
Near-Infrared Reflectance Spectroscopy for Quantitative Analysis of Fat and Fatty Acid Content in Living Tenebrio molitor Larvae to Detect the Influence of Substrate on Larval Composition

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Samples
2.2. Feeding Treatments
2.3. Calculations
2.4. Analysis of Fat Content
2.5. Methylation and Analysis of Fatty Acid Composition
2.6. Statistical Analysis
2.7. Near-Infrared Spectra Collection
2.8. Chemometrics
3. Results
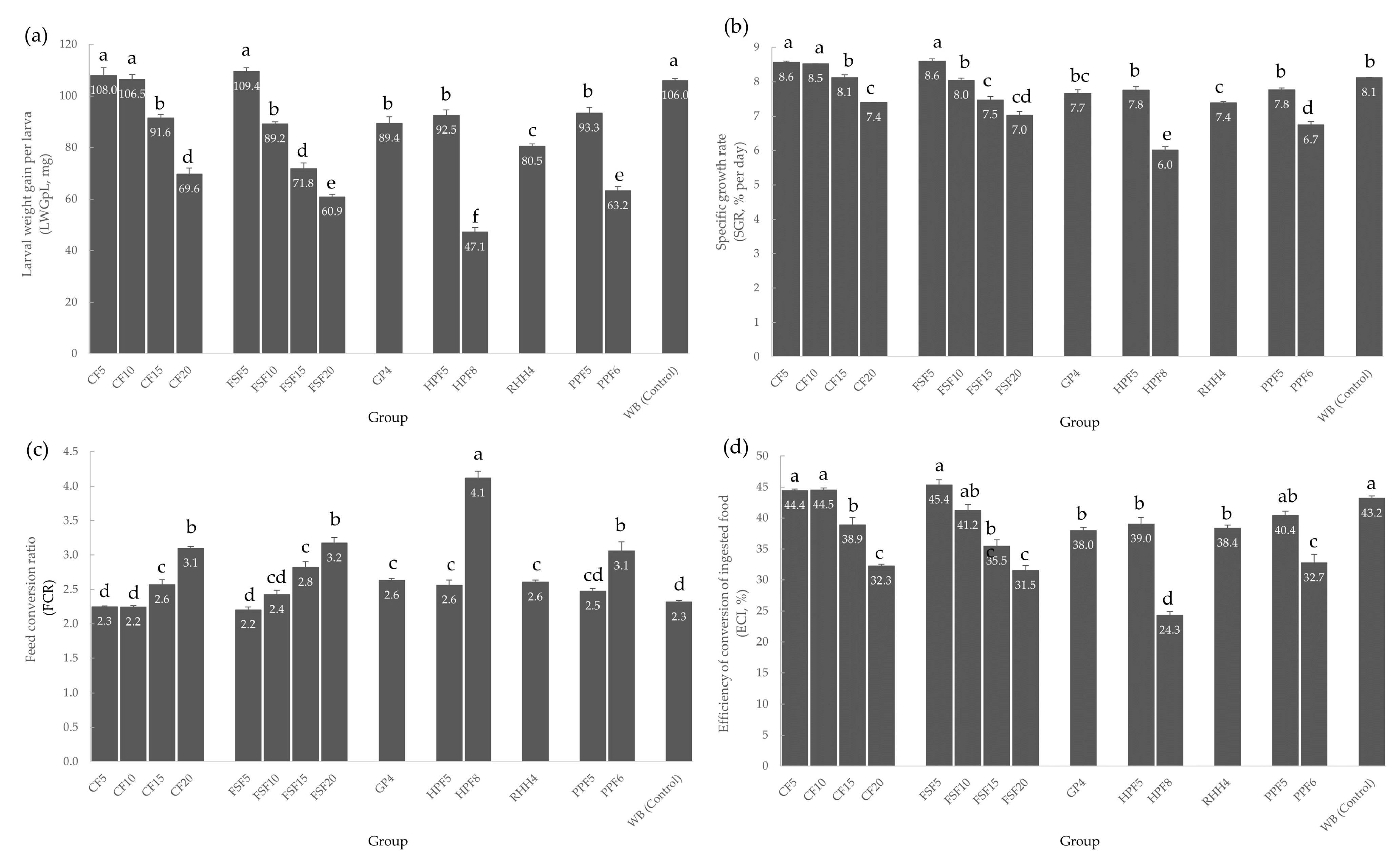
3.1. Larval Growth and Feed Conversion Parameters
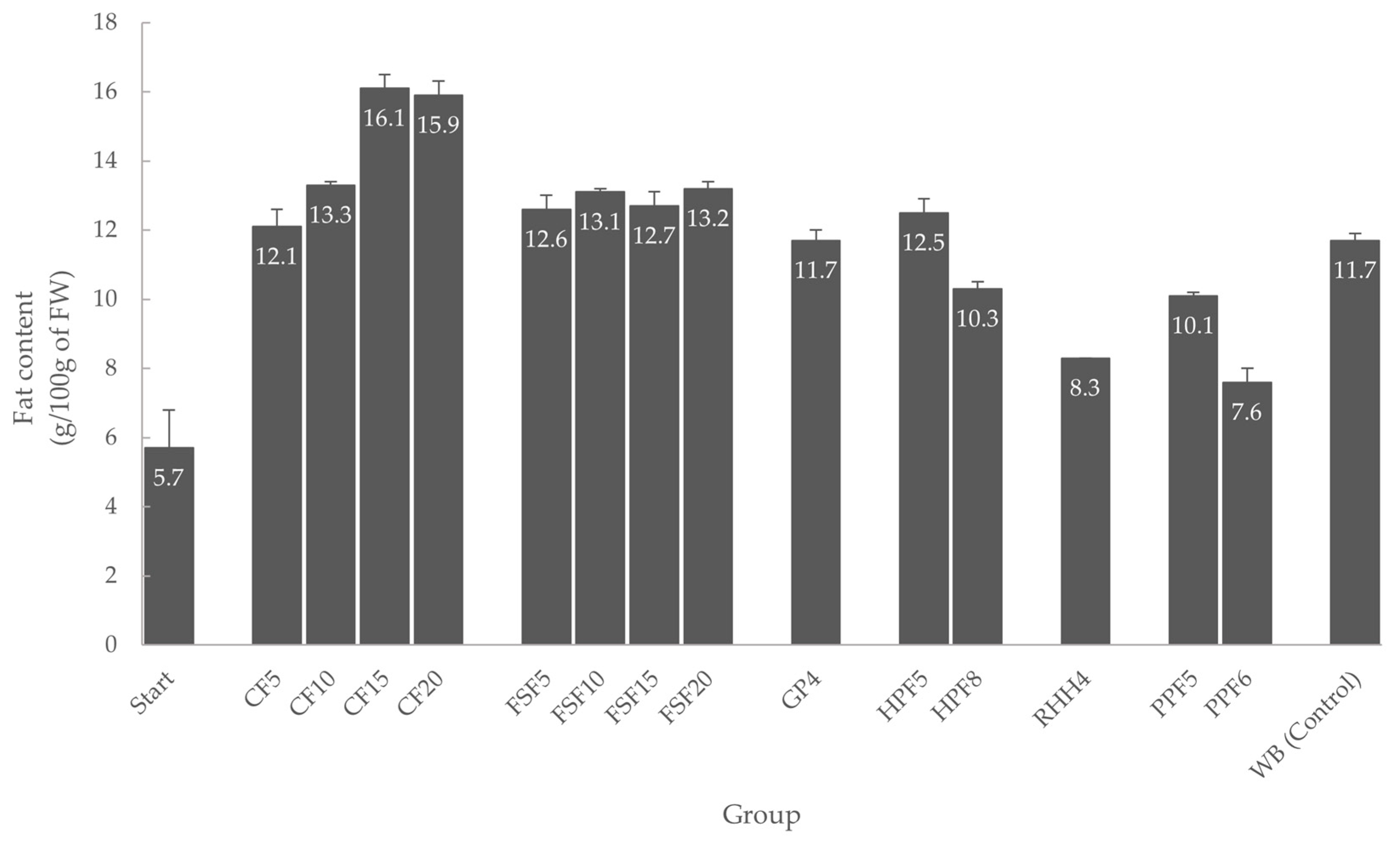
3.2. Fat Content of Mealworm Larvae
3.3. Fatty Acid Composition of Mealworm Larvae
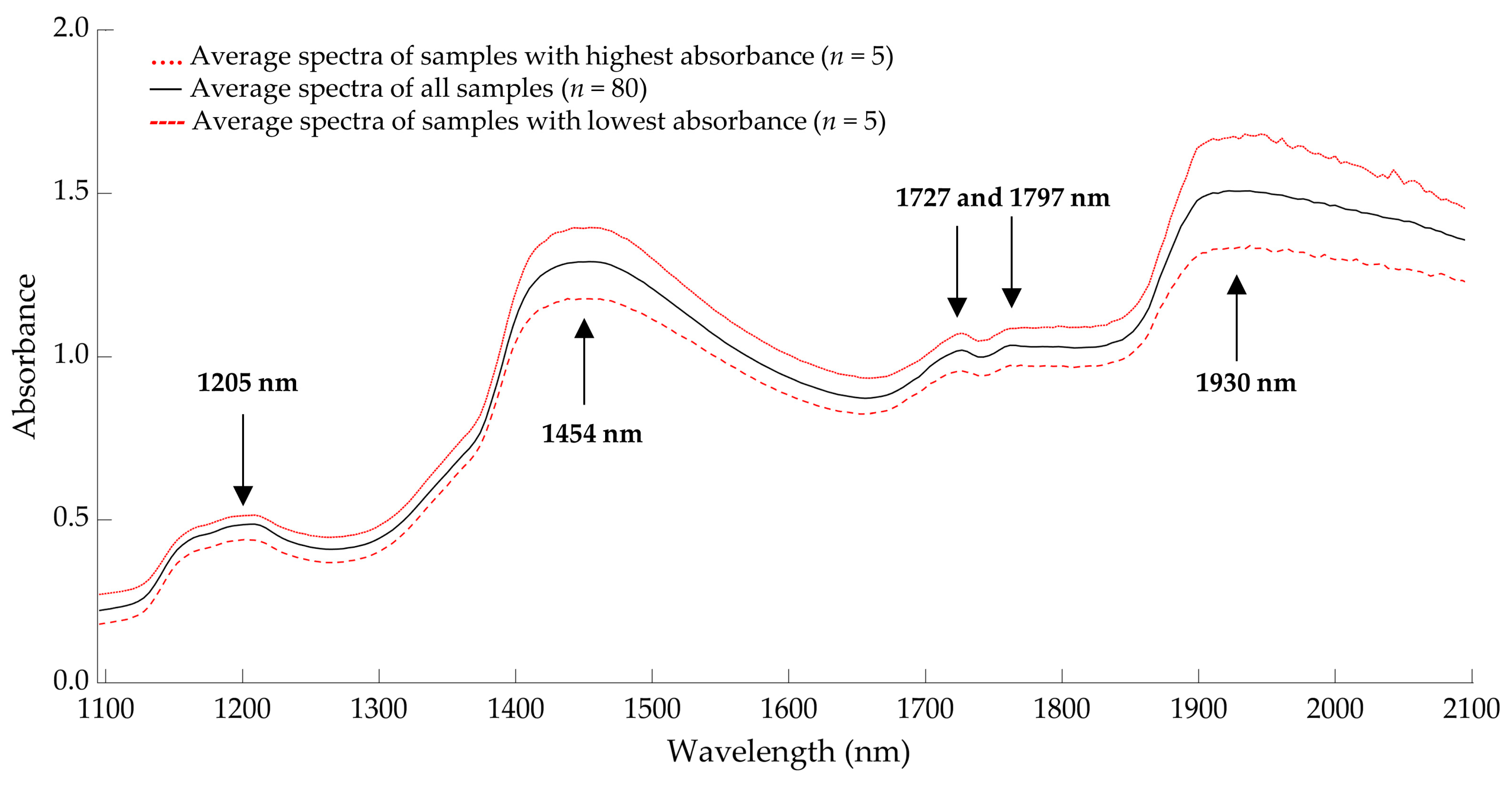
3.4. Near-Infrared Spectra
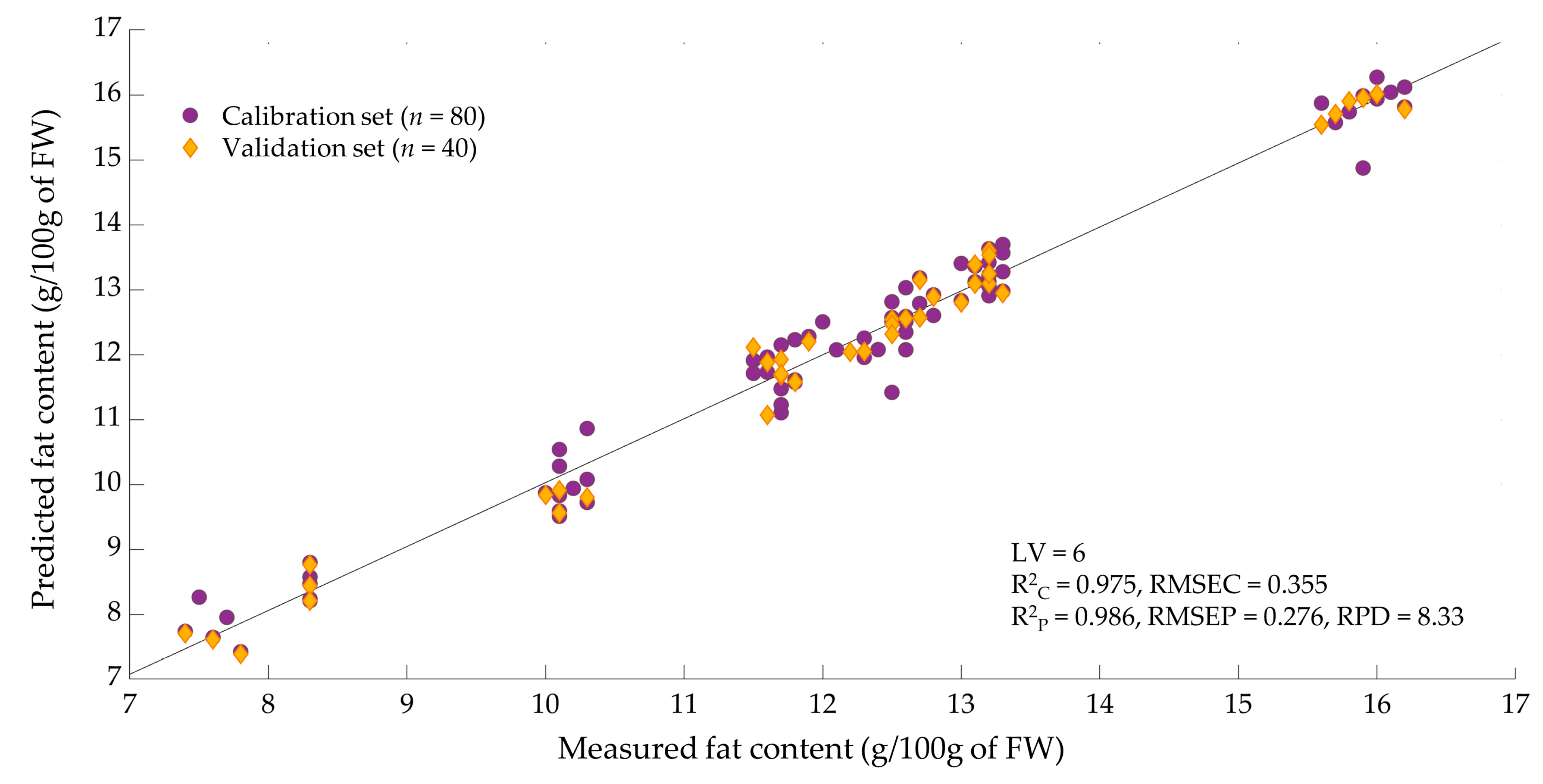
3.5. Prediction Model of Larval Fat Content
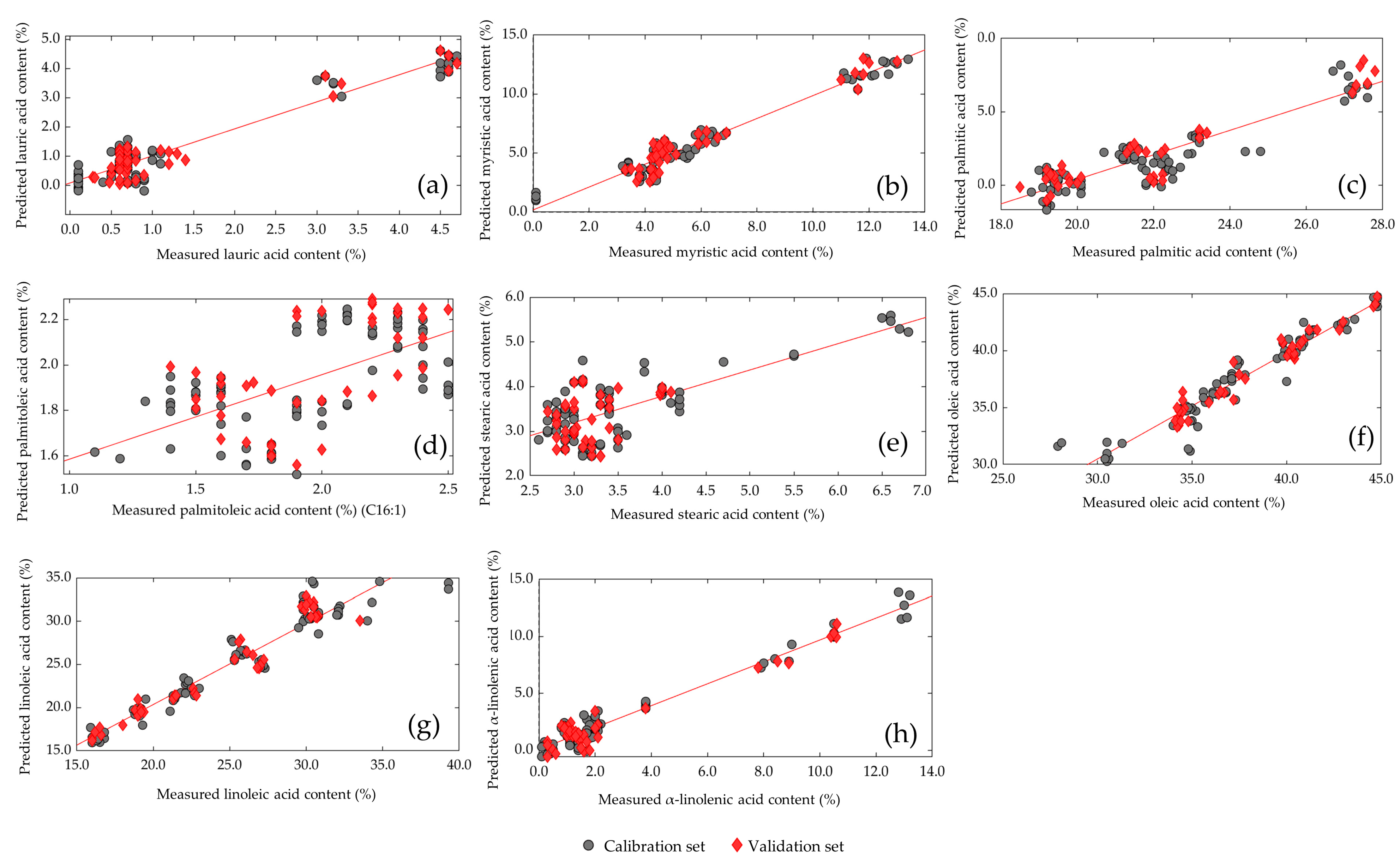
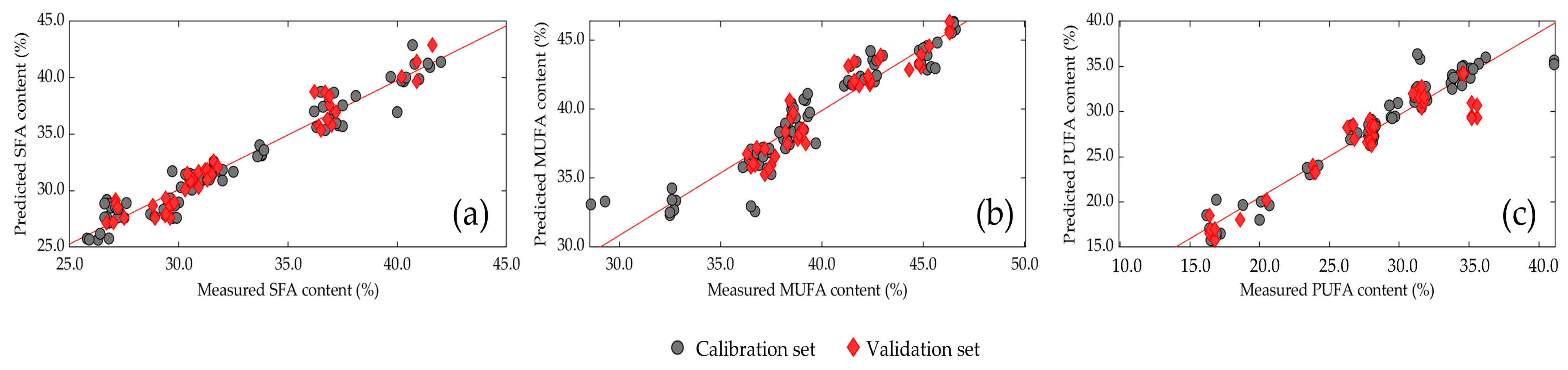
3.6. Prediction Model of Larval Fatty Acid Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grafton, R.Q.; Daugbjerg, C.; Qureshi, M.E. Towards food security by 2050. Food Sec. 2015, 7, 179–183. [Google Scholar] [CrossRef]
- Belluco, S.; Losasso, C.; Maggioletti, M.; Alonzi, C.C.; Paoletti, M.G.; Ricci, A. Edible Insects in a Food Safety and Nutritional Perspective: A Critical Review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 296–313. [Google Scholar] [CrossRef]
- van Huis, A. Edible Insects—Future Prospects For Food and Feed Security; FAO: Rome, Italy, 2014. [Google Scholar]
- van Huis, A.; Oonincx, D.G.A.B. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Dobermann, D.; Swift, J.A.; Field, L.M. Opportunities and hurdles of edible insects for food and feed. Nutr. Bull. 2017, 42, 293–308. [Google Scholar] [CrossRef] [Green Version]
- Finke, M.D. Complete nutrient content of four species of feeder insects. Zoo Biol. 2013, 32, 27–36. [Google Scholar] [CrossRef]
- Oonincx, D.; Finke, M.D. Nutritional value of insects and ways to manipulate their composition. J. Insects Food Feed. 2021, 7, 639–659. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; Laurent, S.; Veenenbos, M.E.; van Loon, J.J.A. Dietary enrichment of edible insects with omega 3 fatty acids. Insect Sci. 2020, 27, 500–509. [Google Scholar] [CrossRef] [Green Version]
- Canavoso, L.E.; Jouni, Z.E.; Karnas, K.J.; Pennington, J.E.; Wells, M.A. Fat metabolism in insects. Annu. Rev. Nutr. 2001, 21, 23–46. [Google Scholar] [CrossRef]
- Francardi, V.; Cito, A.; Fusi, S.; Botta, M.; Dreassi, E. Linseed to increase n-3 fatty acids in Tenebrio molitor (Coleoptera Tenebrionidae). Redia 2017, 100, 73–76. [Google Scholar] [CrossRef]
- Prieto, N.; Roehe, R.; Lavín, P.; Batten, G.; Andrés, S. Application of near infrared reflectance spectroscopy to predict meat and meat products quality: A review. Meat Sci. 2009, 83, 175–186. [Google Scholar] [CrossRef]
- Duthen, S.; Levasseur-Garcia, C.; Kleiber, D.; Violleau, F.; Vaca-Garcia, C.; Tsuchikawa, S.; Delgado Raynaud, C.; Daydé, J. Using near-infrared spectroscopy to determine moisture content, gel strength, and viscosity of gelatin. Food Hydrocoll. 2021, 115, 106627. [Google Scholar] [CrossRef]
- Benes, E.; Gere, A.; Fodor, M. Predicting macronutrients and energy content of snack products using FT-NIR analysis and chemometric techniques. J. Food Eng. 2020, 280, 109954. [Google Scholar] [CrossRef]
- Ingle, P.D.; Christian, R.; Purohit, P.; Zarraga, V.; Handley, E.; Freel, K.; Abdo, S. Determination of Protein Content by NIR Spectroscopy in Protein Powder Mix Products. J. AOAC Int. 2016, 99, 360–363. [Google Scholar] [CrossRef]
