

Hemp Waste as a Substrate for Hermetia illucens (L.) (Diptera: Stratiomyidae) and Tenebrio molitor L. (Coleoptera: Tenebrionidae) Rearing

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Black Soldier Fly Colony
2.2. Yellow Mealworms Colony
2.3. Hemp Material
2.4. Black Soldier Fly Larvae Feeding Experiment
2.5. Mealworm Feeding Experiment
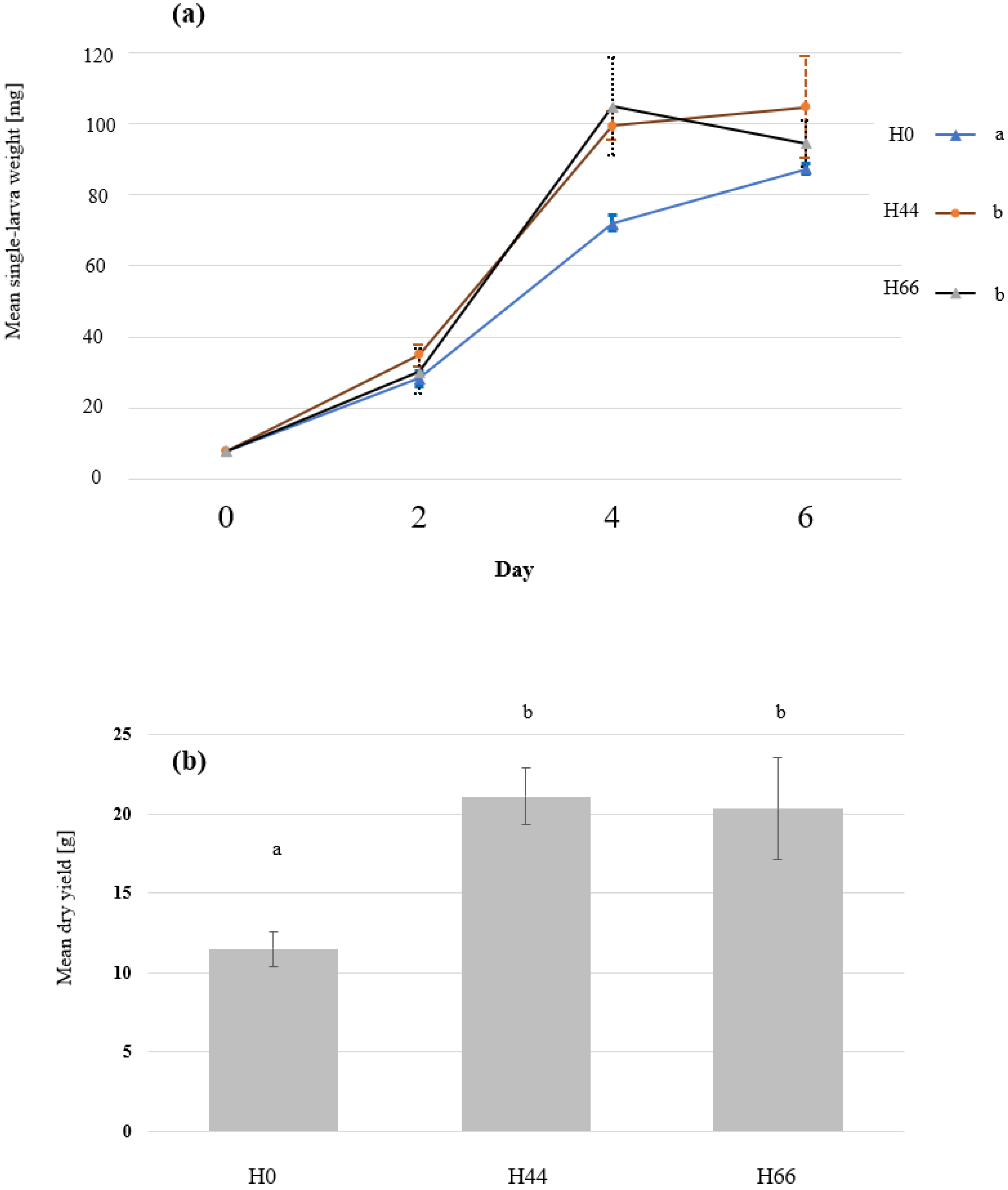
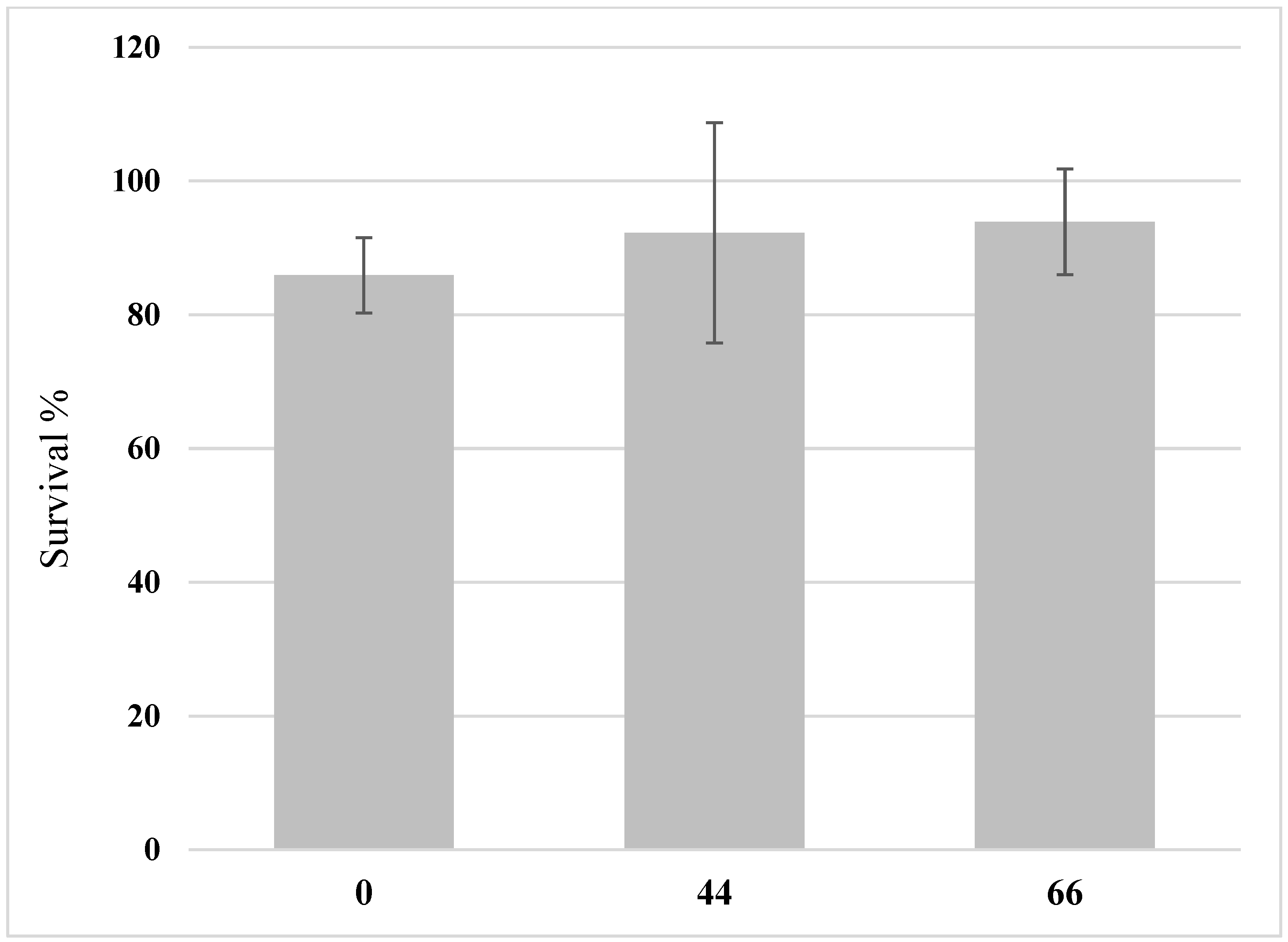
2.6. Determination of Growth and Biomass Parameters
