
Global Distribution of Aedes aegypti and Aedes albopictus in a Climate Change Scenario of Regional Rivalry

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Description
2.2. Data Preparation
2.3. Environmental Data
2.4. Model Building, Fitting, and Evaluation
2.5. Spatial Prediction
3. Results
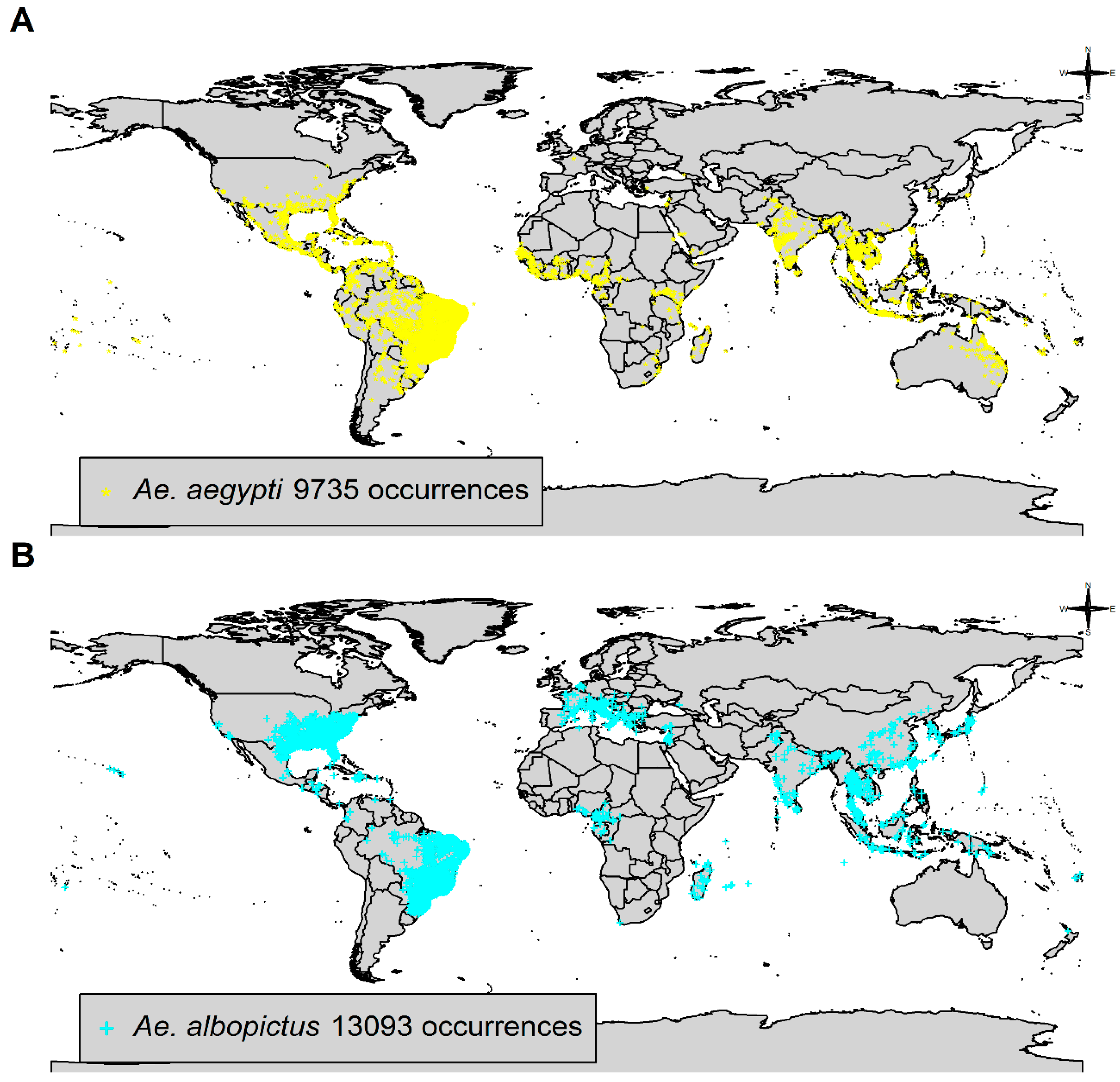
3.1. Species Occurrence Data
3.2. Environmental Data
3.3. Boosted Regression Tree Model
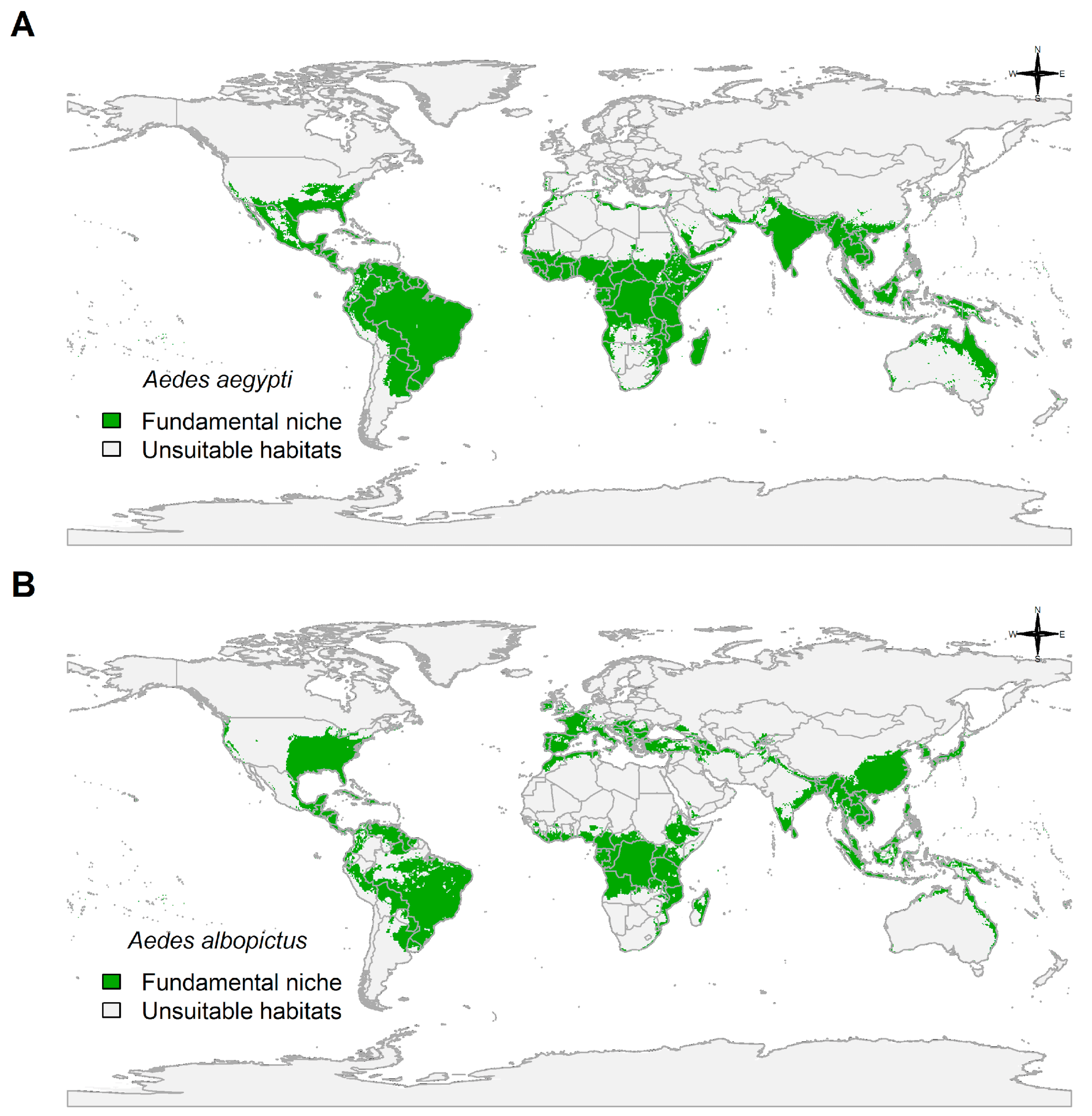
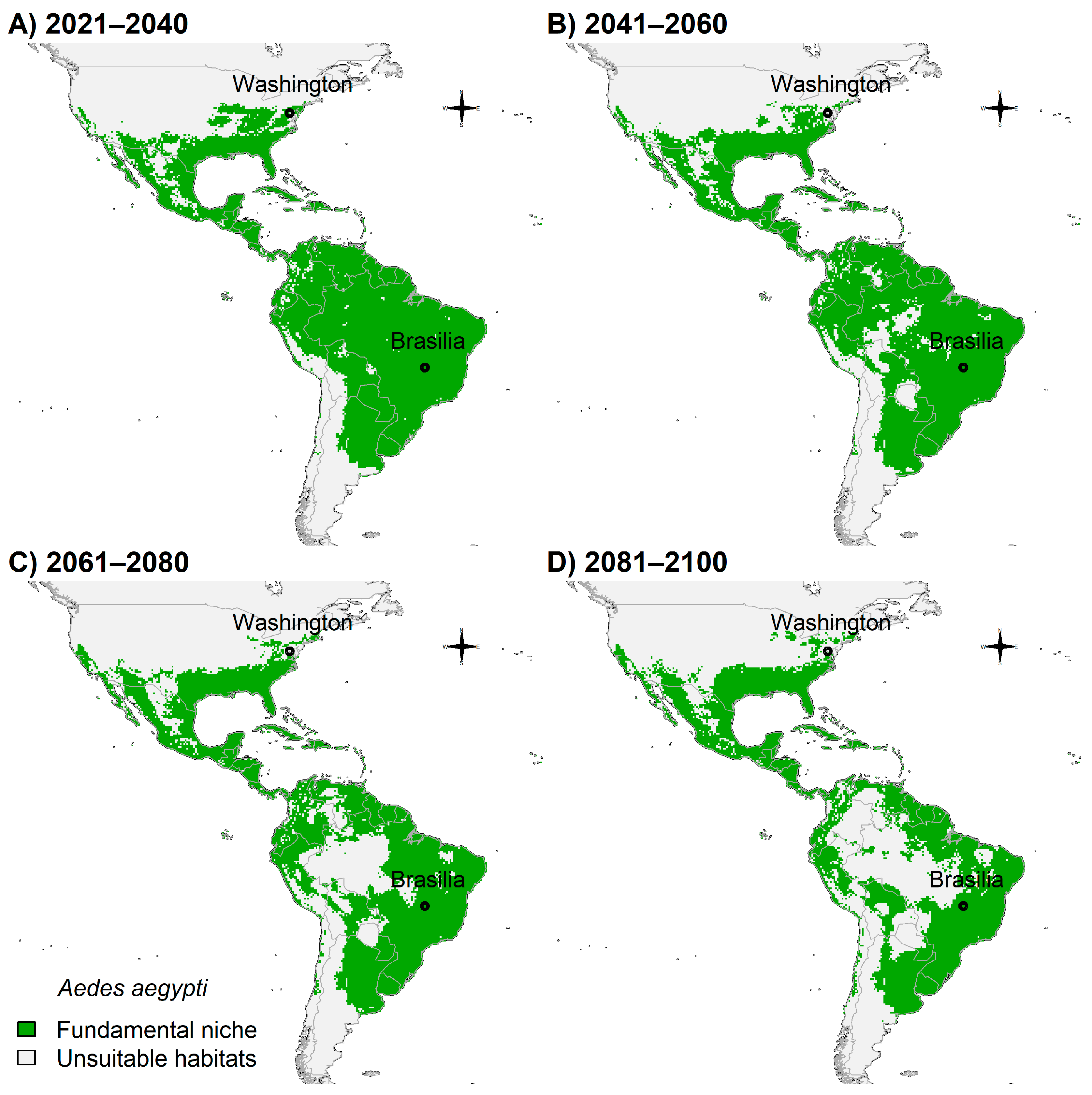
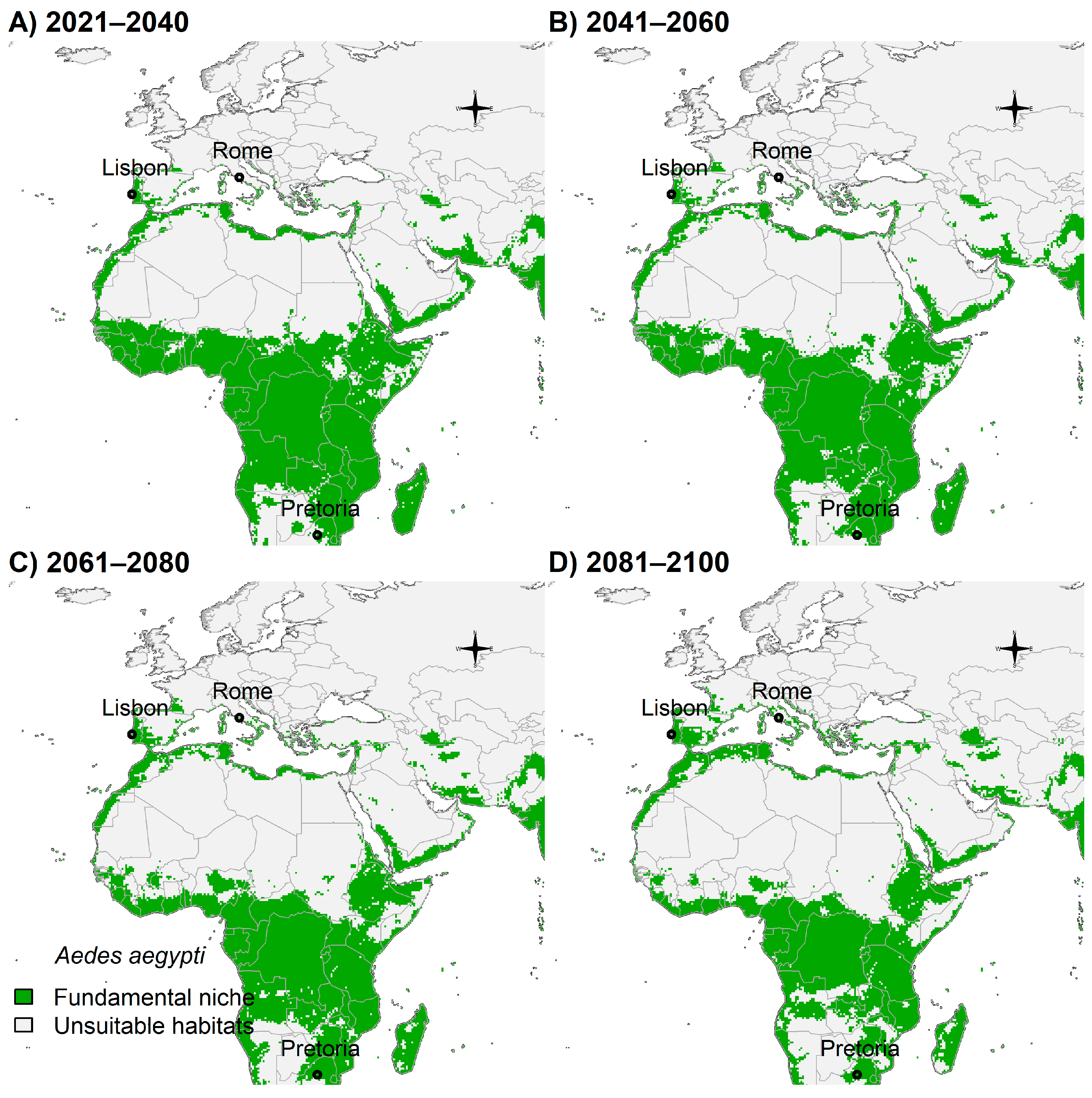
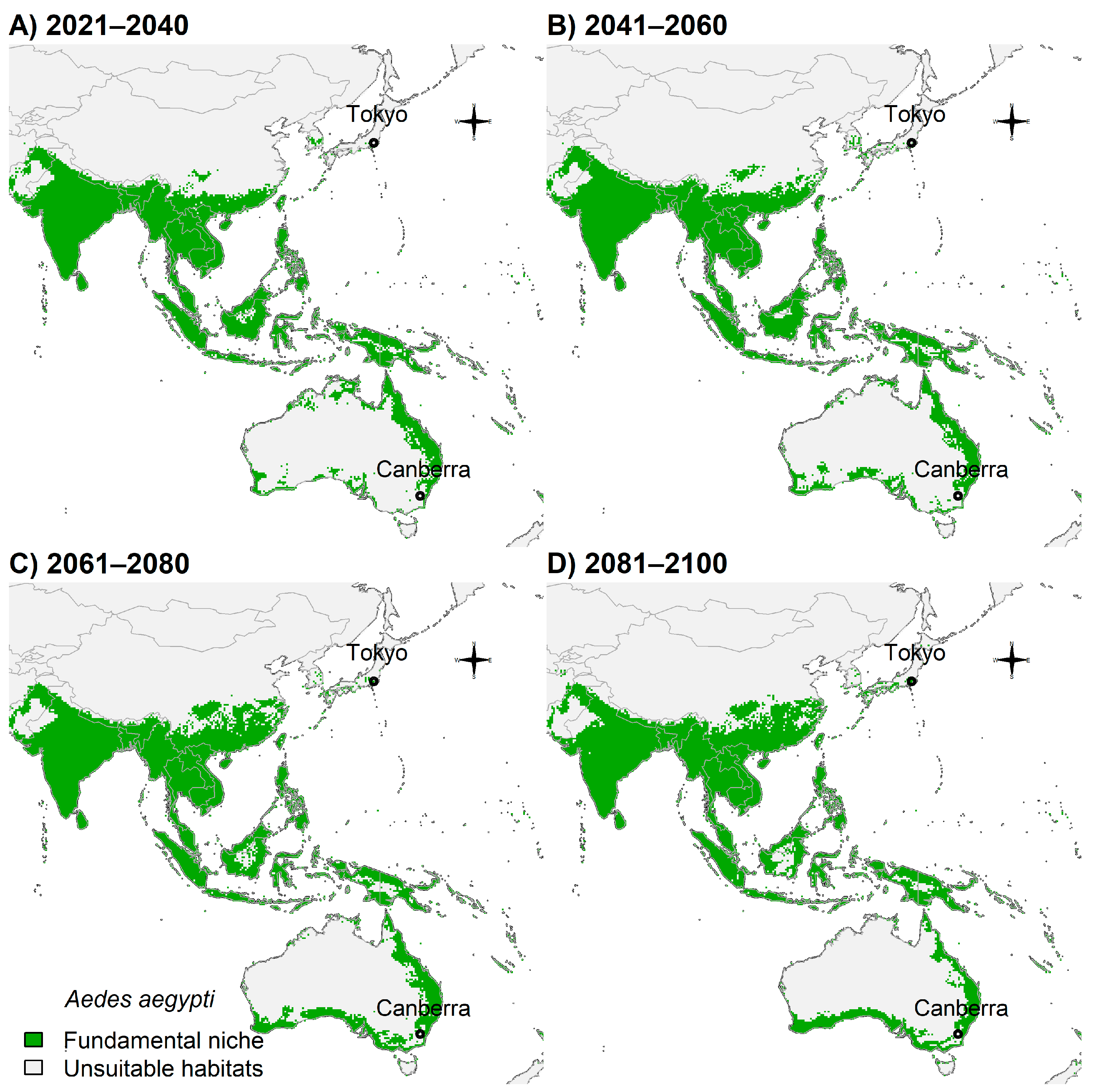
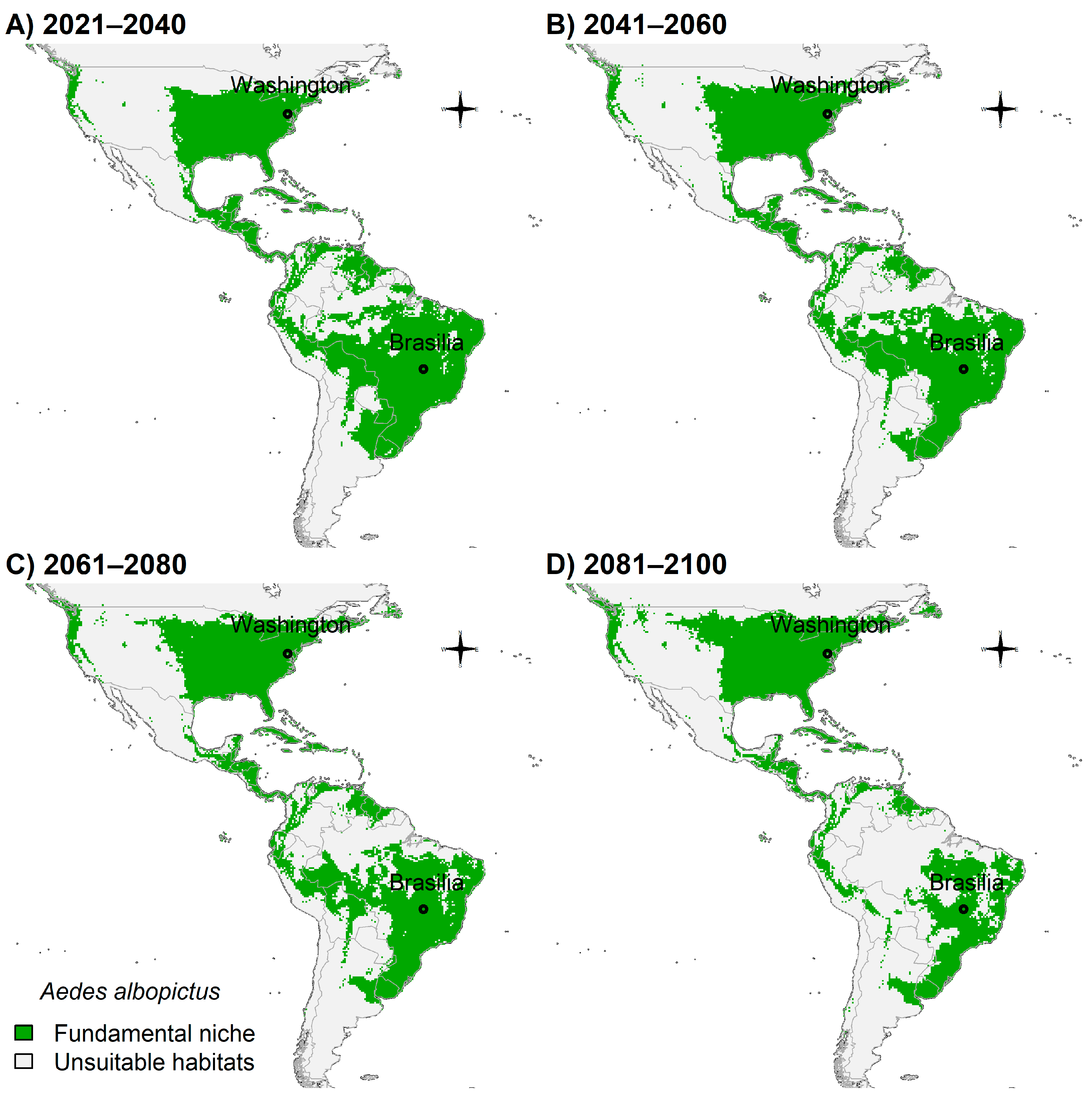
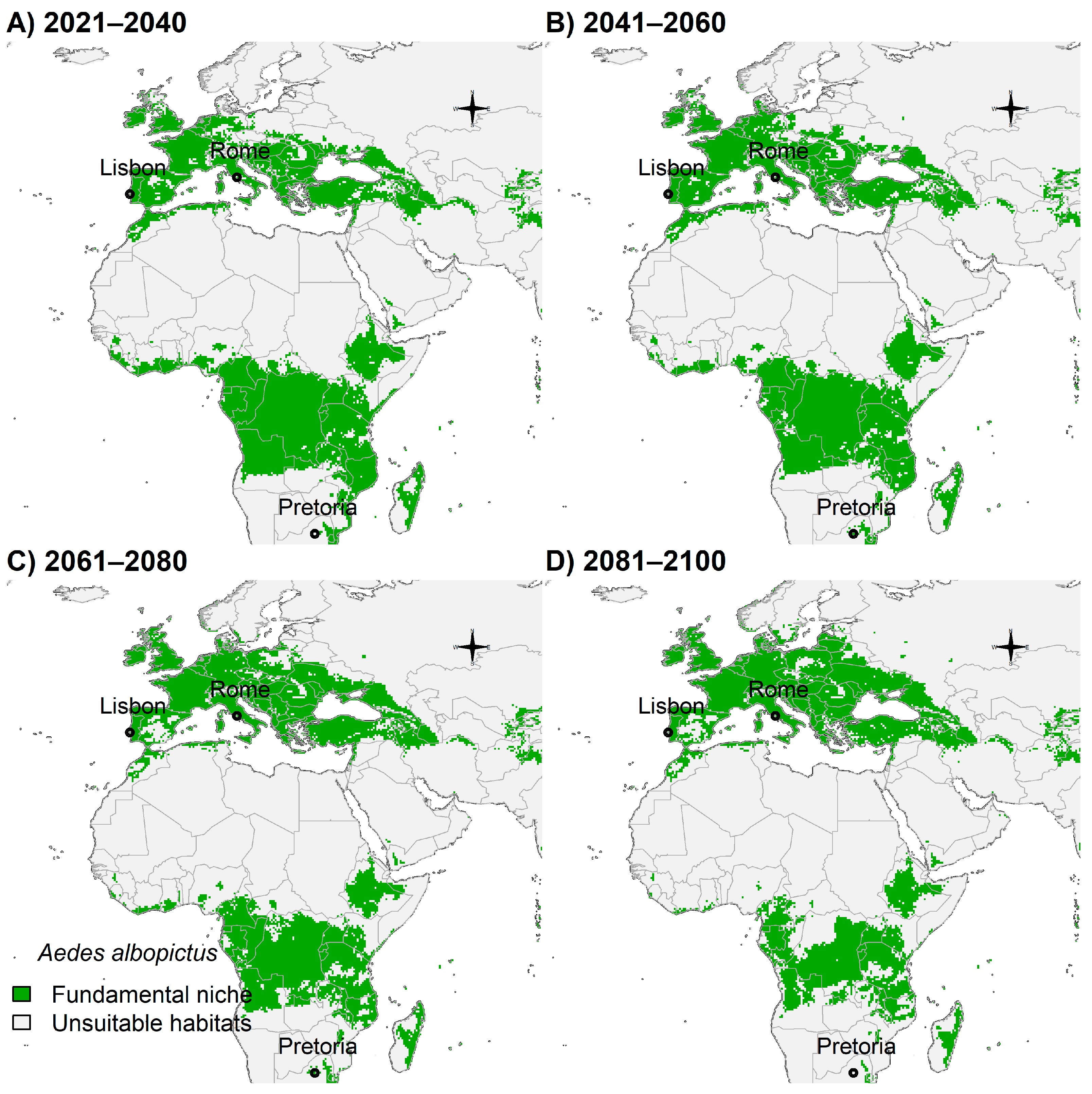
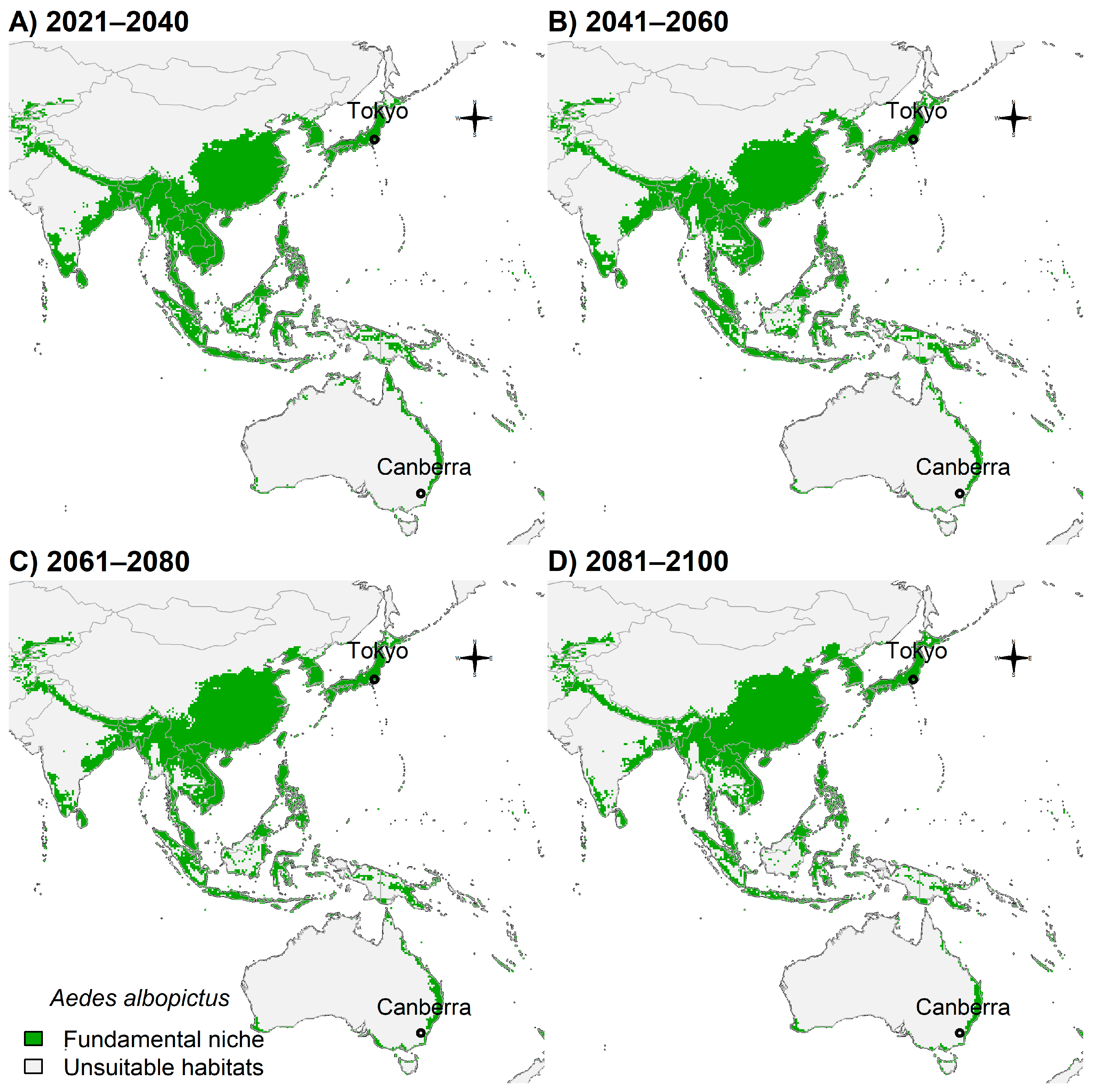
3.4. Fundamental Niche and Potential Future Distributions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.N.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The Global Distribution of the Arbovirus Vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.; Shearer, F.M.; Brady, O.J.; Messina, J.P.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; et al. The Global Compendium of Aedes aegypti and Ae. albopictus Occurrence. Sci. Data 2015, 2, 150035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lwande, O.W.; Obanda, V.; Lindström, A.; Ahlm, C.; Evander, M.; Näslund, J.; Bucht, G. Globe-Trotting