; Ortelli, F.; Abgrall, C.; Hemingway, J.; Sharakhova, M.V.; Unger, M.F.; Collins, F.H.; Feyereisen, R. Evolution of supergene families associated with insecticide resistance. N. Y. Sci. J. 2002, 298, 179–181. [Google Scholar] [CrossRef]
- Soderlund, D.M. Molecular mechanisms of pyrethroid insecticide neurotoxicity: Recent advances. Arch. Toxicol. 2012, 86, 165–181. [Google Scholar] [CrossRef] [Green Version]
- Feyereisen, R.; Dermauw, W.; Van Leeuwen, T. Genotype to phenotype, the molecular and physiological dimensions of resistance in arthropods. Pestic. Biochem. Physiol. 2015, 121, 61–77. [Google Scholar] [CrossRef]
- Cui, F.; Li, M.X.; Chang, H.J.; Mao, Y.; Zhang, H.Y.; Lu, L.X.; Yan, S.G.; Lang, M.L.; Liu, L.; Qiao, C.L. Carboxylesterase-mediated insecticide resistance: Quantitative increase induces broader metabolic resistance than qualitative change. Pestic. Biochem. Physiol. 2015, 121, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Mutero, A.; Pralavorio, M.; Bride, J.M.; Fournier, D. Resistance-associated point mutations in insecticide-insensitive acetylcholinesterase. Proc. Natl. Acad. Sci. USA 1994, 91, 5922–5926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weill, M.; Malcolm, C.; Chandre, F.; Mogensen, K.; Berthomieu, A.; Marquine, M.; Raymond, M. The unique mutation in ace-1 giving high insecticide resistance is easily detectable in mosquito vectors. Insect Mol. Biol. 2004, 13, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Field, L.M.; Blackman, R.L.; Tyler-Smith, C.; Devonshire, A.L. Relationship between amount of esterase and gene copy number in insecticide-resistant Myzus persicae (Sulzer). Biochem. J. 1999, 339 (Pt 3), 737–742. [Google Scholar] [CrossRef]
- Devonshire, A.L.; Moores, G.D. A carboxylesterase with broad substrate specificity causes organophosphorus, carbamate and pyrethroid resistance in peach-potato aphids (Myzus persicae). Pestic. Biochem. Physiol. 1982, 18, 235–246. [Google Scholar] [CrossRef]
- Devonshire, A.L.; Field, L.M.; Foster, S.P.; Moores, G.D.; Williamson, M.S.; Blackman, R.L. The evolution of insecticide resistance in the peach–potato aphid, Myzus persicae. Philos. Trans. R. Soc. 1998, 353, 1677–1684. [Google Scholar] [CrossRef] [Green Version]
- Bass, C.; Puinean, A.M.; Zimmer, C.T.; Denholm, I.; Field, L.M.; Foster, S.P.; Gutbrod, O.; Nauen, R.; Slater, R.; Williamson, M.S. The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem. Mol. Biol. 2014, 51, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Small, G.J.; Hemingway, J. Molecular characterization of the amplified carboxylesterase gene associated with organophosphorus insecticide resistance in the brown planthopper, Nilaparvata lugens. Insect Mol. Biol. 2000, 9, 647–653. [Google Scholar] [CrossRef]
- Vontas, J.G.; Small, G.J.; Hemingway, J. Comparison of esterase gene amplification, gene expression and esterase activity in insecticide susceptible and resistant strains of the brown planthopper, Nilaparvata lugens (Stål). Insect Mol. Biol. 2000, 9, 655–660. [Google Scholar] [CrossRef]
- Claudianos, C.; Russell, R.J.; Oakeshott, J.G. The same amino acid substitution in orthologous esterases confers organophosphate resistance on the house fly and a blowfly. Insect Biochem. Mol. Biol. 1999, 29, 675–686. [Google Scholar] [CrossRef]