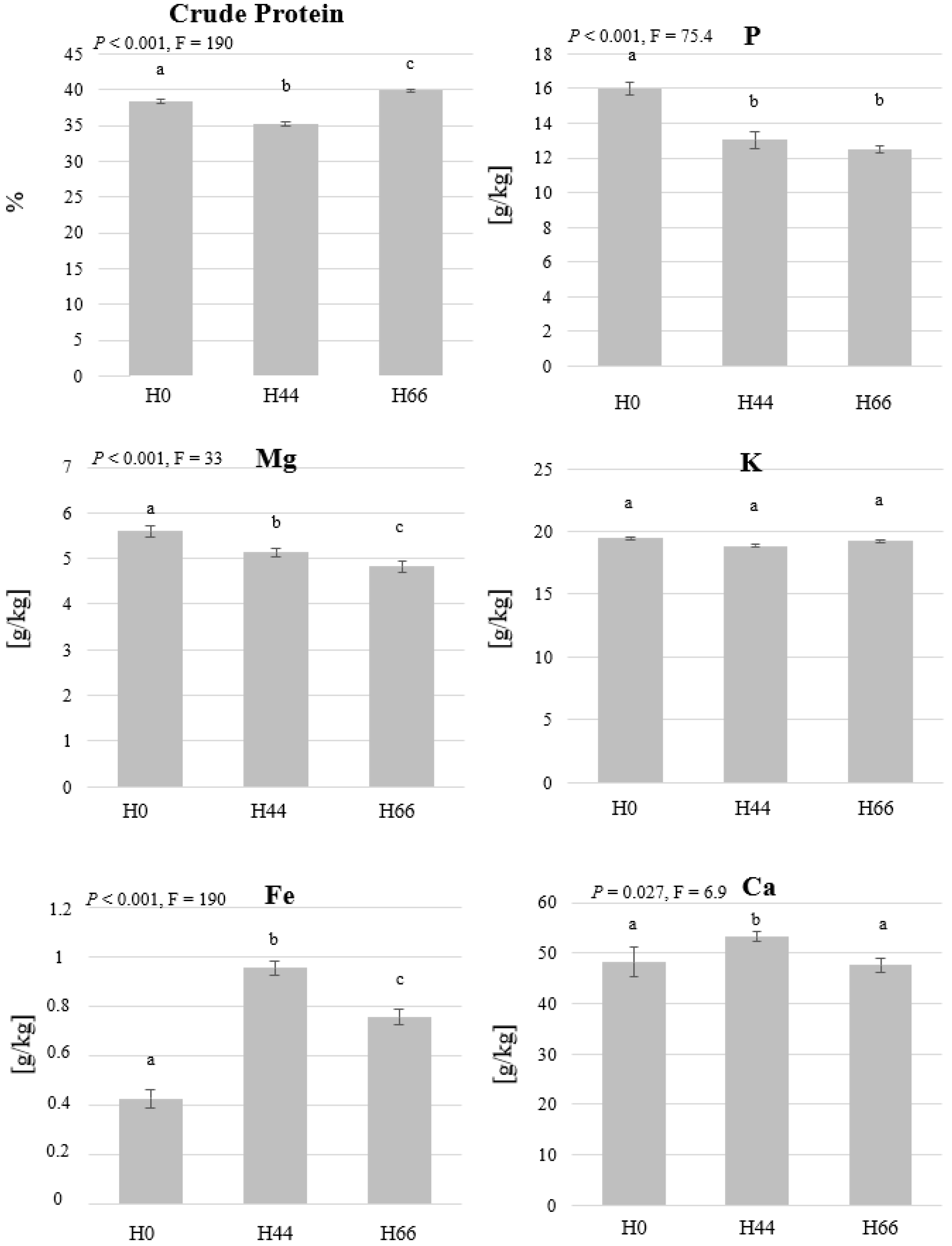
2.7. Elemental Analysis and Protein Quantification
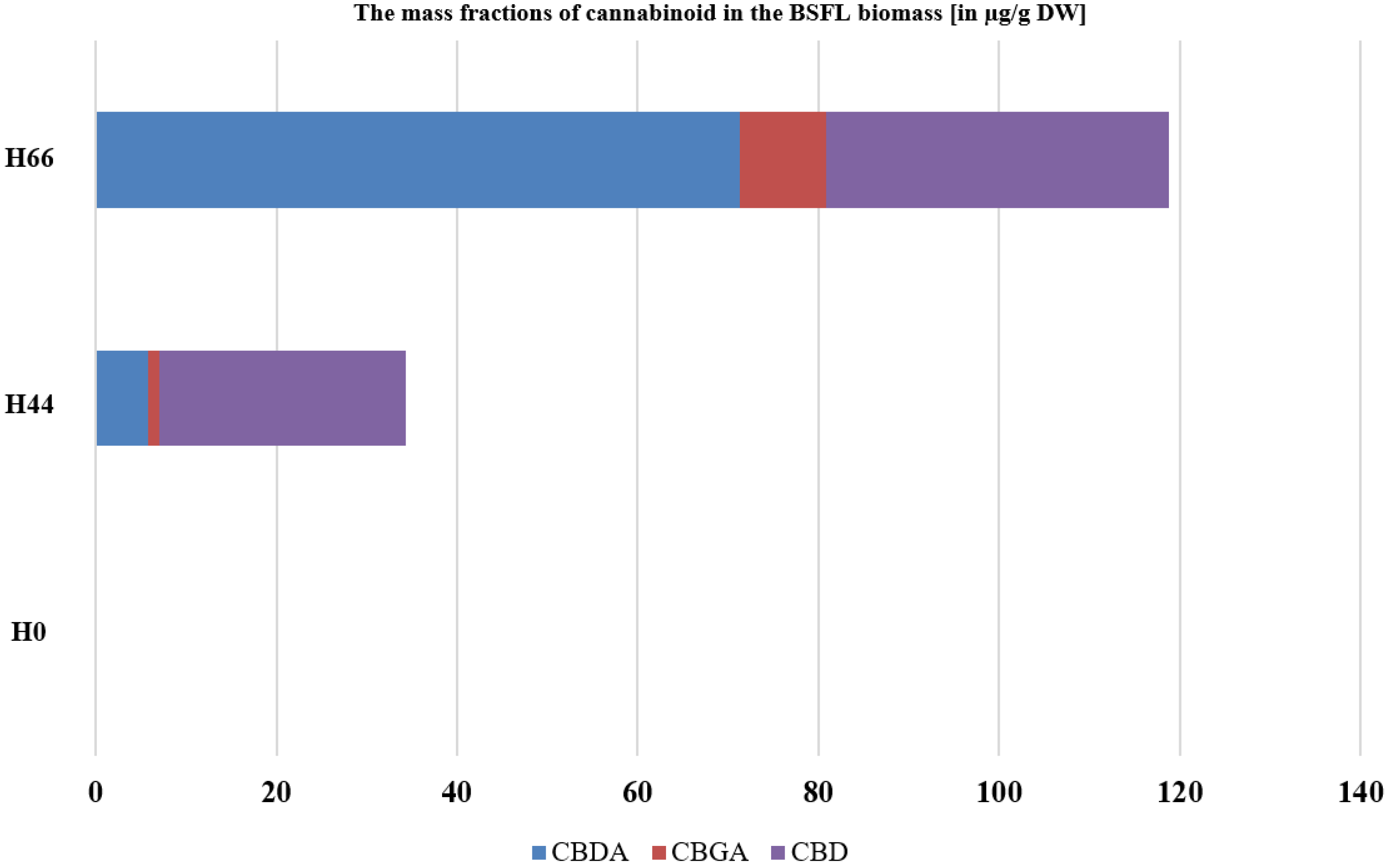
2.8. The Detection and Quantification of Cannabinoids
2.9. Statistical Analyses
3. Results
3.1. BSFL Feeding Experiment
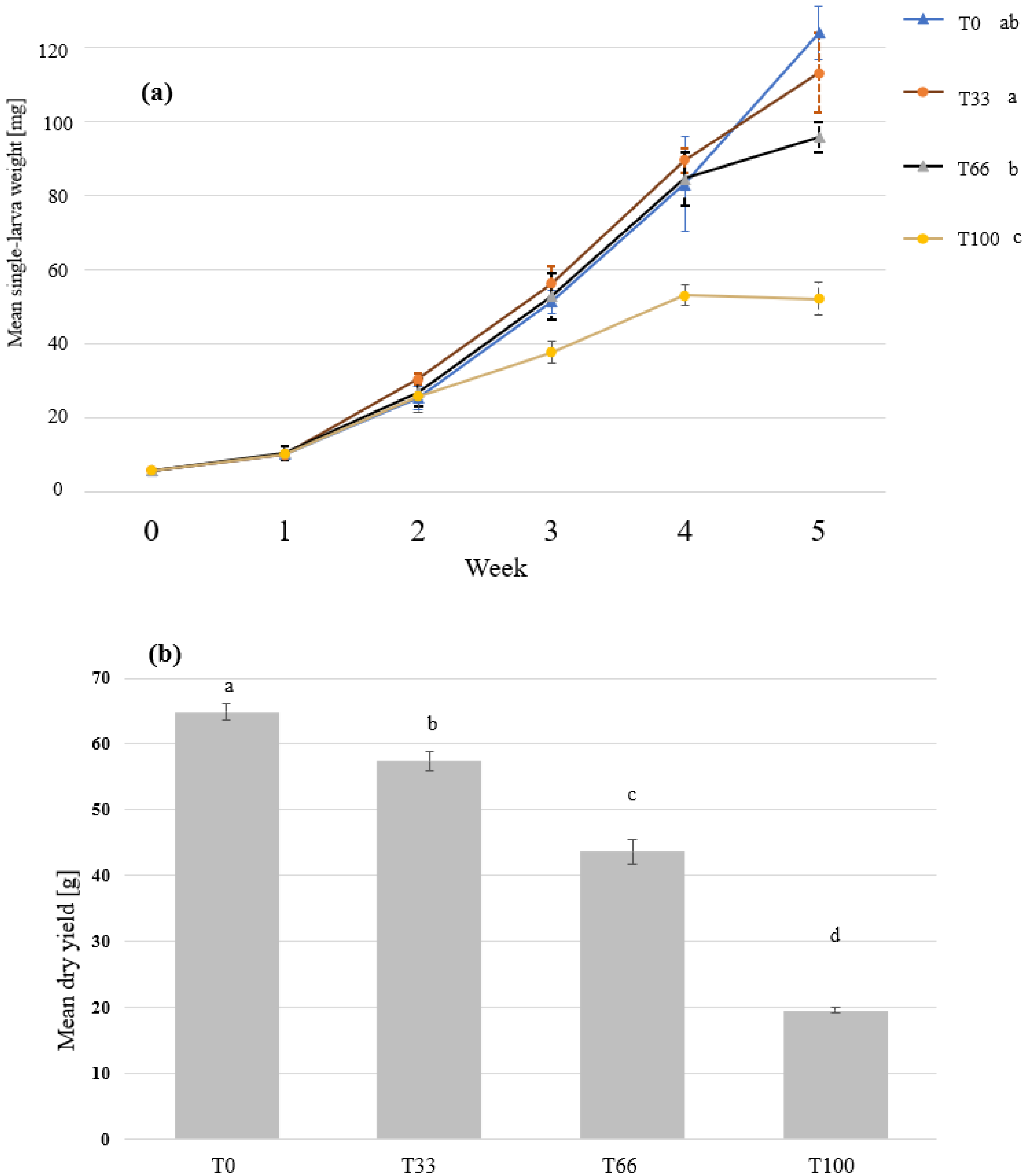
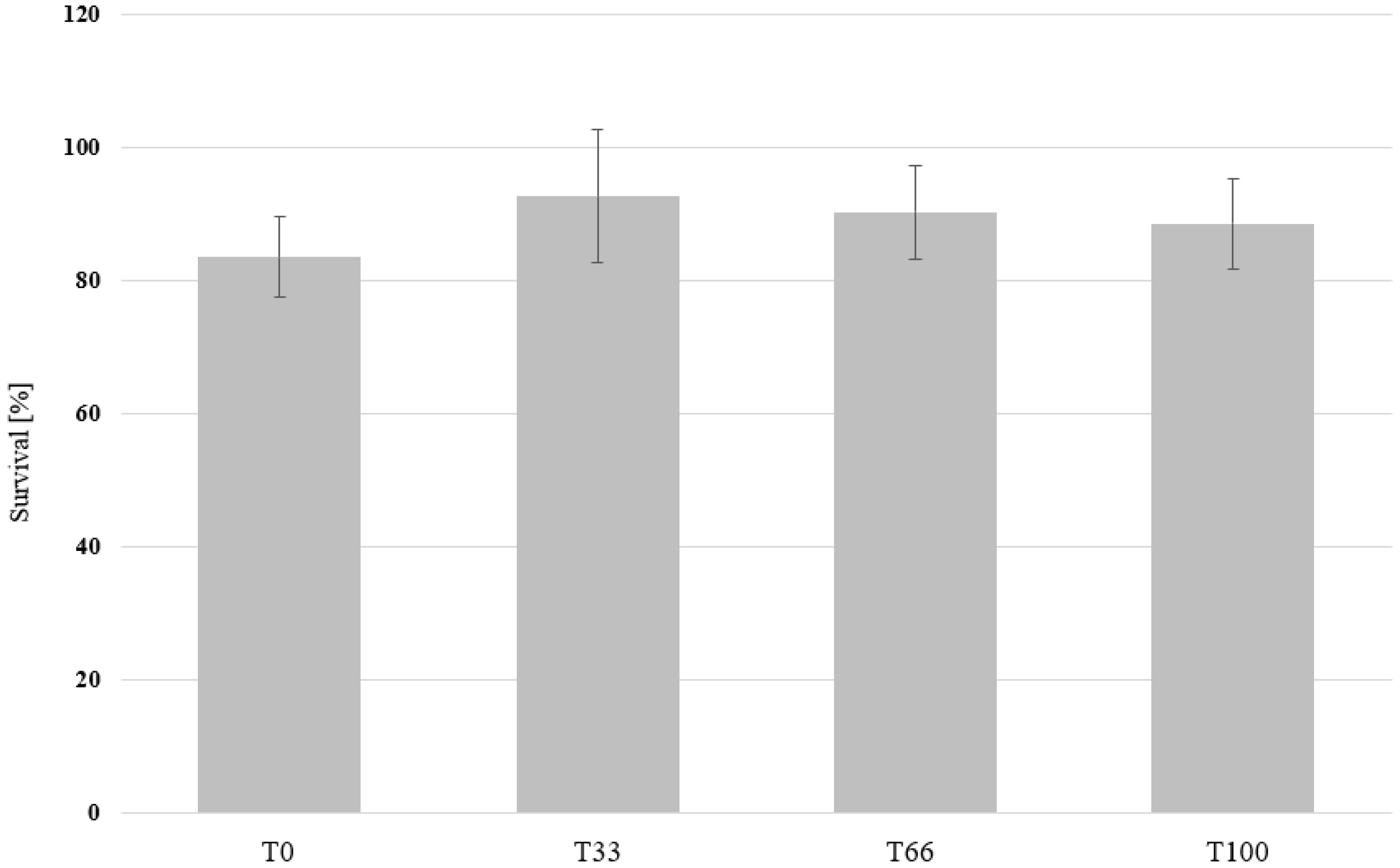
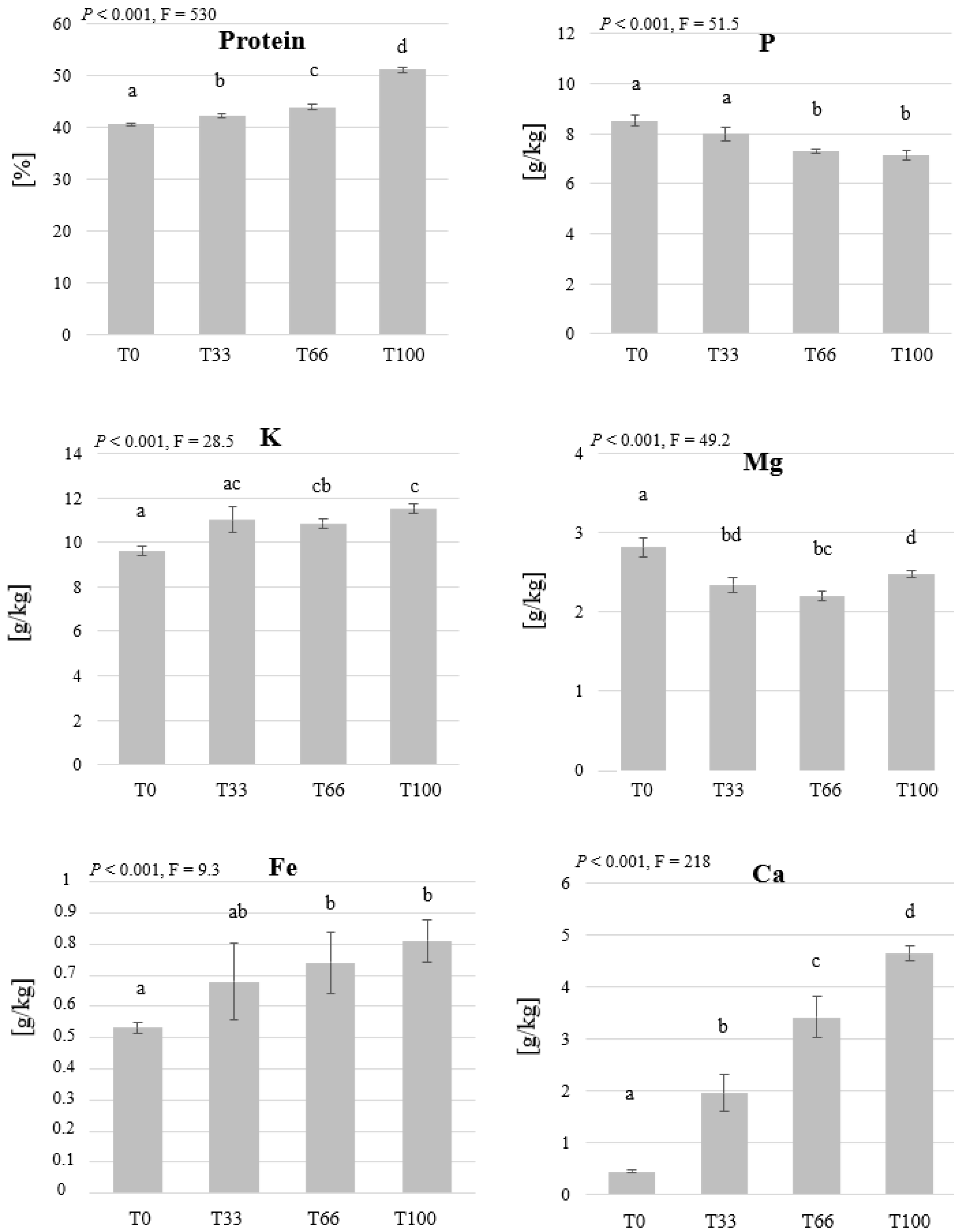
3.2. Mealworm Feeding Experiment
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Small, E. Classification of Cannabis sativa L. in relation to agricultural, biotechnological, medical and recreational utilization. In Cannabis sativa L.—Botany and Biotechnology; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–62. [Google Scholar]
- Farag, S.; Kayser, O. The cannabis plant: Botanical aspects. In Handbook of Cannabis and Related Pathologies; Elsevier: Amsterdam, The Netherlands, 2017; pp. 3–12. [Google Scholar]
- Small, E. Evolution and classification of Cannabis sativa (marijuana, hemp) in relation to human utilization. Bot. Rev. 2015, 81, 189–294. [Google Scholar] [CrossRef]
- Fiani, B.; Sarhadi, K.J.; Soula, M.; Zafar, A.; Quadri, S.A. Current application of cannabidiol (CBD) in the management and treatment of neurological disorders. Neurol. Sci. 2020, 41, 3085–3098. [Google Scholar] [CrossRef]
- Commission, E. Hemp Production in the EU. Available online: https://agriculture.ec.europa.eu/farming/crop-productions-and-plant-based-products/hemp_en (accessed on 4 October 2022).