Aedes aegypti and Aedes albopictus: Risk Factors for Arbovirus Pandemics. Vector Borne Zoonotic Dis. 2020, 20, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, W.K.; Cortez-Escalante, J.; De Oliveira, W.T.G.H.; do Carmo, G.M.I.; Henriques, C.M.P.; Coelho, G.E.; Araújo de França, G.V. Increase in Reported Prevalence of Microcephaly in Infants Born to Women Living in Areas with Confirmed Zika Virus Transmission During the First Trimester of Pregnancy—Brazil, 2015. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 242–247. [Google Scholar] [CrossRef]
- Girard, M.; Nelson, C.B.; Picot, V.; Gubler, D.J. Arboviruses: A Global Public Health Threat. Vaccine 2020, 38, 3989–3994. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The Global Distribution and Burden of Dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [Green Version]
- Brady, O.J.; Hay, S.I. The Global Expansion of Dengue: How Aedes aegypti Mosquitoes Enabled the First Pandemic Arbovirus. Annu. Rev. Entomol. 2020, 65, 191–208. [Google Scholar] [CrossRef] [Green Version]
- Näslund, J.; Ahlm, C.; Islam, K.; Evander, M.; Bucht, G.; Lwande, O.W. Emerging Mosquito-Borne Viruses Linked to Aedes aegypti and Aedes albopictus: Global Status and Preventive Strategies. Vector Borne Zoonotic Dis. 2021, 21, 731–746. [Google Scholar] [CrossRef]