- Afseth, N.K.; Dankel, K.; Andersen, P.V.; Difford, G.F.; Horn, S.S.; Sonesson, A.; Hillestad, B.; Wold, J.P.; Tengstrand, E. Raman and near Infrared Spectroscopy for Quantification of Fatty Acids in Muscle Tissue-A Salmon Case Study. Foods 2022, 11, 962. [Google Scholar] [CrossRef]
- Noel, S.J.; Jørgensen, H.J.H.; Bach Knudsen, K.E. Prediction of protein and amino acid composition and digestibility in individual feedstuffs and mixed diets for pigs using near-infrared spectroscopy. Anim. Nutr. 2021, 7, 1242–1252. [Google Scholar] [CrossRef]
- Dowell, F.E.; Throne, J.E.; Wang, D.; Baker, J.E. Identifying Stored-Grain Insects Using Near-Infrared Spectroscopy. J. Econ. Entomol. 1999, 92, 165–169. [Google Scholar] [CrossRef]
- Perez-Mendoza, J.; Throne, J.E.; Dowell, F.E.; Baker, J.E. Detection of insect fragments in wheat flour by near-infrared spectroscopy. J. Stored Prod. Res. 2003, 39, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Prevolnik, M.; Čandek-Potokar, M.; Škorjanc, D.; Velikonja-Bolta, Š.; Škrlep, M.; Žnidaršic, T.; Babnik, D. Predicting Intramuscular Fat Content in Pork and Beef by near Infrared Spectroscopy. J. Near Infrared Spectrosc. 2005, 13, 77–85. [Google Scholar] [CrossRef]
- Wang, H.L.; Wan, X.Y.; Bi, J.C.; Wang, J.K.; Jiang, L.; Chen, L.M.; Zhai, H.Q.; Wan, J.M. Quantitative Analysis of Fat Content in Rice by Near-Infrared Spectroscopy Technique. Cereal Chem. J. 2006, 83, 402–406. [Google Scholar] [CrossRef]
- Hashimoto, J.C.; Lima, J.C.; Celeghini, R.M.S.; Nogueira, A.B.; Efraim, P.; Poppi, R.J.; Pallone, J.A.L. Quality Control of Commercial Cocoa Beans (Theobroma cacao L.) by Near-infrared Spectroscopy. Food Anal. Methods 2018, 11, 1510–1517. [Google Scholar] [CrossRef]
- Kröncke, N.; Benning, R. Determination of Moisture and Protein Content in Living Mealworm Larvae (Tenebrio molitor L.) Using Near-Infrared Reflectance Spectroscopy (NIRS). Insects 2022, 13, 560. [Google Scholar] [CrossRef] [PubMed]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Broekhoven, S.; Oonincx, D.G.A.B.; van Huis, A.; van Loon, J.J.A. Growth performance and feed conversion efficiency of three edible mealworm species (Coleoptera: Tenebrionidae) on diets composed of organic by-products. J. Insect Physiol. 2015, 73, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Waldbauer, G.P. The Consumption and Utilization of Food by Insects. In Advances in Insect Physiology; Academic Press: London, UK, 1968; Volume 5, pp. 229–288. [Google Scholar]
- VDLUFA. VDLUFA Methodenbuch III. In Band III die Chemische Untersuchung von Futtermitteln; VDLUFA: Bonn, Germany, 2013; 2190p. [Google Scholar]
- Chen, L.; Yang, Z.; Han, L. A Review on the Use of Near-Infrared Spectroscopy for Analyzing Feed Protein Materials. Appl. Spectrosc. Rev. 2013, 48, 509–522. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, C.; Shao, Q.; Yang, Z.; Zhang, X.; Xu, X.; Hassan, M. Determination of water content in corn stover silage using near-infrared spectroscopy. Int. J. Agric. Biol. Eng. 2019, 12, 143–148. [Google Scholar] [CrossRef]
- Lin, C.; Chen, X.; Jian, L.; Shi, C.; Jin, X.; Zhang, G. Determination of grain protein content by near-infrared spectrometry and multivariate calibration in barley. Food Chem. 2014, 162, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Daszykowski, M.; Wrobel, M.S.; Czarnik-Matusewicz, H.; Walczak, B. Near-infrared reflectance spectroscopy and multivariate calibration techniques applied to modelling the crude protein, fibre and fat content in rapeseed meal. Analyst 2008, 133, 1523–1531. [Google Scholar] [CrossRef] [PubMed]