- Apostol, L. Studies on using hemp seed as functional ingredient in the production of functional food products. J. Ecoagritourism 2017, 13, 12–17. [Google Scholar]
- Bouloc, P. Hemp: Industrial Production and Uses; CABI: Wallingford, UK, 2013. [Google Scholar]
- Pecoraro, M.T.; Mellinas, C.; Piccolella, S.; Garrigos, M.C.; Pacifico, S. Hemp Stem Epidermis and Cuticle: From Waste to Starter in Bio-Based Material Development. Polymers 2022, 14, 2816. [Google Scholar] [CrossRef] [PubMed]
- Paini, J.; Benedetti, V.; Ail, S.S.; Castaldi, M.J.; Baratieri, M.; Patuzzi, F. Valorization of wastes from the food production industry: A review towards an integrated agri-food processing biorefinery. Waste Biomass Valorization 2022, 13, 31–50. [Google Scholar] [CrossRef]
- Ubando, A.T.; Felix, C.B.; Chen, W.-H. Biorefineries in circular bioeconomy: A comprehensive review. Bioresour. Technol. 2020, 299, 122585. [Google Scholar] [CrossRef]
- Rumbos, C.; Oonincx, D.; Karapanagiotidis, I.; Vrontaki, M.; Gourgouta, M.; Asimaki, A.; Mente, E.; Athanassiou, C. Agricultural by-products from Greece as feed for yellow mealworm larvae: Circular economy at a local level. J. Insects Food Feed 2022, 8, 9–22. [Google Scholar] [CrossRef]
- Sangiorgio, P.; Verardi, A.; Dimatteo, S.; Spagnoletta, A.; Moliterni, S.; Errico, S. Valorisation of agri-food waste and mealworms rearing residues for improving the sustainability of Tenebrio molitor industrial production. J. Insects Food Feed 2022, 8, 509–524. [Google Scholar] [CrossRef]
- Manurung, R.; Supriatna, A.; Esyanthi, R.R.; Putra, R.E. Bioconversion of rice straw waste by black soldier fly larvae (Hermetia illucens L.): Optimal feed rate for biomass production. J. Entomol. Zool. Stud. 2016, 4, 1036–1041. [Google Scholar]
- Scieuzo, C.; Franco, A.; Salvia, R.; Triunfo, M.; Addeo, N.F.; Vozzo, S.; Piccolo, G.; Bovera, F.; Ritieni, A.; Francia, A.D. Enhancement of fruit byproducts through bioconversion by Hermetia illucens (Diptera: Stratiomyidae). Insect Sci. 2022. [Google Scholar] [CrossRef] [PubMed]
- Scala, A.; Cammack, J.A.; Salvia, R.; Scieuzo, C.; Franco, A.; Bufo, S.A.; Tomberlin, J.K.; Falabella, P. Rearing substrate impacts growth and macronutrient composition of Hermetia illucens (L.) (Diptera: Stratiomyidae) larvae produced at an industrial scale. Sci. Rep. 2020, 10, 19448. [Google Scholar] [CrossRef] [PubMed]
- Benzertiha, A.; Kierończyk, B.; Rawski, M.; Mikołajczak, Z.; Urbański, A.; Nogowski, L.; Józefiak, D. Insect fat in animal nutrition—A review. Ann. Anim. Sci. 2020, 20, 1217–1240. [Google Scholar] [CrossRef]
- Gasco, L.; Biasato, I.; Dabbou, S.; Schiavone, A.; Gai, F. Animals fed insect-based diets: State-of-the-art on digestibility, performance and product quality. Animals 2019, 9, 170. [Google Scholar] [CrossRef]
- EFSA Panel on Nutrition; Novel Foods and Food Allergens (NDA); Turck, D.; Bohn, T.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; et al. Safety of frozen and dried formulations from whole yellow mealworm (Tenebrio molitor larva) as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06778. [Google Scholar] [CrossRef] [PubMed]
- Alagappan, S.; Rowland, D.; Barwell, R.; Mantilla, S.; Mikkelsen, D.; James, P.; Yarger, O.; Hoffman, L. Legislative landscape of black soldier fly (Hermetia illucens) as feed. J. Insects Food Feed 2022, 8, 343–355. [Google Scholar] [CrossRef]
- Melenchón, F.; Larrán, A.; De Mercado, E.; Hidalgo, M.; Cardenete, G.; Barroso, F.; Fabrikov, D.; Lourenço, H.; Pessoa, M.; Tomás-Almenar, C. Potential use of black soldier fly (Hermetia illucens) and mealworm (Tenebrio molitor) insectmeals in diets for rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 2021, 27, 491–505. [Google Scholar] [CrossRef]
- Llagostera, P.F.; Kallas, Z.; Reig, L.; De Gea, D.A. The use of insect meal as a sustainable feeding alternative in aquaculture: Current situation, Spanish consumers’ perceptions and willingness to pay. J. Clean. Prod. 2019, 229, 10–21. [Google Scholar] [CrossRef]
- Oddon, S.B.; Biasato, I.; Imarisio, A.; Pipan, M.; Dekleva, D.; Colombino, E.; Capucchio, M.T.; Meneguz, M.; Bergagna, S.; Barbero, R. Black Soldier Fly and Yellow Mealworm live larvae for broiler chickens: Effects on bird performance and health status. J. Anim. Physiol. Anim. Nutr. 2021, 105, 10–18. [Google Scholar] [CrossRef] [PubMed]
- DiGiacomo, K.; Leury, B. Insect meal: A future source of protein feed for pigs? Animal 2019, 13, 3022–3030. [Google Scholar] [CrossRef]
- Hong, J.; Han, T.; Kim, Y.Y. Mealworm (Tenebrio molitor Larvae) as an alternative protein source for monogastric animal: A review. Animals 2020, 10, 2068. [Google Scholar] [CrossRef]
- Verheyen, G.R.; Ooms, T.; Vogels, L.; Vreysen, S.; Bovy, A.; Van Miert, S.; Meersman, F. Insects as an alternative source for the production of fats for cosmetics. J. Cosmet. Sci. 2018, 69, 187–202. [Google Scholar]
- Li, Q.; Zheng, L.; Cai, H.; Garza, E.; Yu, Z.; Zhou, S. From organic waste to biodiesel: Black soldier fly, Hermetia illucens, makes it feasible. Fuel 2011, 90, 1545–1548. [Google Scholar] [CrossRef]
- Franco, A.; Scieuzo, C.; Salvia, R.; Petrone, A.M.; Tafi, E.; Moretta, A.; Schmitt, E.; Falabella, P. Lipids from Hermetia illucens, an innovative and sustainable source. Sustainability 2021, 13, 10198. [Google Scholar] [CrossRef]
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- Van Huis, A.; Oonincx, D.G. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef]
- Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J. Bioconversion Potential of Agro-Industrial Byproducts by Tenebrio molitor—Long-Term Results. Insects 2022, 13, 810. [Google Scholar] [CrossRef]
- Yakti, W.; Schulz, S.; Marten, V.; Mewis, I.; Padmanabha, M.; Hempel, A.-J.; Kobelski, A.; Streif, S.; Ulrichs, C. The Effect of Rearing Scale and Density on the Growth and Nutrient Composition of Hermetia illucens (L.) (Diptera: Stratiomyidae) Larvae. Sustainability 2022, 14, 1772. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.-P.; van den Broek, L.A.; Fogliano, V.; Lakemond, C.M. Nitrogen-to-protein conversion factors for three edible insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef]
- Jones, C.G.; Hare, J.D.; Compton, S.J. Measuring plant protein with the Bradford assay. J. Chem. Ecol. 1989, 15, 979–992. [Google Scholar] [CrossRef] [PubMed]
- Mertens, C.; Dehon, L.; Bourgeois, A.; Verhaeghe-Cartrysse, C.; Blecker, C. Agronomical factors influencing the legumin/vicilin ratio in pea (Pisum sativum L.) seeds. J. Sci. Food Agric. 2012, 92, 1591–1596. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Mandrioli, M.; Tura, M.; Scotti, S.; Toschi, T.G. Fast detection of 10 cannabinoids by RP-HPLC-UV method in Cannabis sativa L. Molecules 2019, 24, 2113. [Google Scholar] [CrossRef]
- Rathi, V.; Singh, G.; Kumar, P.; Chaudhary, M.; Singh, P.; Mishra, M. Legality of worldwide cannabis use and associated economic benefits. In Revolutionizing the Potential of Hemp and Its Products in Changing the Global Economy; Springer: Berlin/Heidelberg, Germany, 2022; pp. 27–68. [Google Scholar]
- Struik, P.; Amaducci, S.; Bullard, M.; Stutterheim, N.; Venturi, G.; Cromack, H. Agronomy of fibre hemp (Cannabis sativa L.) in Europe. Ind. Crops Prod. 2000, 11, 107–118. [Google Scholar] [CrossRef]
- Tang, K.; Struik, P.; Yin, X.; Thouminot, C.; Bjelková, M.; Stramkale, V.; Amaducci, S. Comparing hemp (Cannabis sativa L.) cultivars for dual-purpose production under contrasting environments. Ind. Crops Prod. 2016, 87, 33–44. [Google Scholar] [CrossRef]
- Amaducci, S.; Gusovius, H.J. Hemp–cultivation, extraction and processing. In Industrial Applications of Natural Fibres: Structure, Properties and Technical Applications; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; pp. 109–134. [Google Scholar]
- Kleinhenz, M.D.; Magnin, G.; Ensley, S.M.; Griffin, J.J.; Goeser, J.; Lynch, E.; Coetzee, J.F. Nutrient concentrations, digestibility, and cannabinoid concentrations of industrial hemp plant components. Appl. Anim. Sci. 2020, 36, 489–494. [Google Scholar] [CrossRef]
- Commission European. Regulation (EC) No 767/2009 of the European Parliament and of the Council of 13 July 2009 on the placing on the market and use of feed, amending European Parliament and Council Regulation (EC) No 1831/2003 and repealing Council Directive 79/373/EEC, Commission Directive 80/511/EEC, Council Directives 82/471/EEC, 83/228/EEC, 93/74/EEC, 93/113/EC and 96/25/EC and Commission Decision 2004/217/EC. Off. J. Eur. Union. 2009, 229, 1–28. [Google Scholar]
- Shumo, M.; Osuga, I.M.; Khamis, F.M.; Tanga, C.M.; Fiaboe, K.K.; Subramanian, S.; Ekesi, S.; van Huis, A.; Borgemeister, C. The nutritive value of black soldier fly larvae reared on common organic waste streams in Kenya. Sci. Rep. 2019, 9, 10110. [Google Scholar] [CrossRef]
- Mertenat, A.; Diener, S.; Zurbrügg, C. Black Soldier Fly biowaste treatment–Assessment of global warming potential. Waste Manag. 2019, 84, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Broeckx, L.; Frooninckx, L.; Slegers, L.; Berrens, S.; Noyens, I.; Goossens, S.; Verheyen, G.; Wuyts, A.; Van Miert, S. Growth of Black Soldier Fly Larvae Reared on Organic Side-Streams. Sustainability 2021, 13, 12953. [Google Scholar] [CrossRef]
- Semwogerere, F.; Katiyatiya, C.L.; Chikwanha, O.C.; Marufu, M.C.; Mapiye, C. Bioavailability and bioefficacy of hemp by-products in ruminant meat production and preservation: A review. Front. Vet. Sci. 2020, 7, 572906. [Google Scholar] [CrossRef]
- Shahzad, A. Hemp fiber and its composites–a review. J. Compos. Mater. 2012, 46, 973–986. [Google Scholar] [CrossRef]
- Yakti, W.; Müller, M.; Klost, M.; Mewis, I.; Dannehl, D.E.; Ulrichs, C. Physical Properties of Substrates as a Driver for Hermetia illucens (L.) (Diptera: Stratiomyidae) Larvae Growth; Humboldt-Universität zu Berlin: Berlin, Germany, 2023. [Google Scholar]
- Lalander, C.; Ermolaev, E.; Wiklicky, V.; Vinnerås, B. Process efficiency and ventilation requirement in black soldier fly larvae composting of substrates with high water content. Sci. Total Environ. 2020, 729, 138968. [Google Scholar] [CrossRef]
- Cammack, J.A.; Tomberlin, J.K. The impact of diet protein and carbohydrate on select life-history traits of the black soldier fly Hermetia illucens (L.)(Diptera: Stratiomyidae). Insects 2017, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Tschirner, M.; Simon, A. Influence of different growing substrates and processing on the nutrient composition of black soldier fly larvae destined for animal feed. J. Insects Food Feed 2015, 1, 249–259. [Google Scholar] [CrossRef]
- Villazana, J.; Alyokhin, A. Tolerance of immature black soldier flies (Diptera: Stratiomyidae) to cold temperatures above and below freezing point. J. Econ. Entomol. 2019, 112, 2632–2637. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; Adler, P.H.; Myers, H.M. Development of the black soldier fly (Diptera: Stratiomyidae) in relation to temperature. Environ. Entomol. 2009, 38, 930–934. [Google Scholar] [CrossRef]