- Campbell, P.M.; Newcomb, R.D.; Russell, R.J.; Oakeshott, J.G. Two different amino acid substitutions in the ali-esterase, E3, confer alternative types of organophosphorus insecticide resistance in the sheep blowfly, Lucilia cuprina. Insect Biochem. Mol. Biol. 1998, 28, 139–150. [Google Scholar] [CrossRef]
- Tang, B.; Dai, W.; Qi, L.; Du, S.; Zhang, C. Functional characterization of an α-esterase gene associated with malathion detoxification in Bradysia odoriphaga. J. Agric. Food Chem. 2020, 68, 6076–6083. [Google Scholar] [CrossRef]
- Whyard, S.; Downe, A.E.; Walker, V.K. Characterization of a novel esterase conferring insecticide resistance in the mosquito Culex tarsalis. Arch. Insect Biochem. Physiol. 1995, 29, 329–342. [Google Scholar] [CrossRef]
- Feng, X.; Liu, N. Functional analyses of house fly carboxylesterases involved in insecticide resistance. Front. Physiol. 2020, 11, 595009. [Google Scholar] [CrossRef]
- Karunaratne, S.H.P.P.; Damayanthi, B.T.; Fareena, M.H.J.; Imbuldeniya, V.; Hemingway, J. Insecticide resistance in the tropical bedbug Cimex hemipterus. Pestic. Biochem. Physiol. 2007, 88, 102–107. [Google Scholar] [CrossRef]
- Punchihewa, R.; de Silva, W.; Weeraratne, T.C.; Karunaratne, S. Insecticide resistance mechanisms with novel ‘kdr’ type gene mutations in the tropical bed bug Cimex hemipterus. Parasites Vectors 2019, 12, 310. [Google Scholar] [CrossRef]
- Wei, P.; Demaeght, P.; De Schutter, K.; Grigoraki, L.; Labropoulou, V.; Riga, M.; Vontas, J.; Nauen, R.; Dermauw, W.; Van Leeuwen, T. Overexpression of an alternative allele of carboxyl/choline esterase 4 (CCE04) of Tetranychus urticae is associated with high levels of resistance to the keto-enol acaricide spirodiclofen. Pest. Manag. Sci. 2020, 76, 1142–1153. [Google Scholar] [CrossRef]
- Li, R.; Zhu, B.; Liang, P.; Gao, X. Identification of carboxylesterase genes contributing to multi-insecticide resistance in Plutella xylostella (L.). Entomol. Gen. 2022, 42, 967–976. [Google Scholar] [CrossRef]
- Li, R.; Zhu, B.; Hu, X.P.; Shi, X.Y.; Qi, L.L.; Liang, P.; Gao, X.W. Overexpression of PxαE14 contributing to detoxification of multiple insecticides in Plutella xylostella (L.). J. Agric. Food Chem. 2022, 70, 5794–5804. [Google Scholar] [CrossRef]
- Li, F.; Zhaojun Han, Z. Mutations in acetylcholinesterase associated with insecticide resistance in the cotton aphid, Aphis gossypii Glover. Insect Biochem. Mol. Biol. 2004, 34, 397–405. [Google Scholar] [CrossRef]
- Devonshire, A.L.; Moores, G.D. Different forms of insensitive acetylcholinesterase in insecticide-resistant house flies (Musca domestica). Pestic. Biochem. Physiol. 1984, 21, 336–340. [Google Scholar] [CrossRef]
- Ilias, A.; Vontas, J.; Tsagkarakou, A. Global distribution and origin of target site insecticide resistance mutations in Tetranychus urticae. Insect Biochem. Mol. Biol. 2014, 48, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Margolies, D.C.; Zhu, K.Y.; Buschman, L.L. Host plant-induced changes in detoxification enzymes and susceptibility to pesticides in the twospotted spider mite (Acari: Tetranychidae). J. Econ. Entomol. 2001, 94, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, F.; He, Q.; He, G. Molecular dynamics of detoxification and toxin-tolerance genes in brown planthopper (Nilaparvata lugens Stål., Homoptera: Delphacidae) feeding on resistant rice plants. Arch. Insect Biochem. Physiol. 2005, 59, 59–66. [Google Scholar] [CrossRef]