- Burns, D.A. Handbook of Near-Infrared Analysis, 3rd ed.; Practical Spectroscopy; Taylor & Francis Group: Baton Rouge, LA, USA, 2007. [Google Scholar]
- Barlocco, N.; Vadell, A.; Ballesteros, F.; Galietta, G.; Cozzolino, D. Predicting intramuscular fat, moisture and Warner-Bratzler shear force in pork muscle using near infrared reflectance spectroscopy. Anim. Sci. 2006, 82, 111–116. [Google Scholar] [CrossRef]
- Chang, C.-W.; Laird, D.A. Near-infrared reflectance spectroscopic analysis of soil C and N. Soil Sci. 2002, 167, 110–116. [Google Scholar] [CrossRef]
- Bjørge, J.D.; Overgaard, J.; Malte, H.; Gianotten, N.; Heckmann, L.-H. Role of temperature on growth and metabolic rate in the tenebrionid beetles Alphitobius diaperinus and Tenebrio molitor. J. Insect Physiol. 2018, 107, 89–96. [Google Scholar] [CrossRef]
- Hoc, B.; Genva, M.; Fauconnier, M.-L.; Lognay, G.; Francis, F.; Caparros Megido, R. About lipid metabolism in Hermetia illucens (L. 1758): On the origin of fatty acids in prepupae. Sci. Rep. 2020, 10, 11916. [Google Scholar] [CrossRef] [PubMed]
- van Dooremalen, C.; Ellers, J. A moderate change in temperature induces changes in fatty acid composition of storage and membrane lipids in a soil arthropod. J. Insect Physiol. 2010, 56, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Lockey, K.H. Lipids of the insect cuticle: Origin, composition and function. Comp. Biochem. Physiol. Part B Comp. Biochem. 1988, 89, 595–645. [Google Scholar] [CrossRef]
- Stanley, D. Prostaglandins and other eicosanoids in insects: Biological significance. Annu. Rev. Entomol. 2006, 51, 25–44. [Google Scholar] [CrossRef] [PubMed]
- Cito, A.; Dreassi, E.; Frosinini, R.; Zanfini, A.; Pianigiani, C.; Botta, M.; Francardi, V. The potential beneficial effects of Tenebrio molitor (Coleoptera Tenebrionidae) and Galleria mellonella (Lepidoptera Pyralidae) on human health. Redia 2017, 100, 125–133. [Google Scholar] [CrossRef]
- Finke, M.D. Complete nutrient composition of commercially raised invertebrates used as food for insectivores. Zoo Biol. 2002, 21, 269–285. [Google Scholar] [CrossRef]
- Paul, A.; Frederich, M.; Megido, R.C.; Alabi, T.; Malik, P.; Uyttenbroeck, R.; Francis, F.; Blecker, C.; Haubruge, E.; Lognay, G.; et al. Insect fatty acids: A comparison of lipids from three Orthopterans and Tenebrio molitor L. larvae. J. Asia-Pac. Entomol. 2017, 20, 337–340. [Google Scholar] [CrossRef]
- Ravzanaadii, N.; Kim, S.-H.; Choi, W.-H.; Hong, S.-J.; Kim, N.-J. Nutritional Value of Mealworm, Tenebrio molitor as Food Source. Int. J. Ind. Entomol. 2012, 25, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Fraenkel, G.; Blewett, M. Linoleic acid and arachidonic acid in the metabolism of two insects, Ephestia kuehniella (Lep.) and Tenebrio molitor (Col.). Biochem. J. 1947, 41, 475–478. [Google Scholar] [CrossRef] [Green Version]
- Stanley-Samuelson, D.W.; Jurenka, R.A.; Cripps, C.; Blomquist, G.J.; Renobales, M. de Fatty acids in insects: Composition, metabolism, and biological significance. Arch. Insect Biochem. Physiol. 1988, 9, 1–33. [Google Scholar] [CrossRef]
- Gilby, A.R. Lipids and Their Metabolism in Insects. Annu. Rev. Entomol. 1965, 10, 141–160. [Google Scholar] [CrossRef]
- Mittler, T.E. Metabolic Aspects of Lipid Nutrition in Insects; CRC Press: Boca Raton, FL, USA, 1983. [Google Scholar]
