
Effects of Climatic Change on Potential Distribution of Spogostylum ocyale (Diptera: Bombyliidae) in the Middle East Using Maxent Modelling

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Research Area
2.2. Distribution Data for S. ocyale
2.3. Environmental Data
2.4. Habitat Suitability Model
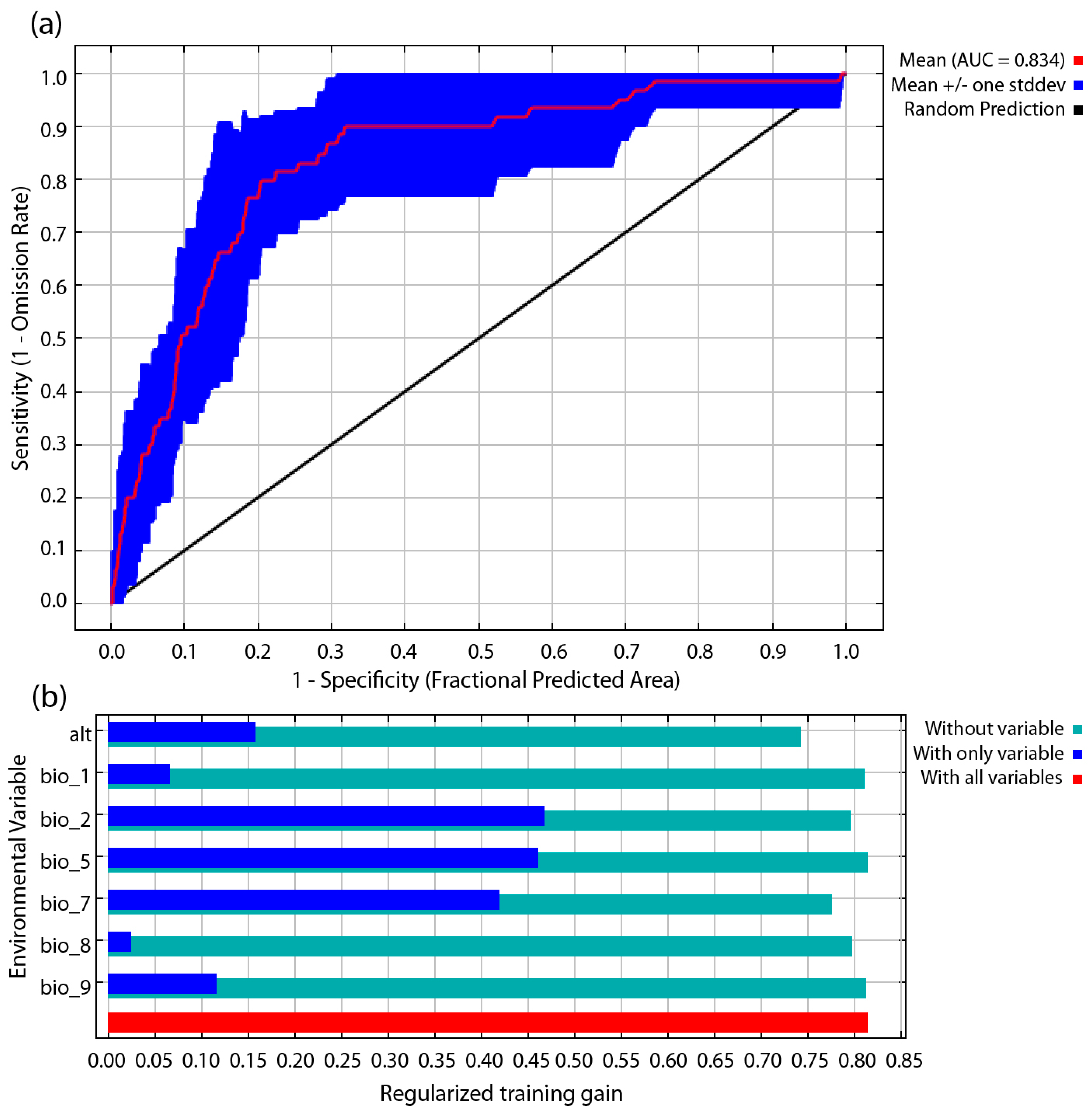
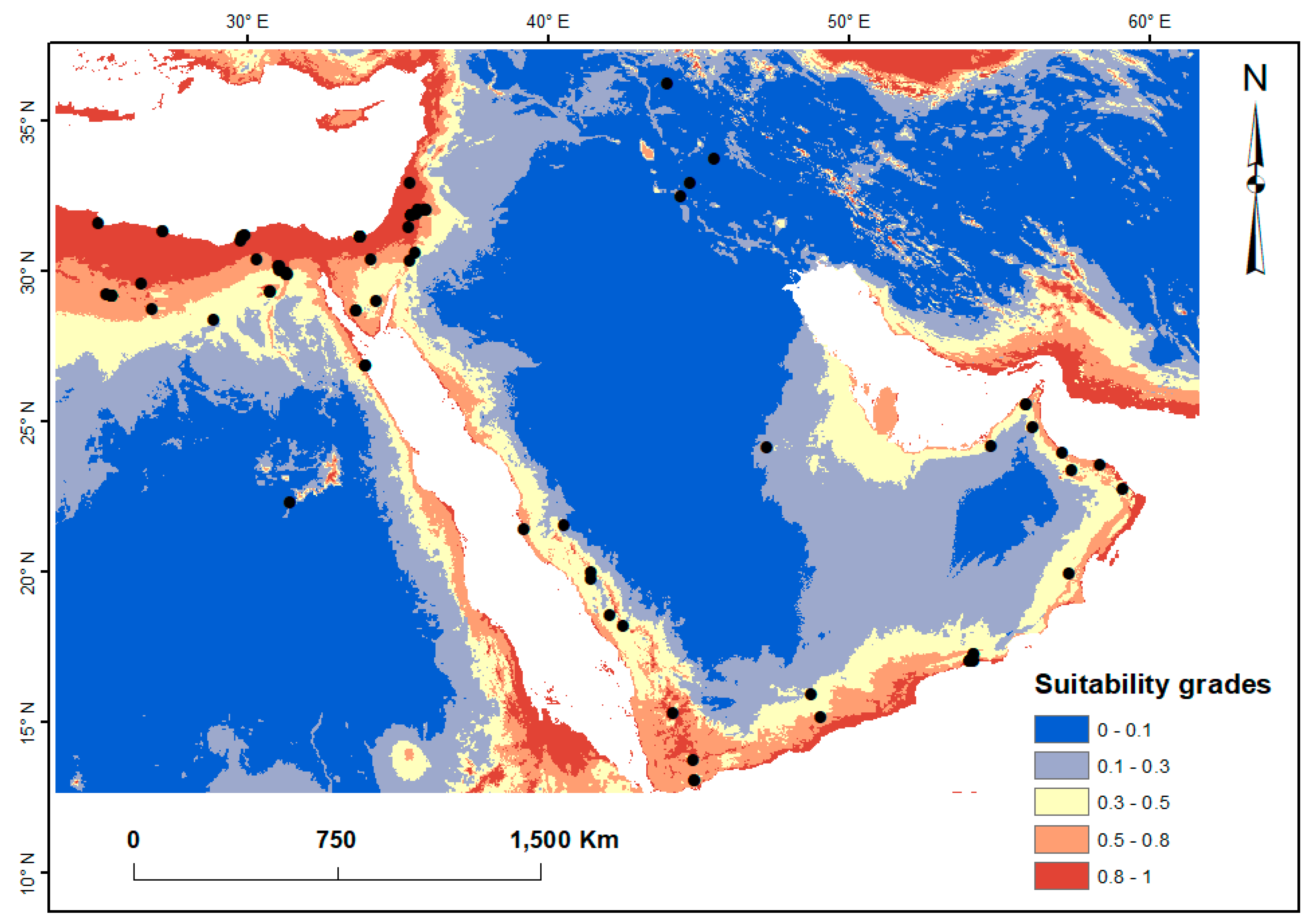
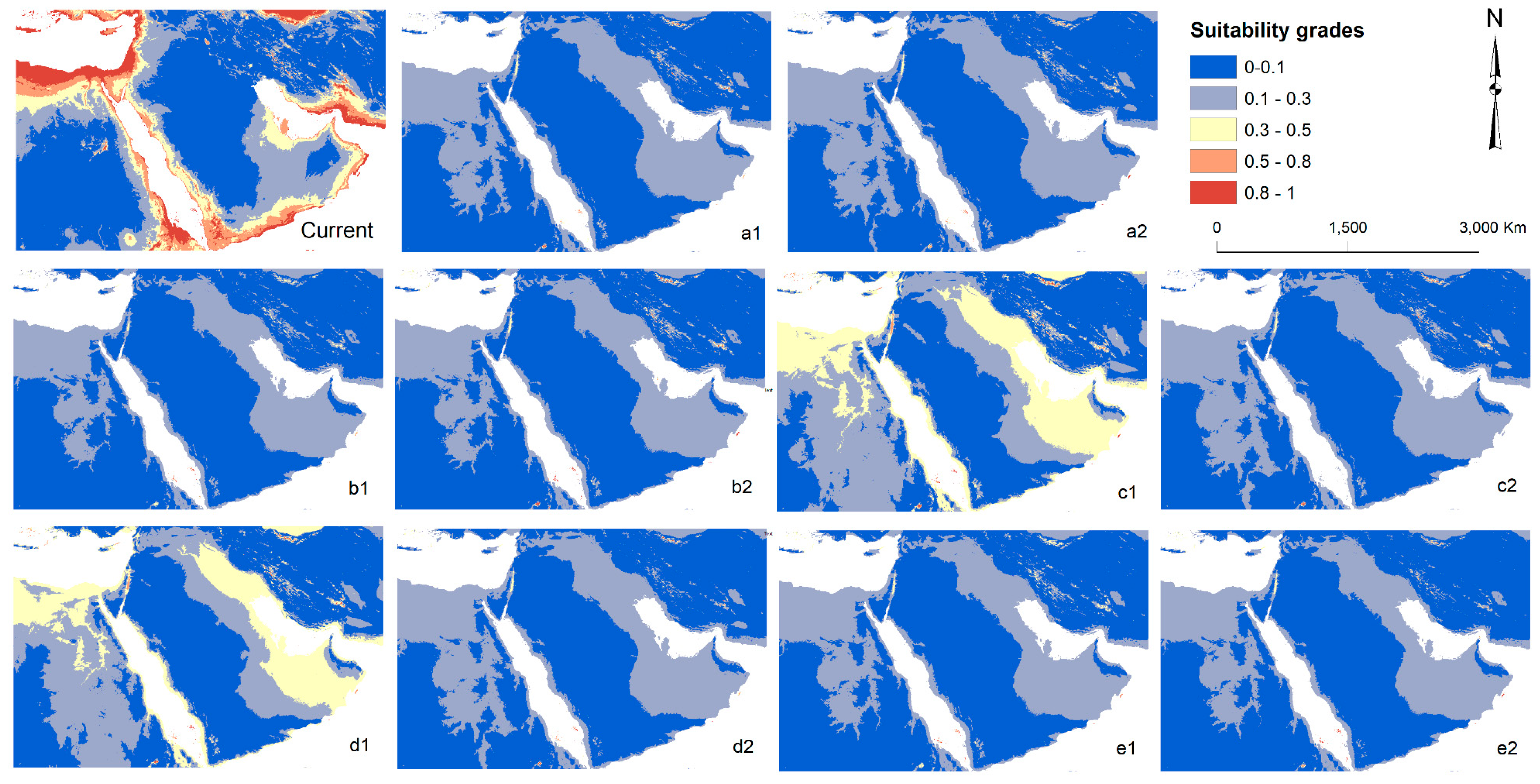
3. Results
3.1. Evaluations of the Model and Its Importance to Variables
3.2. Current Potential Distribution of Spogostylum ocyale
3.3. Future Potential Distribution of Spogostylum ocyale
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Hawagry, M.S.; El-Moursy, A.A.; Gilbert, F.; Zalat, S. The tribe Anthracini Latreille (Bombyliidae, Diptera) from Egypt. Egypt. J. Biol. 2000, 2, 97–117. [Google Scholar]
- El-Moursy, A.; Gilbert, F.; Zalat, S.; El-Hawagry, M. Foraging behaviour of anthracine flies (Diptera: Bombyliidae) in southern Sinai, Egypt. Egypt. J. Biol. 1999, 1, 87–95. [Google Scholar]
- Yeates, D.K.; Greathead, D. The evolutionary pattern of host use in the Bombyliidae (Diptera): A diverse family of parasitoid flies. Biol. J. Linn. Soc. 1997, 60, 149–185. [Google Scholar]