- Yang, F.; Tomberlin, J.K. Comparing Selected Life-History Traits of Black Soldier Fly (Diptera: Stratiomyidae) Larvae Produced in Industrial and Bench-Top-Sized Containers. J. Insect Sci. 2020, 20, 25. [Google Scholar] [CrossRef]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- Fowsiya, J.; Madhumitha, G. A review of bioinsecticidal activity and mode of action of plant derived alkaloids. Res. J. Pharm. Technol. 2020, 13, 963–973. [Google Scholar]
- Jeschke, V.; Kearney, E.E.; Schramm, K.; Kunert, G.; Shekhov, A.; Gershenzon, J.; Vassão, D.G. How glucosinolates affect generalist lepidopteran larvae: Growth, development and glucosinolate metabolism. Front. Plant Sci. 2017, 8, 1995. [Google Scholar] [CrossRef] [PubMed]
- Weththasinghe, P.; Hansen, J.Ø.; Mydland, L.T.; Øverland, M. A systematic meta-analysis based review on black soldier fly (Hermetia illucens) as a novel protein source for salmonids. Rev. Aquac. 2022, 14, 938–956. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Gort, G.; Dicke, M.; van Loon, J.J. Effects of dietary protein and carbohydrate on life-history traits and body protein and fat contents of the black soldier fly Hermetia illucens. Physiol. Entomol. 2019, 44, 148–159. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J. Influence of larval density and dietary nutrient concentration on performance, body protein, and fat contents of black soldier fly larvae (Hermetia illucens). Entomol. Exp. Appl. 2018, 166, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.; Romano, N.; Sinha, A.K. Conversion of spent coffee and donuts by black soldier fly (Hermetia illucens) larvae into potential resources for animal and plant farming. Insects 2021, 12, 332. [Google Scholar] [CrossRef]
- Oonincx, D.; Finke, M. Nutritional value of insects and ways to manipulate their composition. J. Insects Food Feed 2021, 7, 639–659. [Google Scholar] [CrossRef]
- Pinotti, L.; Manoni, M.; Ferrari, L.; Tretola, M.; Cazzola, R.; Givens, I. The Contribution of Dietary Magnesium in Farm Animals and Human Nutrition. Nutrients 2021, 13, 509. [Google Scholar] [CrossRef] [PubMed]
- Delezie, E.; Bierman, K.; Nollet, L.; Maertens, L. Impacts of calcium and phosphorus concentration, their ratio, and phytase supplementation level on growth performance, foot pad lesions, and hock burn of broiler chickens. J. Appl. Poult. Res. 2015, 24, 115–126. [Google Scholar] [CrossRef]
- Chia, S.Y.; Tanga, C.M.; Osuga, I.M.; Cheseto, X.; Ekesi, S.; Dicke, M.; van Loon, J.J. Nutritional composition of black soldier fly larvae feeding on agro-industrial by-products. Entomol. Exp. Appl. 2020, 168, 472–481. [Google Scholar] [CrossRef]
- Oonincx, D.G.; Van Broekhoven, S.; Van Huis, A.; van Loon, J.J. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef]
- Liland, N.S.; Biancarosa, I.; Araujo, P.; Biemans, D.; Bruckner, C.G.; Waagbø, R.; Torstensen, B.E.; Lock, E.-J. Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLoS ONE 2017, 12, e0183188. [Google Scholar] [CrossRef]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef]
- Mackie, K. Cannabinoid receptors as therapeutic targets. Annu. Rev. Pharmacol. Toxicol. 2006, 46, 101–122. [Google Scholar] [CrossRef]
- Pertwee, R. The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: Δ9-tetrahydrocannabinol, cannabidiol and Δ9-tetrahydrocannabivarin. Br. J. Pharmacol. 2008, 153, 199–215. [Google Scholar] [CrossRef] [PubMed]
- McPartland, J.; Di Marzo, V.; De Petrocellis, L.; Mercer, A.; Glass, M. Cannabinoid receptors are absent in insects. J. Comp. Neurol. 2001, 436, 423–429. [Google Scholar] [CrossRef]
- Park, S.-H.; Staples, S.K.; Gostin, E.L.; Smith, J.P.; Vigil, J.J.; Seifried, D.; Kinney, C.; Pauli, C.S.; Heuvel, B.D.V. Contrasting roles of cannabidiol as an insecticide and rescuing agent for ethanol–induced death in the tobacco hornworm Manduca sexta. Sci. Rep. 2019, 9, 10481. [Google Scholar] [CrossRef]
- He, J.; Tan, A.M.X.; Ng, S.Y.; Rui, M.; Yu, F. Cannabinoids modulate food preference and consumption in Drosophila melanogaster. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Borel, P.; Hammaz, F.; Morand-Laffargue, L.; Creton, B.; Halimi, C.; Sabatier, D.; Desmarchelier, C. Using black soldier fly larvae reared on fruits and vegetables waste as a sustainable dietary source of provitamin a carotenoids. Food Chem. 2021, 359, 129911. [Google Scholar] [CrossRef]
- Rademacher, J.; Krüger, K.; Wernecke, H.; Ehrhardt, E.; Reda, V.; Leifheit, M.; Yakti, W. Method for producing a feed enriched with at least one carotenoid or a feed component enriched with at least one carotenoid. European Patent Application EP3662760A1, 7 December 2019. [Google Scholar]
- Formato, M.; Crescente, G.; Scognamiglio, M.; Fiorentino, A.; Pecoraro, M.T.; Piccolella, S.; Catauro, M.; Pacifico, S. (−)-Cannabidiolic acid, a still overlooked bioactive compound: An introductory review and preliminary research. Molecules 2020, 25, 2638. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G.; Rock, E.M.; Guenther, K.; Limebeer, C.L.; Stevenson, L.A.; Haj, C.; Smoum, R.; Parker, L.A.; Mechoulam, R. Cannabidiolic acid methyl ester, a stable synthetic analogue of cannabidiolic acid, can produce 5-HT1A receptor-mediated suppression of nausea and anxiety in rats. Br. J. Pharmacol. 2018, 175, 100–112. [Google Scholar] [CrossRef]