- Stanley-Samuelson, D.W.; Dadd, R.H. Long-chain polyunsaturated fatty acids: Patterns of occurrence in insects. Insect Biochem. 1983, 13, 549–558. [Google Scholar] [CrossRef]
- Siemianowska, E.; Kosewska, A.; Aljewicz, M.; Skibniewska, K.A.; Polak-Juszczak, L.; Jarocki, A.; Jędras, M. Larvae of mealworm (Tenebrio molitor L.) as European novel food. ASC 2013, 4, 287–291. [Google Scholar] [CrossRef] [Green Version]
- St-Hilaire, S.; Cranfill, K.; McGuire, M.A.; Mosley, E.E.; Tomberlin, J.K.; Newton, L.; Sealey, W.; Sheppard, C.; Irving, S. Fish Offal Recycling by the Black Soldier Fly Produces a Foodstuff High in Omega-3 Fatty Acids. J. World Aquaculture Soc. 2007, 38, 309–313. [Google Scholar] [CrossRef]
- Chapman, R.F. The Insects. Structure and Function, 4th ed.; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Jenkins, T.C.; Wallace, R.J.; Moate, P.J.; Mosley, E.E. Board-invited review: Recent advances in biohydrogenation of unsaturated fatty acids within the rumen microbial ecosystem. J. Anim. Sci. 2008, 86, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Lawal, K.G.; Kavle, R.R.; Akanbi, T.O.; Mirosa, M.; Agyei, D. Enrichment in specific fatty acids profile of Tenebrio molitor and Hermetia illucens larvae through feeding. Future Foods 2021, 3, 100016. [Google Scholar] [CrossRef]
- Giannetto, A.; Oliva, S.; Ceccon Lanes, C.F.; de Araújo Pedron, F.; Savastano, D.; Baviera, C.; Parrino, V.; Lo Paro, G.; Spanò, N.C.; Cappello, T.; et al. Hermetia illucens (Diptera: Stratiomydae) larvae and prepupae: Biomass production, fatty acid profile and expression of key genes involved in lipid metabolism. J. Biotechnol. 2020, 307, 44–54. [Google Scholar] [CrossRef]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- Jones, L.D.; Cooper, R.W.; Harding, R.S. Composition of Mealworm Tenebrio molitor Larvae. J. Zoo Anim. Med. 1972, 3, 34. [Google Scholar] [CrossRef]
- Mariod, A.A. (Ed.) African Edible Insects as Alternative Source of Food, Oil, Protein and Bioactive Components; Springer: Cham, Germany, 2020. [Google Scholar]
- Yi, L.; Lakemond, C.M.M.; Sagis, L.M.C.; Eisner-Schadler, V.; van Huis, A.; van Boekel, M.A.J.S. Extraction and characterisation of protein fractions from five insect species. Food Chem. 2013, 141, 3341–3348. [Google Scholar] [CrossRef]
- Silva, L.B.; de Souza, R.G.; da Silva, S.R.; Da Feitosa, A.C.; Lopes, E.C.; Lima, S.B.P.; Dourado, L.R.B.; Pavan, B.E. Development of Tenebrio molitor (Coleoptera: Tenebrionidae) on Poultry Litter-Based Diets: Effect on Chemical Composition of Larvae. J. Insect Sci. 2021, 21, ieaa145. [Google Scholar] [CrossRef] [PubMed]
- Dreassi, E.; Cito, A.; Zanfini, A.; Materozzi, L.; Botta, M.; Francardi, V. Dietary fatty acids influence the growth and fatty acid composition of the yellow mealworm Tenebrio molitor (Coleoptera: Tenebrionidae). Lipids 2017, 52, 285–294. [Google Scholar] [CrossRef]
- Scriber, J.M. Host-Plant Suitability. In Chemical Ecology of Insects; Springer: Boston, MA, USA, 1984; pp. 159–202. [Google Scholar]
- Woods, H.A.; Chamberlin, M.E. Effects of dietary protein concentration on L-proline transport by Manduca sexta midgut. J. Insect Physiol. 1999, 45, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Raubenheimer, D.; Simpson, S.J. The geometry of compensatory feeding in the locust. Anim. Behav. 1993, 45, 953–964. [Google Scholar] [CrossRef] [Green Version]