- Karuppaiah, V.; Srivastava, C.; Subramanian, S. Effect of host plants on insecticide susceptibility and detoxification enzymes activity in Spodoptera litura Fabricius (Noctuidae: Lepidoptera). Proc. Natl. Acad. Sci. India B-Biol. Sci. 2016, 86, 715–721. [Google Scholar] [CrossRef]
- Liu, B.; Coy, M.; Wang, J.J.; Stelinski, L.L. The effect of host plant species on the detoxifying enzymes of the Asian citrus psyllid, Diaphorina citri (Hemiptera: Liviidae). Fla. Entomol. 2015, 98, 997–999. [Google Scholar] [CrossRef]
- Xu, H.X.; Hong, Y.; Zhang, M.Z.; Wang, Y.L.; Liu, S.S.; Wang, X.W. Transcriptional responses of invasive and indigenous whiteflies to different host plants reveal their disparate capacity of adaptation. Sci. Rep. 2015, 5, 10774. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Zhu, F.; Sun, Q.; Zhou, X. Editorial: Mechanisms and strategies of arthropod adaptation to the chemical environment. Front. Physiol. 2022, 13, 889757. [Google Scholar] [CrossRef]
- Lu, K.; Song, Y.; Zeng, R. The role of cytochrome P450-mediated detoxification in insect adaptation to xenobiotics. Curr. Opin. Insect. Sci. 2021, 43, 103–107. [Google Scholar] [CrossRef]
- Zhu, F.; Moural, T.W.; Nelson, D.R.; Palli, S.R. A specialist herbivore pest adaptation to xenobiotics through up-regulation of multiple Cytochrome P450s. Sci. Rep. 2016, 6, 20421. [Google Scholar] [CrossRef] [Green Version]
- Scriber, J.M.; Lindroth, R.L.; Nitao, J.K. Toxic phenolic glycosides from populus: Physiological adaptations of the Western North American tiger awallowtail butterfly, Papilio rutulus (Lepidoptera: Papilionidae). Gt. Lakes Entomol. 1991, 24, 173–180. [Google Scholar]
- Lindroth, R.L.; Weisbrod, A.V. Genetic-variation in response of the Gypsy-moth to aspen phenolic glycosides. Biochem. Syst. Ecol. 1991, 19, 97–103. [Google Scholar] [CrossRef]
- Hemming, J.D.C.; Lindroth, R.L. Effects of phenolic glycosides and protein on gypsy moth (Lepidoptera: Lymantriidae) and forest tent caterpillar (Lepidoptera: Lasiocampidae) performance and detoxication activities. Environ. Entomol. 2000, 29, 1108–1115. [Google Scholar] [CrossRef]
- Cai, Q.N.; Han, Y.; Cao, Y.Z.; Hu, Y.; Zhao, X.; Bi, J.L. Detoxification of gramine by the cereal aphid Sitobion avenae. J. Chem. Ecol. 2009, 35, 320–325. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.-Y.; Zhang, Y.; Wang, H.-Y.; Xia, X.-M.; Liu, T.-X. Influence of three diets on susceptibility of selected insecticides and activities of detoxification esterases of Helicoverpa assulta (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2010, 96, 51–55. [Google Scholar] [CrossRef]
- Xue, M.; Pang, Y.H.; Li, Q.L.; Liu, T.X. Effects of four host plants on susceptibility of Spodoptera litura (Lepidoptera: Noctuidae) larvae to five insecticides and activities of detoxification esterases. Pest. Manag. Sci. 2010, 66, 1273–1279. [Google Scholar] [CrossRef]
- Ghumare, S.S.; Mukherjee, S.N.; Sharma, R.N. Effect of rutin on the neonate sensitivity, dietary utilization and mid-gut carboxylesterase activity of Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae). Anim. Sci. J. 1989, 98, 399–404. [Google Scholar]
- Song, L.-M.; Jiang, X.; Wang, X.-M.; Li, J.-D.; Zhu, F.; Tu, X.-B.; Zhang, Z.-H.; Ban, L.-P. Male tarsi specific odorant-binding proteins in the diving beetle Cybister japonicus Sharp. Sci. Rep. 2016, 6, 31848. [Google Scholar] [CrossRef] [Green Version]
- Homberg, U.; Christensen, T.A.; Hildebrand, J.G. Structure and function of the deutocerebrum in insects. Annu. Rev. Entomol. 1989, 34, 477–501. [Google Scholar] [CrossRef]