- Iwamura, T.; Guzman-Holst, A.; Murray, K.A. Accelerating Invasion Potential of Disease Vector Aedes aegypti under Climate Change. Nat. Commun. 2020, 11, 2130. [Google Scholar] [CrossRef]
- Khormi, H.M.; Kumar, L. Climate Change and the Potential Global Distribution of Aedes aegypti: Spatial Modelling Using GIS and CLIMEX. Geospat. Health 2014, 8, 405–415. [Google Scholar] [CrossRef]
- Rochlin, I.; Ninivaggi, D.V.; Hutchinson, M.L.; Farajollahi, A. Climate Change and Range Expansion of the Asian Tiger Mosquito (Aedes albopictus) in Northeastern USA: Implications for Public Health Practitioners. PLoS ONE 2013, 8, e60874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenzweig, C. Global Climate Change: Predictions and Observations. Am. J. Agric. Econ. 1989, 71, 1265–1271. [Google Scholar] [CrossRef]
- Juliano, S.A.; O’Meara, G.F.; Morrill, J.R.; Cutwa, M.M. Desiccation and Thermal Tolerance of Eggs and the Coexistence of Competing Mosquitoes. Oecologia 2002, 130, 458–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, K.L.; Gómez Martínez, C.; Almanza, A.; Rovira, J.R.; McMillan, W.O.; Enriquez, V.; Barraza, E.; Diaz, M.; Sanchez-Galan, J.E.; Whiteman, A.; et al. High