- Raubenheimer, D.; Simpson, S.J. Integrative models of nutrient balancing: Application to insects and vertebrates. Nutr. Res. Rev. 1997, 10, 151–179. [Google Scholar] [CrossRef] [Green Version]
- Woods, H.A. Patterns and mechanisms of growth of fifth-instar Manduca sexta caterpillars following exposure to low- or high-protein food during early instars. Physiol. Biochem. Zool. PBZ 1999, 72, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Kröncke, N.; Benning, R. Self-Selection of Feeding Substrates by Tenebrio molitor Larvae of Different Ages to Determine Optimal Macronutrient Intake and the Influence on Larval Growth and Protein Content. Insects 2022, 13, 657. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Rojas, M.G.; Shapiro-Ilan, D.I.; Tedders, W.L. Self-selection of two diet components by Tenebrio molitor (Coleoptera: Tenebrionidae) larvae and its impact on fitness. Environ. Entomol. 2011, 40, 1285–1294. [Google Scholar] [CrossRef] [Green Version]
- Morales-Ramos, J.A.; Rojas, M.G.; Kelstrup, H.C.; Emery, V. Self-Selection of Agricultural By-Products and Food Ingredients by Tenebrio molitor (Coleoptera: Tenebrionidae) and Impact on Food Utilization and Nutrient Intake. Insects 2020, 11, 827. [Google Scholar] [CrossRef]
- Alves, A.V.; Sanjinez-Argandoña, E.J.; Linzmeier, A.M.; Cardoso, C.A.L.; Macedo, M.L.R. Food Value of Mealworm Grown on Acrocomia aculeata Pulp Flour. PloS ONE 2016, 11, e0151275. [Google Scholar] [CrossRef]
- Harsányi, E.; Juhász, C.; Kovács, E.; Huzsvai, L.; Pintér, R.; Fekete, G.; Varga, Z.I.; Aleksza, L.; Gyuricza, C. Evaluation of Organic Wastes as Substrates for Rearing Zophobas morio, Tenebrio molitor, and Acheta domesticus Larvae as Alternative Feed Supplements. Insects 2020, 11, 604. [Google Scholar] [CrossRef] [PubMed]
- Behmer, S.T. Insect herbivore nutrient regulation. Annu. Rev. Entomol. 2009, 54, 165–187. [Google Scholar] [CrossRef]
- Joern, A.; Behmer, S.T. Importance of dietary nitrogen and carbohydrates to survival, growth, and reproduction in adults of the grasshopper Ageneotettix deorum (Orthoptera: Acrididae). Oecologia 1997, 112, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Blanco, C.A.; Portilla, M.; Abel, C.A.; Winters, H.; Ford, R.; Streett, D. Soybean flour and wheat germ proportions in artificial diet and their effect on the growth rates of the tobacco budworm, Heliothis virescens. J. Insect Sci. 2009, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fast, P.G. Insect lipids. Prog. Chem. Fats Other Lipids 1970, 11, 181–242. [Google Scholar]
- Finkel, A.J. The lipid composition of Tenebrio molitor larvae. Physiol. Zool. 1948, 21, 111–133. [Google Scholar] [CrossRef]
- Pierce, M.M.; Wehling, R.L. Comparison of sample handling and data treatment methods for determining moisture and fat in Cheddar cheese by near-infrared spectroscopy. J. Agric. Food Chem. 1994, 42, 2830–2835. [Google Scholar] [CrossRef]
- Kobayashi, K.-I.; Matsui, Y.; Maebuchi, Y.; Toyota, T.; Nakauchi, S. Near Infrared Spectroscopy and Hyperspectral Imaging for Prediction and Visualisation of Fat and Fatty Acid Content in Intact Raw Beef Cuts. J. Near Infrared Spectrosc. 2010, 18, 301–315. [Google Scholar] [CrossRef]
- Davrieux, F.; Allal, F.; Piombo, G.; Kelly, B.; Okulo, J.B.; Thiam, M.; Diallo, O.B.; Bouvet, J.-M. Near infrared spectroscopy for high-throughput characterization of Shea tree (Vitellaria paradoxa) nut fat profiles. J. Agric. Food Chem. 2010, 58, 7811–7819. [Google Scholar] [CrossRef]
- Benes, E.; Biró, B.; Fodor, M.; Gere, A. Analysis of wheat flour-insect powder mixtures based on their near infrared spectra. Food Chem. X 2022, 13, 100266. [Google Scholar] [CrossRef]