- Wu, H.; Liu, Y.; Shi, X.; Zhang, X.; Ye, C.; Zhu, K.Y.; Zhu, F.; Zhang, J.; Ma, E. Transcriptome analysis of antennal cytochrome P450s and their transcriptional responses to plant and locust volatiles in Locusta migratoria. Int. J. Biol. Macromol. 2020, 149, 741–753. [Google Scholar] [CrossRef]
- Younus, F.; Chertemps, T.; Pearce, S.L.; Pandey, G.; Bozzolan, F.; Coppin, C.W.; Russell, R.J.; Maibeche-Coisne, M.; Oakeshott, J.G. Identification of candidate odorant degrading gene/enzyme systems in the antennal transcriptome of Drosophila melanogaster. Insect Biochem. Mol. Biol. 2014, 53, 30–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, C.; Chertemps, T.; Maïbèche, M. Diversity of biotransformation enzymes in insect antennae: Possible roles in odorant inactivation and xenobiotic processing. In Olfactory Concepts of Insect Control—Alternative to Insecticides; Picimbon, J.-F., Ed.; Springer International Publishing: Cham, Switzerland, 2019; Volume 2, pp. 115–145. [Google Scholar]
- Durand, N.; Carot-Sans, G.; Chertemps, T.; Bozzolan, F.; Party, V.; Renou, M.; Debernard, S.; Rosell, G.; Maïbèche-Coisne, M. Characterization of an antennal carboxylesterase from the pest moth Spodoptera littoralis degrading a host plant odorant. PLoS ONE 2010, 5, e15026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand, N.; Carot-Sans, G.; Bozzolan, F.; Rosell, G.; Siaussat, D.; Debernard, S.; Chertemps, T.; Maïbèche-Coisne, M. Degradation of pheromone and plant volatile components by a same odorant-degrading enzyme in the cotton leafworm, Spodoptera littoralis. PLoS ONE 2011, 6, 29147. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Mang, D.Z.; Wang, H.; Wang, M.M.; Ma, Y.F.; Wang, J.; Chen, G.L.; Zhang, F.; He, M. Molecular characterization and functional analysis of a novel candidate of cuticle carboxylesterase in Spodoptera exigua degradating sex pheromones and plant volatile esters. Pestic. Biochem. Physiol. 2020, 163, 227–234. [Google Scholar] [CrossRef]
- Chertemps, T.; Younus, F.; Steiner, C.; Durand, N.; Coppin, C.W.; Pandey, G.; Oakeshott, J.G.; Maïbèche, M. An antennal carboxylesterase from Drosophila melanogaster, esterase 6, is a candidate odorant-degrading enzyme toward food odorants. Front. Physiol. 2015, 6, 315. [Google Scholar] [CrossRef] [Green Version]
- Steiner, C.; Bozzolan, F.; Montagné, N.; Maïbèche, M.; Chertemps, T. Neofunctionalization of “Juvenile Hormone Esterase Duplication” in Drosophila as an odorant-degrading enzyme towards food odorants. Sci. Rep. 2017, 7, 12629. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.M.; Long, G.J.; Guo, H.; Liu, X.Z.; Wang, H.; Dewer, Y.; Li, Z.Q.; Liu, K.; Zhang, Q.L.; Ma, Y.F.; et al. Two carboxylesterase genes in Plutella xylostella associated with sex pheromones and plant volatiles degradation. Pest Manag. Sci. 2021, 77, 2737–2746. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruse, C.; Moural, T.W.; Zhu, F. Dynamic Roles of Insect Carboxyl/Cholinesterases in Chemical Adaptation. Insects 2023, 14, 194. https://doi.org/10.3390/insects14020194
Cruse C, Moural TW, Zhu F. Dynamic Roles of Insect Carboxyl/Cholinesterases in Chemical Adaptation. Insects. 2023; 14(2):194. https://doi.org/10.3390/insects14020194
Chicago/Turabian StyleCruse, Casey, Timothy Walter Moural, and Fang Zhu. 2023. "Dynamic Roles of Insect Carboxyl/Cholinesterases in Chemical Adaptation" Insects 14, no. 2: 194. https://doi.org/10.3390/insects14020194