- El-Hawagry, M.S.; Dhafer, H. The family Bombyliidae in the Kingdom of Saudi Arabia (Diptera: Brachycera: Asiloidea). Zootaxa 2019, 4590, 59. [Google Scholar]
- Greathead, D.J.; Evenhuis, N.L. Family Bombyliidae. In Manual of Palaearctic Diptera (with Special Reference to Flies of Economic Importance); Papp, L., Darvas, B. Eds.; Science Herald, Budapest Hungary: 1997; Volume 2, pp. 487–512.
- Nasser, M.; El-Hawagry, M.; Okely, M. Environmental niche modeling for some species of the genus Anthrax Scopoli (Diptera: Bombyliidae) in Egypt, with special notes on St. Catherine protected area as a suitable habitat. J. Insect Conserv. 2019, 23, 831–841. [Google Scholar] [CrossRef]
- Toft, C.A. Resource shifts in bee flies (Bombyliidae): Interactions among species determine choice of resources. Oikos 1984, 43, 104–112. [Google Scholar] [CrossRef]
- El-Hawagry, M.S. A new species and an updated checklist of the genus Spogostylum Macquart (Bombyliidae: Diptera) from Egypt. Egypt. J. Biol. Pest Control. 2018, 28, 1–6. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araújo, M.B. Niche properties and geographical extent as predictors of species sensitivity to climate change. Global Ecol. Biogeogr. 2005, 14, 347–357. [Google Scholar]