- Cozzolino, D.; Murray, I. Effect of Sample Presentation and Animal Muscle Species on the Analysis of Meat by near Infrared Reflectance Spectroscopy. J. Near Infrared Spectrosc. 2002, 10, 37–44. [Google Scholar] [CrossRef]
- Milliken, R.E.; Mustard, J.F. Quantifying absolute water content of minerals using near-infrared reflectance spectroscopy. J. Geophys. Res. 2005, 110, 2534. [Google Scholar] [CrossRef] [Green Version]
- Tkachuk, R. Oil and protein analysis of whole rapeseed kernels by near infrared reflectance spectroscopy. J. Amer. Oil Chem. Soc. 1981, 58, 819–822. [Google Scholar] [CrossRef]
- Rødbotten, R.; Nilsen, B.; Hildrum, K. Prediction of beef quality attributes from early post mortem near infrared reflectance spectra. Food Chem. 2000, 69, 427–436. [Google Scholar] [CrossRef]
- Ripoll, G.; Failla, S.; Panea, B.; Hocquette, J.-F.; Dunner, S.; Olleta, J.L.; Christensen, M.; Ertbjerg, P.; Richardson, I.; Contò, M.; et al. Near-Infrared Reflectance Spectroscopy for Predicting the Phospholipid Fraction and the Total Fatty Acid Composition of Freeze-Dried Beef. Sensors 2021, 21, 4230. [Google Scholar] [CrossRef]
- Kovalenko, I.V.; Rippke, G.R.; Hurburgh, C.R. Measurement of soybean fatty acids by near-infrared spectroscopy: Linear and nonlinear calibration methods. J. Amer. Oil Chem. Soc. 2006, 83, 421–427. [Google Scholar] [CrossRef]
- Ng, W.; Minasny, B.; Mendes, W.d.S.; Demattê, J.A.M. The influence of training sample size on the accuracy of deep learning models for the prediction of soil properties with near-infrared spectroscopy data. Soil 2020, 6, 565–578. [Google Scholar] [CrossRef]
- de Marchi, M.; Riovanto, R.; Penasa, M.; Cassandro, M. At-line prediction of fatty acid profile in chicken breast using near infrared reflectance spectroscopy. Meat Sci. 2012, 90, 653–657. [Google Scholar] [CrossRef]
- Fernández-Cabanás, V.M.; Polvillo, O.; Rodríguez-Acuña, R.; Botella, B.; Horcada, A. Rapid determination of the fatty acid profile in pork dry-cured sausages by NIR spectroscopy. Food Chem. 2011, 124, 373–378. [Google Scholar] [CrossRef]
- Cozzolino, D.; Murray, I.; Chree, A.; Scaife, J.R. Multivariate determination of free fatty acids and moisture in fish oils by partial least-squares regression and near-infrared spectroscopy. LWT Food Sci. Technol. 2005, 38, 821–828. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Substrate Amount (%) | ||||||
|---|---|---|---|---|---|---|---|
| Coconut Flour (CF) | Flaxseed Flour (FSF) | Rose Hip Hulls (RHH) | Grape Pomace (GP) | Hemp Protein Flour (HPF) | Pea Protein Flour (PPF) | Wheat Bran (WB) | |
| CF5 | 3.4 | - | - | - | - | - | 96.6 |
| CF10 | 23.8 | - | - | - | - | - | 76.2 |
| CF15 | 46.2 | - | - | - | - | - | 53.8 |
| CF20 | 68.6 | - | - | - | - | - | 31.4 |
| FSF5 | - | 1.1 | - | - | - | - | 98.9 |
| FSF10 | - | 18.7 | - | - | - | - | 81.3 |
| FSF15 | - | 36.4 | - | - | - | - | 63.6 |
| FSF20 | - | 54.1 | - | - | - | - | 45.9 |
| GP4 | - | - | - | 42.5 | - | - | 57.5 |
| HPF5 | - | - | - | - | 14.5 | - | 85.5 |
| HPF8 | - | - | - | - | 71.5 | - | 28.5 |
| RHH4 | - | - | 36.4 | - | - | - | 63.6 |
| PPF5 | - | - | - | - | - | 7.8 | 92.2 |
| PPF6 | - | - | - | - | - | 38.6 | 61.4 |
| WB (Control) | - | - | - | - | - | - | 100.0 |
| Substrate | Moisture (%) | Protein (% FW) | Fat (% FW) | Carbohydrate (% FW) | Fiber (% FW) | Ash (% FW) |
|---|---|---|---|---|---|---|
| CF5 | 12.0 | 14.9 | 5.0 | 44.9 | 17.6 | 5.6 |