- Rock, E.M.; Limebeer, C.L.; Parker, L.A. Effect of cannabidiolic acid and∆ 9-tetrahydrocannabinol on carrageenan-induced hyperalgesia and edema in a rodent model of inflammatory pain. Psychopharmacology 2018, 235, 3259–3271. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.d.M.; Santiago, N.R.; Alves, E.C.R.; Chaves, D.S.d.A.; Visacri, M.B. Use of cannabis in the treatment of animals: A systematic review of randomized clinical trials. Anim. Health Res. Rev. 2022, 23, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Melis, R.; Braca, A.; Sanna, R.; Spada, S.; Mulas, G.; Fadda, M.L.; Sassu, M.M.; Serra, G.; Anedda, R. Metabolic response of yellow mealworm larvae to two alternative rearing substrates. Metabolomics 2019, 15, 1–13. [Google Scholar] [CrossRef]
- Deruytter, D.; Coudron, C. The effects of density on the growth, survival and feed conversion of Tenebrio molitor larvae. J. Insects Food Feed 2022, 8, 141–146. [Google Scholar] [CrossRef]
- Eberle, S.; Schaden, L.-M.; Tintner, J.; Stauffer, C.; Schebeck, M. Effect of Temperature and Photoperiod on Development, Survival, and Growth Rate of Mealworms, Tenebrio molitor. Insects 2022, 13, 321. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Masri, J.; Perez, V.; Maya, C.; Zhao, J. Growth performance and nutrient composition of mealworms (Tenebrio molitor) fed on fresh plant materials-supplemented diets. Foods 2020, 9, 151. [Google Scholar] [CrossRef] [PubMed]
- Harsányi, E.; Juhász, C.; Kovács, E.; Huzsvai, L.; Pintér, R.; Fekete, G.; Varga, Z.I.; Aleksza, L.; Gyuricza, C. Evaluation of organic wastes as substrates for rearing Zophobas morio, Tenebrio molitor, and Acheta domesticus larvae as alternative feed supplements. Insects 2020, 11, 604. [Google Scholar] [CrossRef]
- González, C.M.; Garzón, R.; Rosell, C.M. Insects as ingredients for bakery goods. A comparison study of H. illucens, A. domestica and T. molitor flours. Innov. Food Sci. Emerg. Technol. 2019, 51, 205–210. [Google Scholar] [CrossRef]
- Ravzanaadii, N.; Kim, S.-H.; Choi, W.-H.; Hong, S.-J.; Kim, N.-J. Nutritional value of mealworm, Tenebrio molitor as food source. Int. J. Ind. Entomol. 2012, 25, 93–98. [Google Scholar] [CrossRef]
- Adámková, A.; Mlček, J.; Adámek, M.; Fišera, M.; Borkovcová, M.; Bednářová, M.; Hlobilová, V.; Vojáčková, K. Effect of temperature and feed on the mineral content and the content of selected heavy metals in mealworm. J. Elem. 2020, 25, 1077–1088. [Google Scholar]
- Simon, E.; Baranyai, E.; Braun, M.; Fábián, I.; Tóthmérész, B. Elemental concentration in mealworm beetle (Tenebrio molitor L.) during metamorphosis. Biol. Trace Elem. Res. 2013, 154, 81–87. [Google Scholar] [CrossRef]
- Finke, M.D. Complete nutrient content of four species of feeder insects. Zoo Biol. 2013, 32, 27–36. [Google Scholar] [CrossRef]
- Terra, W.R.; Ferreira, C. Evolutionary trends of digestion and absorption in the major insect orders. Arthropod Struct. Dev. 2020, 56, 100931. [Google Scholar] [CrossRef] [PubMed]
- Krenn, H.W. Form and function of insect mouthparts. In Insect Mouthparts; Springer: Berlin/Heidelberg, Germany, 2019; pp. 9–46. [Google Scholar]
- Bar-Hai, A.; Domb, A.J.; Hoffman, A. Strategies for enhancing the oral bioavailability of cannabinoids. Expert Opin. Drug Metab. Toxicol. 2022, 18, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Lähteenmäki-Uutela, A.; Marimuthu, S.; Meijer, N. Regulations on insects as food and feed: A global comparison. J. Insects Food Feed 2021, 7, 849–856. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T | Substrate Composition (Weight-Based) | The Mass Fraction of Cannabinoids [in µg/g DW] | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CF [%] | Water [%] | Straw [%] | Hemp [%] | CBDA | CBGA | CBG | CBD | ∆9-THC | CBC | ∆9-THC-A | CBGA | TC | |
| H0 | 12.85 | 63.73 | 24.42 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| H44 | 12.85 | 63.73 | 8.14 | 16.3 | 219.7 | 36.5 | 8.3 | 100.5 | 2.29 | 23.7 | 8.9 | 36.9 | 436.0 |
| H66 | 12.85 | 63.73 | 0 | 24.4 | 310.4 | 46.1 | 12.2 | 132 | 3.53 | 31.4 | 12.4 | 49.7 | 597.6 |
| Cannabidiolic acid (CBDA) | 1030.68 |
| Cannabigerolic acid (CBGA) | 134.21 |
| Cannabigerol (CBG) | 18.85 |
| Cannabidiol (CBD) | 642.98 |
| ∆-9-tetrahydrocannabinol (∆9-THC) | 14.94 |
| Cannabichromene (CBC) | 67.18 |
| ∆-9-tetrahydrocannabinolic acid (∆9-THC-A) | 46.54 |
| Cannabichromenic acid (CBCA) | 109.62 |
| Total cannabinoids | 2064.99 |
| Feeding Experiment | Treatment | Ca | Fe | K | Mg | P | Protein |
|---|---|---|---|---|---|---|---|
| g/kg | g/kg | g/kg | g/kg | g/kg | % | ||
| BSFL experiment | H0 | 14.89 | 0.27 | 9.51 | 1.37 | 3.93 | 5.41 |
| H44 | 20.10 | 0.65 | 20.98 | 1.60 | 5.18 | 7.58 | |
| H66 | 22.43 | 0.78 | 26.55 | 1.88 | 5.65 | 9.02 | |
| MW experiment | T0 | 1.07 | 0.16 | 15.38 | 5.44 | 12.71 | 10.82 |
| T33 | 9.19 | 0.19 | 19.13 | 5.25 | 10.07 | 9.82 | |
| T66 | 17.62 | 0.21 | 22.00 | 5.12 | 7.03 | 8.89 | |
| T100 | 26.06 | 0.23 | 25.79 | 4.97 | 4.10 | 7.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yakti, W.; Förster, N.; Müller, M.; Mewis, I.; Ulrichs, C. Hemp Waste as a Substrate for Hermetia illucens (L.) (Diptera: Stratiomyidae) and Tenebrio molitor L. (Coleoptera: Tenebrionidae) Rearing. Insects 2023, 14, 183. https://doi.org/10.3390/insects14020183
Yakti W, Förster N, Müller M, Mewis I, Ulrichs C. Hemp Waste as a Substrate for Hermetia illucens (L.) (Diptera: Stratiomyidae) and Tenebrio molitor L. (Coleoptera: Tenebrionidae) Rearing. Insects. 2023; 14(2):183. https://doi.org/10.3390/insects14020183
Chicago/Turabian StyleYakti, Wael, Nadja Förster, Marcus Müller, Inga Mewis, and Christian Ulrichs. 2023. "Hemp Waste as a Substrate for Hermetia illucens (L.) (Diptera: Stratiomyidae) and Tenebrio molitor L. (Coleoptera: Tenebrionidae) Rearing" Insects 14, no. 2: 183. https://doi.org/10.3390/insects14020183