- Almarinez, B.J.M.; Fadri, M.J.A.; Lasina, R.; Tavera, M.A.A.; Carvajal, T.M.; Watanabe, K.; Legaspi, J.C.; Amalin, D.M. A bioclimate-based maximum entropy model for Comperiella calauanica barrion, almarinez and amalin (Hymenoptera: Encyrtidae) in the Philippines. Insects 2021, 12, 26. [Google Scholar]
- Chhetri, B.; Badola, H.K.; Barat, S. Modelling climate change impacts on distribution of Himalayan pheasants. Ecol. Indic. 2021, 123, 107368. [Google Scholar] [CrossRef]
- Stephenson, K.; Wilson, B.; Taylor, M.; McLaren, K.; van Veen, R.; Kunna, J.; Campbell, J. Modelling Climate Change Impacts on Tropical Dry Forest Fauna. Sustainability 2022, 14, 4760. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Rymer, P.D.; Byrne, M.; Ruthrof, K.X.; Whinam, J.; McGeoch, M.; Bergstrom, D.M.; Guerin, G.R.; Sparrow, B.; Joseph, L. Impacts of recent climate change on terrestrial flora and fauna: Some emerging Australian examples. Austral Ecol. 2019, 44, 3–27. [Google Scholar] [CrossRef] [Green Version]
- Hotta, M.; Tsuyama, I.; Nakao, K.; Ozeki, M.; Higa, M.; Kominami, Y.; Hamada, T.; Matsui, T.; Yasuda, M.; Tanaka, N. Modeling future wildlife habitat suitability: Serious climate change impacts on the potential distribution of the Rock Ptarmigan Lagopus muta japonica in Japan’s northern Alps. BMC Ecol. 2019, 19, 1–14. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2014. [Google Scholar]