| CF10 | 11.2 | 15.2 | 10.0 | 43.3 | 15.6 | 4.8 |
| CF15 | 10.4 | 15.4 | 15.0 | 41.8 | 13.5 | 3.9 |
| CF20 | 9.6 | 15.7 | 20.0 | 40.2 | 11.5 | 3.0 |
| FSF5 | 12.0 | 15.0 | 5.0 | 44.8 | 17.6 | 5.7 |
| FSF10 | 11.2 | 16.4 | 10.0 | 41.3 | 15.6 | 5.5 |
| FSF15 | 10.5 | 17.8 | 15.0 | 37.7 | 13.7 | 5.3 |
| FSF20 | 9.8 | 19.2 | 20.0 | 34.2 | 11.7 | 5.1 |
| GP4 | 11.0 | 12.0 | 4.4 | 51.0 | 16.7 | 5.0 |
| HPF5 | 11.6 | 20.0 | 5.3 | 39.6 | 18.0 | 5.5 |
| HPF8 | 9.9 | 40.0 | 7.8 | 18.4 | 19.3 | 4.6 |
| RHH4 | 12.0 | 10.8 | 3.5 | 51.0 | 17.0 | 5.7 |
| PPF5 | 11.3 | 20.0 | 5.0 | 41.9 | 16.6 | 5.3 |
| PPF6 | 8.4 | 40.0 | 6.0 | 29.5 | 12.5 | 3.6 |
| WB (Control) | 12.0 | 14.9 | 4.7 | 45.0 | 17.7 | 5.7 |
| Fatty Acid | Group | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Start | CF5 | CF10 | CF15 | CF20 | FSF5 | FSF10 | FSF15 | FSF20 | GP4 | HPF5 | HPF8 | RHH4 | PPF5 | PPF6 | WB | |
| Lauric acid (C12:0) | n.d. | 0.7 ± 0.1 | 4.6 ± 0.1 | 4.7 ± 0.1 | 3.2 ± 0.1 | 0.6 ± 0.0 | 0.6 ± 0.0 | n.d. | 0.5 ± 0.1 | 0.7 ± 0.1 | 0.7 ± 0.1 | 0.9 ± 0.1 | 0.7 ± 0.0 | 1.0 ± 0.1 | n.d. | 0.8 ± 0.1 |
| Myristic acid (C14:0) | 3.3 ± 0.0 | 4.8 ± 0.1 | 12.2 ± 0.7 | 12.9 ± 0.7 | 11.4 ± 0.4 | 3.3 ± 0.1 | 4.4 ± 0.1 | n.d. | 4.2 ± 0.1 | 6.4 ± 0.6 | 5.0 ± 0.6 | 4.4 ± 0.1 | 6.1 ± 0.5 | 4.4 ± 0.1 | 5.6 ± 0.4 | 3.8 ± 0.2 |
| Palmitic acid (C16:0) | 23.0 ± 2.6 | 22.2 ± 0.5 | 21.4 ± 0.2 | 20.1 ± 0.0 | 19.2 ± 0.1 | 19.3 ± 0.2 | 19.3 ± 0.7 | 22.2 ± 0.8 | 19.5 ± 0.5 | 27.1 ± 0.6 | 22.5 ± 0.7 | 19.2 ± 0.3 | 27.1 ± 0.5 | 23.1 ± 0.2 | 21.4 ± 0.2 | 21.7 ± 1.0 |
| Palmitoleic acid (C16:1) | 1.4 ± 0.5 | 1.6 ± 0.1 | 2.4 ± 0.1 | 1.5 ± 0.0 | 1.7 ± 0.1 | 2.1 ± 0.1 | 1.9 ± 0.0 | 1.4 ± 0.1 | 1.8 ± 0.1 | 2.3 ± 0.1 | 2.2 ± 0.1 | 2.5 ± 0.1 | 2.0 ± 0.1 | 2.3 ± 0.1 | 2.1 ± 0.1 | 1.6 ± 0.0 |
| Stearic acid (C18:0) | 4.7 ± 1.2 | 2.9 ± 0.0 | 3.1 ± 0.1 | 2.7 ± 0.1 | 3.2 ± 0.1 | 3.5 ± 0.0 | 3.4 ± 0.0 | 4.2 ± 0.0 | 4.0 ± 0.0 | 2.9 ± 0.1 | 3.4 ± 0.1 | 2.9 ± 0.1 | 2.8 ± 0.1 | 3.0 ± 0.0 | 6.7 ± 0.2 | 3.1 ± 0.1 |
| Oleic acid (C18:1 ω9) | 31.3 ± 4.9 | 39.8 ± 0.4 | 36.1 ± 0.5 | 41.2 ± 0.4 | 44.8 ± 0.1 | 43.1 ± 0.7 | 40.8 ± 0.2 | 37.1 ± 0.0 | 40.3 ± 0.7 | 36.8 ± 0.7 | 34.7 ± 0.6 | 35.7 ± 0.3 | 34.4 ± 0.7 | 35.0 ± 0.4 | 30.5 ± 0.2 | 37.4 ± 0.6 |
| Linoleic acid (C18:2 ω6) | 34.8 ± 6.3 | 27.2 ± 0.3 | 19.1 ± 0.4 | 16.5 ± 0.6 | 16.3 ± 0.6 | 25.9 ± 0.2 | 21.3 ± 0.3 | 22.1 ± 0.5 | 19.2 ± 0.4 | 22.7 ± 0.6 | 29.5 ± 0.6 | 30.8 ± 0.1 | 25.3 ± 0.3 | 29.8 ± 0.1 | 32.1 ± 0.1 | 30.3 ± 0.4 |
| α-Linolenic acid (C18:3 ω3) | 1.4 ± 0.6 | 0.9 ± 0.1 | 1.0 ± 0.1 | 0.5 ± 0.1 | 0.2 ± 0.1 | 2.1 ± 0.1 | 8.4 ± 0.7 | 13.0 ± 0.3 | 10.5 ± 0.0 | 1.1 ± 0.0 | 2.0 ± 0.0 | 3.8 ± 0.0 | 1.4 ± 0.1 | 1.4 ± 0.0 | 1.7 ± 0.1 | 1.3 ± 0.1 |
| ∑ SFA | 31.0 ± 1.4 | 30.6 ± 0.7 | 41.4 ± 0.9 | 40.4 ± 0.9 | 40.0 ± 0.4 | 26.8 ± 0.2 | 27.6 ± 0.8 | 26.3 ± 0.7 | 28.2 ± 0.4 | 36.2 ± 1.4 | 31.6 ± 1.3 | 27.3 ± 0.3 | 36.9 ± 0.9 | 31.6 ± 1.3 | 33.6 ± 0.4 | 29.3 ± 1.1 |
| ∑ MUFA | 32.8 ± 5.4 | 41.4 ± 0.5 | 38.6 ± 0.6 | 42.7 ± 0.4 | 46.5 ± 0.1 | 45.2 ± 0.5 | 42.7 ± 0.2 | 38.5 ± 0.1 | 42.1 ± 0.8 | 39.1 ± 0.8 | 36.4 ± 0.7 | 38.2 ± 0.4 | 36.4 ± 0.5 | 36.9 ± 0.7 | 32.6 ± 0.1 | 39.1 ± 0.6 |
| ∑ PUFA | 36.2 ± 6.8 | 28.0 ± 0.1 | 20.1 ± 0.3 | 16.9 ± 0.5 | 16.6 ± 0.5 | 28.0 ± 0.3 | 29.7 ± 1.0 | 35.1 ± 0.8 | 29.7 ± 0.4 | 23.8 ± 0.6 | 31.5 ± 0.6 | 34.5 ± 0.1 | 26.7 ± 0.3 | 31.5 ± 0.6 | 33.8 ± 0.3 | 31.7 ± 0.5 |
| Statistics | Calibration Set | Validation Set |
|---|---|---|
| Fat (g/100 g of FW) | Fat (g/100 g of FW) | |
| Mean | 12.0 | 12.1 |
| Minimum | 7.4 | 7.3 |