Infestation of Invasive Aedes Mosquitoes in Used Tires along the Local Transport Network of Panama. Parasit. Vectors 2019, 12, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, D.H.; Wilkerson, R.C.; Birney, I.; Harrison, S.; Christensen, J.; Rueda, L.M. MosquitoMap and the Mal-Area Calculator: New Web Tools to Relate Mosquito Species Distribution with Vector Borne Disease. Int. J. Health Geogr. 2010, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkerson, R.C.; Linton, Y.-M.; Strickman, D. Mosquitoes of the World; Johns Hopkins University Press: Baltimore, MD, USA, 2020; ISBN 978-1-4214-3814-6. [Google Scholar]
- Crawford, J.E.; Alves, J.M.; Palmer, W.J.; Day, J.P.; Sylla, M.; Ramasamy, R.; Surendran, S.N.; Black, W.C.; Pain, A.; Jiggins, F.M. Population Genomics Reveals That an Anthropophilic Population of Aedes aegypti Mosquitoes in West Africa Recently Gave Rise to American and Asian Populations of This Major Disease Vector. BMC Biol. 2017, 15, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.E.; McBride, C.S.; Johnson, P.; Ritchie, S.; Paupy, C.; Bossin, H.; Lutomiah, J.; Fernandez-Salas, I.; Ponlawat, A.; Cornel, A.J.; et al. Worldwide Patterns of Genetic Differentiation Imply Multiple “domestications” of Aedes aegypti, a Major Vector of Human Diseases. Proc. Biol. Sci. 2011, 278, 2446–2454. [Google Scholar] [CrossRef] [Green Version]