- Rousseau, J.S.; Betts, M.G. Factors influencing transferability in species distribution models. Ecography 2022, 2022, e06060. [Google Scholar]
- Millar, C.I.; Stephenson, N.L.; Stephens, S.L. Climate change and forests of the future: Managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef]
- Sallam, M.F.; Al Ahmed, A.M.; Abdel-Dayem, M.S.; Abdullah, M.A. Ecological niche modeling and land cover risk areas for rift valley fever vector, culex tritaeniorhynchus giles in Jazan, Saudi Arabia. PLoS ONE 2013, 8, e65786. [Google Scholar] [CrossRef]
- Naeem, M.; Alahmed, A.; Kheir, S.; Sallam, M. Spatial distribution modeling of Stegomyia aegypti and Culex tritaeniorhynchus (Diptera: Culicidae) in Al-bahah Province, Kingdom of Saudi Arabia. Trop Biomed 2016, 33, 295–310. [Google Scholar]
- Hochkirch, A. The insect crisis we can’t ignore. Nature 2016, 539, 141. [Google Scholar]
- El-Hawagry, M.; Gilbert, F. Zoogeographical affinities and faunal relationships of bee flies (Diptera: Bombyliidae) in Egypt. Zool. Middle East 2014, 60, 50–56. [Google Scholar] [CrossRef]
- Austen, E.E. The Bombyliidae of Palestine; British Museum (Natural History): London, UK, 1937. [Google Scholar]