| Maximum | 16.2 | 16.2 |
| SD | 2.3 | 2.3 |
| Item | Mathematical Treatment | No. of Latent Variables | Calibration Set | Validation Set | |||
|---|---|---|---|---|---|---|---|
| R2C | RMSEC | R2P | RMSEP | RPD | |||
| Fat | None | 6 | 0.955 | 0.482 | 0.967 | 0.431 | 5.34 |
| MSC | 5 | 0.962 | 0.443 | 0.969 | 0.412 | 5.58 | |
| MC | 6 | 0.975 | 0.355 | 0.986 | 0.276 | 8.33 | |
| 1D | 4 | 0.949 | 0.512 | 0.954 | 0.502 | 4.58 | |
| 2D | 5 | 0.958 | 0.461 | 0.961 | 0.467 | 4.93 | |
| Fatty Acid (% of DW) | Calibration Set | Validation Set | ||||||
|---|---|---|---|---|---|---|---|---|
| Minimum | Maximum | Mean | SD | Minimum | Maximum | Mean | SD | |
| Lauric acid (C12:0) | 0.4 | 4.7 | 1.2 | 1.5 | 0.3 | 4.9 | 1.1 | 1.4 |
| Myristic acid (C14:0) | 3.2 | 13.4 | 5.8 | 3.4 | 3.3 | 13.0 | 5.6 | 3.3 |
| Palmitic acid (C16:0) | 18.8 | 27.6 | 21.8 | 2.5 | 18.5 | 27.8 | 22.0 | 2.7 |
| Palmitoleic acid (C16:1) | 1.1 | 2.5 | 1.9 | 0.4 | 1.3 | 2.4 | 1.8 | 0.4 |
| Stearic acid (C18:0) | 2.6 | 6.8 | 3.5 | 1.0 | 2.9 | 6.5 | 3.6 | 1.0 |
| Oleic acid (C18:1 ω9) | 27.9 | 44.8 | 37.5 | 4.0 | 28.7 | 45.2 | 37.3 | 4.2 |
| Linoleic acid (C18:2 ω6) | 15.9 | 39.3 | 25.3 | 5.9 | 16.0 | 35.4 | 25.1 | 5.6 |
| α-Linolenic acid (C18:3 ω3) | 0.1 | 13.2 | 3.2 | 3.8 | 0.3 | 13.5 | 3.3 | 4.0 |
| SFA | 25.8 | 42.0 | 32.4 | 4.8 | 26.2 | 41.6 | 32.2 | 4.6 |
| MUFA | 28.6 | 46.6 | 39.5 | 4.1 | 30.5 | 46.3 | 39.4 | 4.2 |
| PUFA | 16.2 | 41.1 | 28.2 | 6.2 | 16.4 | 40.2 | 28.5 | 6.4 |
| Fatty Acid | Mathematical Treatment | No. of Latent Variables | Calibration Set | Validation Set | ||||
|---|---|---|---|---|---|---|---|---|
| R2C | RMSEC | R2P | RMSEP | RPD | R2F | |||
| Lauric acid (C12:0) | None | 8 | 0.915 | 0.418 | 0.917 | 0.375 | 3.73 | 0.253 |
| Myristic acid (C14:0) | MSC | 8 | 0.947 | 0.773 | 0.930 | 0.803 | 4.11 | 0.331 |
| Palmitic acid (C16:0) | MSC + Detrend | 8 | 0.812 | 1.077 | 0.877 | 1.012 | 2.66 | 0.269 |
| Palmitoleic acid (C16:1) | Detrend | 8 | 0.337 | 0.290 | 0.345 | 0.255 | 1.57 | 0.156 |
| Stearic acid (C18:0) | 2D | 8 | 0.579 | 0.638 | 0.510 | 0.509 | 1.96 | 0.071 |
| Oleic acid (C18:1 ω9) | MSC | 8 | 0.922 | 1.104 | 0.949 | 0.756 | 5.55 | 0.527 |
| Linoleic acid (C18:2 ω6) | MSC | 8 | 0.925 | 1.602 | 0.931 | 1.411 | 3.98 | 0.568 |
| α-Linolenic acid (C18:3 ω3) | MSC | 8 | 0.964 | 0.713 | 0.945 | 0.817 | 4.90 | 0.019 |
| SFA | MSC | 8 | 0.948 | 1.088 | 0.942 | 1.081 | 4.26 | 0.091 |
| MUFA | MSC | 8 | 0.886 | 1.371 | 0.903 | 1.050 | 4.00 | 0.511 |
| PUFA | MSC | 8 | 0.943 | 1.466 | 0.878 | 2.115 | 3.03 | 0.282 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kröncke, N.; Neumeister, M.; Benning, R. Near-Infrared Reflectance Spectroscopy for Quantitative Analysis of Fat and Fatty Acid Content in Living Tenebrio molitor Larvae to Detect the Influence of Substrate on Larval Composition. Insects 2023, 14, 114. https://doi.org/10.3390/insects14020114
Kröncke N, Neumeister M, Benning R. Near-Infrared Reflectance Spectroscopy for Quantitative Analysis of Fat and Fatty Acid Content in Living Tenebrio molitor Larvae to Detect the Influence of Substrate on Larval Composition. Insects. 2023; 14(2):114. https://doi.org/10.3390/insects14020114
Chicago/Turabian StyleKröncke, Nina, Monique Neumeister, and Rainer Benning. 2023. "Near-Infrared Reflectance Spectroscopy for Quantitative Analysis of Fat and Fatty Acid Content in Living Tenebrio molitor Larvae to Detect the Influence of Substrate on Larval Composition" Insects 14, no. 2: 114. https://doi.org/10.3390/insects14020114