- Chadee, D.D.; Martinez, R. Aedes aegypti (L.) in Latin American and Caribbean Region: With Growing Evidence for Vector Adaptation to Climate Change? Acta Trop. 2016, 156, 137–143. [Google Scholar] [CrossRef]
- Chretien, J.-P.; Anyamba, A.; Bedno, S.A.; Breiman, R.F.; Sang, R.; Sergon, K.; Powers, A.M.; Onyango, C.O.; Small, J.; Tucker, C.J.; et al. Drought-Associated Chikungunya Emergence along Coastal East Africa. Am. J. Trop. Med. Hyg. 2007, 76, 405–407. [Google Scholar] [CrossRef] [Green Version]
- Cunze, S.; Kochmann, J.; Koch, L.K.; Klimpel, S. Aedes albopictus and Its Environmental Limits in Europe. PLoS ONE 2016, 11, e0162116. [Google Scholar] [CrossRef]
- Farjana, T.; Tuno, N.; Higa, Y. Effects of Temperature and Diet on Development and Interspecies Competition in Aedes aegypti and Aedes Albopictus. Med. Vet. Entomol. 2012, 26, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.U.; Ogden, N.H.; Fazil, A.A.; Gachon, P.H.; Dueymes, G.U.; Greer, A.L.; Ng, V. Current and Projected Distributions of Aedes Aegypti and Ae. albopictus in Canada and the U.S. Environ. Health Perspect. 2020, 128, 57007. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.H.; Milka, R.; Caminade, C.; Gachon, P. Recent and Projected Future Climatic Suitability of North America for the Asian Tiger Mosquito xi. Parasit. Vectors 2014, 7, 532. [Google Scholar] [CrossRef]