- Greathead, D.J. A sympatric species pair: Spogostylum ocyale (Wiedemann, 1828) and S. griseipenne Macquart, 1850 (Diptera: Bombyliidae). Zootaxa 2003, 274, 1–6. [Google Scholar]
- Arabyat, S.; Katbeh-Bader, A.; Greathead, D. The bee flies (Diptera: Bombyliidae) of Jordan. Zootaxa 2004, 654, 1–48. [Google Scholar] [CrossRef]
- Zaitsev, V. On the fauna of bee flies (Diptera, Bombyliidae) of Iraq. Entomol. Obozr. 1976, 55, 691–697. [Google Scholar]
- Greathead, D. Bee flies (Bombyliidae: Diptera) from Oman. J. Oman Stud. Spec. Rep. 1980, 2, 233–250. [Google Scholar]
- Greathead, D. Insects of Saudi Arabia. Diptera: Fam. Bombyliidae. Fauna Saudi Arab. 1980, 2, 291–337. [Google Scholar]
- Greathead, D. Diptera: Fam. Bombyliidae of Saudi Arabia. Fauna Saudi Arab. 1988, 9, 90–113. [Google Scholar]
- Brun, P.; Zimmermann, N.E.; Hari, C.; Pellissier, L.; Karger, D.N. Global climate-related predictors at kilometre resolution for the past and future. Earth Syst. Sci. Data Discuss. 2022, 2022, 1–44. [Google Scholar]
- Lange, S. Trend-preserving bias adjustment and statistical downscaling with ISIMIP3BASD (v1. 0). Geosci. Model Dev. 2019, 12, 3055–3070. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. Maxent Software for Modeling Species Niches and Distributions (Version 3.4.4); 2022. Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 30 July 2022).