- Hill, M.P.; Axford, J.K.; Hoffmann, A.A. Predicting the Spread of Aedes albopictus in Australia under Current and Future Climates: Multiple Approaches and Datasets to Incorporate Potential Evolutionary Divergence: Invasion Potential of A. albopictus in Australia. Austral Ecol. 2014, 39, 469–478. [Google Scholar] [CrossRef]
- Kamal, M.; Kenawy, M.A.; Rady, M.H.; Khaled, A.S.; Samy, A.M. Mapping the Global Potential Distributions of Two Arboviral Vectors Aedes aegypti and Ae. albopictus under Changing Climate. PLoS ONE 2018, 13, e0210122. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; in press. [Google Scholar]
- Fujimori, S.; Hasegawa, T.; Masui, T.; Takahashi, K.; Herran, D.S.; Dai, H.; Hijioka, Y.; Kainuma, M. SSP3: AIM Implementation of Shared Socioeconomic Pathways. Global Environ. Chang. 2017, 42, 268–283. [Google Scholar] [CrossRef]
- Vittor, A.Y.; Laporta, G.Z.; Sallum, M.A.M.; Walker, R.T. The COVID-19 Crisis and Amazonia’s Indigenous People: Implications for Conservation and Global Health. World Dev. 2021, 145, 105533. [Google Scholar] [CrossRef]
- Wieczorek, J.; Guo, Q.; Hijmans, R. The Point-Radius Method for Georeferencing Locality Descriptions and Calculating Associated Uncertainty. Int. J. Geogr. Inf. Sci. 2004, 18, 745–767. [Google Scholar] [CrossRef]
- Chapman, A.; Wieczorek, J. Georeferencing Best Practices. GBIG 2020. [Google Scholar] [CrossRef]
- Laporta, G.Z.; Linton, Y.-M.; Wilkerson, R.C.; Bergo, E.S.; Nagaki, S.S.; Sant’Ana, D.C.; Sallum, M.A.M. Malaria Vectors in South America: Current and Future Scenarios. Parasit. Vectors 2015, 8, 426. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample Selection Bias and Presence-Only Distribution Models: Implications for Background and Pseudo-Absence Data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Hastie, T.; Tibshirani, R.; Friedman, J.H. The Elements of Statistical Learning: Data Mining, Inference, and Prediction, 2nd ed.; Springer Series in Statistics; Springer: New York, NY, USA, 2009; ISBN 978-0-387-84857-0. [Google Scholar]
- Santos, J.; Meneses, B.M. An Integrated Approach for the Assessment of the Aedes aegypti and Aedes albopictus Global Spatial Distribution, and Determination of the Zones Susceptible to the Development of Zika Virus. Acta Trop. 2017, 168, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Liu-Helmersson, J.; Brännström, Å.; Sewe, M.O.; Semenza, J.C.; Rocklöv, J. Estimating