- Elith, J.H.; Graham, C.P.; Anderson, R.; Dudík, M.; Ferrier, S.; Guisan, A.J.; Hijmans, R.; Huettmann, F.R.; Leathwick, J.; Lehmann, A. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.S.; Xie, B.Y.; Wan, F.H.; Xiao, Q.M.; Dai, L.Y. The potential geographic distribution of Radopholus similis in China. Agric. Sci. China 2007, 6, 1444–1449. [Google Scholar]
- Yan, H.; He, J.; Zhao, Y.; Zhang, L.; Zhu, C.; Wu, D. Gentiana macrophylla response to climate change and vulnerability evaluation in China. Glob. Ecol. Conserv. 2020, 22, e00948. [Google Scholar] [CrossRef]
- Khanum, R.; Mumtaz, A.; Kumar, S. Predicting impacts of climate change on medicinal asclepiads of Pakistan using Maxent modeling. Acta Oecol. 2013, 49, 23–31. [Google Scholar]
- Gebrewahid, Y.; Abrehe, S.; Meresa, E.; Eyasu, G.; Abay, K.; Gebreab, G.; Kidanemariam, K.; Adissu, G.; Abreha, G.; Darcha, G. Current and future predicting potential areas of Oxytenanthera abyssinica (A. Richard) using MaxEnt model under climate change in Northern Ethiopia. Ecol. Process. 2020, 9, 1–15. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Ciss, M.; Biteye, B.; Fall, A.G.; Fall, M.; Gahn, M.C.B.; Leroux, L.; Apolloni, A. Ecological niche modelling to estimate the distribution of Culicoides, potential vectors of bluetongue virus in Senegal. BMC Ecol. 2019, 19, 1–12. [Google Scholar]
- Valencia-Rodríguez, D.; Jiménez-Segura, L.; Rogéliz, C.A.; Parra, J.L. Ecological niche modeling as an effective tool to predict the distribution of freshwater organisms: The case of the Sabaleta Brycon henni (Eigenmann, 1913). PLoS ONE 2021, 16, e0247876. [Google Scholar]
- Li, Y.; Li, M.; Li, C.; Liu, Z. Optimized maxent model predictions of climate change impacts on the suitable distribution of Cunninghamia lanceolata in China. Forests 2020, 11, 302. [Google Scholar]
- El-Hady, R.M.; El-Hawagry, M.S.; Soliman, M.M. Diversity and temporal variations of the leafhopper fauna (Cicadellidae, Auchenorrhyncha, Hemiptera) in two ecological zones of Egypt. J. Nat. Hist. 2020, 54, 2869–2887. [Google Scholar]
- Soliman, M.M.; El-Hady, R.M.; El-Hawagry, M.S. Zoogeography and diversity patterns of the family Cicadellidae (Hemiptera) in Toshka Region, Egypt. J. Nat. Hist. 2022, 56, 1199–1213. [Google Scholar] [CrossRef]
- Wermelinger, B.; Seifert, M. Temperature-dependent reproduction of the spruce bark beetle Ips typographus, and analysis of the potential population growth. Ecol. Entomol. 1999, 24, 103–110. [Google Scholar]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Hof, A.R.; Svahlin, A. The potential effect of climate change on the geographical distribution of insect pest species in the Swedish boreal forest. Scand. J. For. Res. 2016, 31, 29–39. [Google Scholar] [CrossRef]
- Urbani, F.; D’Alessandro, P.; Biondi, M. Using Maximum Entropy Modeling (MaxEnt) to predict future trends in the distribution of high altitude endemic insects in response to climate change. Bull. Insectol. 2017, 70, 189–200. [Google Scholar]
- Zittis, G.; Almazroui, M.; Alpert, P.; Ciais, P.; Cramer, W.; Dahdal, Y.; Fnais, M.; Francis, D.; Hadjinicolaou, P.; Howari, F. Climate change and weather extremes in the Eastern Mediterranean and Middle East. Rev. Geophys. 2022, 60, e2021RG000762. [Google Scholar] [CrossRef]
- Boys, W.A.; Adam, M. Predicting the distributions of regional endemic dragonflies using a combined model approach. Insect Conserv. Divers. 2021, 14, 52–66. [Google Scholar] [CrossRef]
- McCoshum, S.M.; Andreoli, S.L.; Stenoien, C.M.; Oberhauser, K.S.; Baum, K.A. Species distribution models for natural enemies of monarch butterfly (Danaus plexippus) larvae and pupae: Distribution patterns and implications for conservation. J. Insect Conserv. 2016, 20, 223–237. [Google Scholar] [CrossRef]
- Tavanpour, T.; Mehrnejad, M.R.; Sarafrazi, A.; Imani, S. Distribution modelling of four scelionid egg parasitoids of green stink bugs (Hemiptera: Pentatomidae). Biologia 2017, 72, 53–61. [Google Scholar]
- Koot, E.M.; Morgan-Richards, M.; Trewick, S.A. Climate change and alpine-adapted insects: Modelling environmental envelopes of a grasshopper radiation. R. Soc. Open Sci. 2022, 9, 211596. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 105, 6668–6672. [Google Scholar] [CrossRef] [Green Version]
- Soliman, M.; El-Hawagry, M. Impact of urbanisation and human activities on diversity and abundance of ulidiid flies (Diptera: Ulidiidae) in Wadi El-Natroun, Egypt. Afr. Entomol. 2020, 28, 415–424. [Google Scholar] [CrossRef]
- Titeux, N.; Maes, D.; Marmion, M.; Luoto, M.; Heikkinen, R.K. Inclusion of soil data improves the performance of bioclimatic envelope models for insect species distributions in temperate Europe. J. Biogeogr. 2009, 36, 1459–1473. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Variable Code | Description | Unit |
|---|---|---|
| Bio1 | Annual mean temperature | °C |
| Bio2 | Mean diurnal range (max temp−min temp) (monthly average) | °C |
| Bio3 | Isothermality (Bio1/Bio7) × 100 | °C |
| Bio4 | Temperature seasonality (coefficient of variation) | °C |
| Bio5 | Max temperature of warmest period | °C |
| Bio6 | Min temperature of the coldest period | °C |
| Bio7 | Temperature annual range | °C |
| Bio8 | Mean temperature of wettest quarter | °C |
| Bio9 | Mean temperature of driest quarter | °C |
| Bio10 | Mean temperature of warmest quarter | °C |
| Bio11 | Mean temperature of coldest quarter | °C |
| Bio12 | Annual precipitation | mm |
| Bio13 | Precipitation of wettest period | mm |
| Bio14 | Precipitation of driest period | mm |
| Bio15 | Precipitation seasonality (coefficient of variation) | mm |
| Bio16 | Precipitation of wettest quarter | mm |
| Bio17 | Precipitation of driest quarter | mm |
| Bio18 | Precipitation of warmest quarter | mm |
| Bio19 | Precipitation of coldest quarter | mm |
| Alt | Altitude in degrees | m |
| Variables | % Contribution | Permutation Importance |
|---|---|---|
| Bio5 | 34.4 | 0 |
| Bio7 | 32.5 | 63.3 |
| Bio2 | 14.3 | 4.1 |
| Alt | 12.1 | 15.9 |
| Bio8 | 6.1 | 6.8 |
| Bio1 | 0.5 | 9.9 |
| Bio9 | 0.1 | 0 |
| Model ID | Institution | Native Resolution | Ensemble Member | Priority a |
|---|---|---|---|---|
| GFDL-ESM4 | National Oceanic and Atmospheric Administration, Geophysical Fluid Dynamics Laboratory, USA | 288 × 180 | r1i1p1f1 | 1 |
| IPSL-CM6A-LR | Institute Pierre Simon Laplace, France | 144 × 143 | r1i1p1f1 | 4 |
| MPI-ESM1-2-HR | Max Planck Institute for Meteorology, Germany | 384 × 192 | r1i1p1f1 | 3 |
| MRI-ESM2-0 | Meteorological Research Institute, Japan | 320 × 160 | r1i1p1f1 | 5 |
| UKESM1-0-LL | Met Office Hadley Centre, UK | 192 × 144 | r1i1p1f2 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soliman, M.M.; Al-Khalaf, A.A.; El-Hawagry, M.S.A. Effects of Climatic Change on Potential Distribution of Spogostylum ocyale (Diptera: Bombyliidae) in the Middle East Using Maxent Modelling. Insects 2023, 14, 120. https://doi.org/10.3390/insects14020120
Soliman MM, Al-Khalaf AA, El-Hawagry MSA. Effects of Climatic Change on Potential Distribution of Spogostylum ocyale (Diptera: Bombyliidae) in the Middle East Using Maxent Modelling. Insects. 2023; 14(2):120. https://doi.org/10.3390/insects14020120
Chicago/Turabian StyleSoliman, Mustafa M., Areej A. Al-Khalaf, and Magdi S. A. El-Hawagry. 2023. "Effects of Climatic Change on Potential Distribution of Spogostylum ocyale (Diptera: Bombyliidae) in the Middle East Using Maxent Modelling" Insects 14, no. 2: 120. https://doi.org/10.3390/insects14020120