Past, Present, and Future Trends in the Global Distribution and Abundance of the Arbovirus Vector Aedes aegypti Under Climate Change Scenarios. Front. Public Health 2019, 7, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, P.M.; Betts, R.A.; Collins, M.; Harris, P.P.; Huntingford, C.; Jones, C.D. Amazonian Forest Dieback under Climate-Carbon Cycle Projections for the 21st Century. Theor. Appl. Climatol. 2004, 78, 137–156. [Google Scholar] [CrossRef]
- Malhi, Y.; Roberts, J.T.; Betts, R.A.; Killeen, T.J.; Li, W.; Nobre, C.A. Climate Change, Deforestation, and the Fate of the Amazon. Science 2008, 319, 169–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boisier, J.P.; Ciais, P.; Ducharne, A.; Guimberteau, M. Projected Strengthening of Amazonian Dry Season by Constrained Climate Model Simulations. Nat. Clim Chang. 2015, 5, 656–660. [Google Scholar] [CrossRef]
- Foster, P.G.; de Oliveira, T.M.P.; Bergo, E.S.; Conn, J.E.; Sant’Ana, D.C.; Nagaki, S.S.; Nihei, S.; Lamas, C.E.; González, C.; Moreira, C.C.; et al. Phylogeny of Anophelinae Using Mitochondrial Protein Coding Genes. R. Soc. Open Sci. 2017, 4, 170758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seixas, G.; Salgueiro, P.; Bronzato-Badial, A.; Gonçalves, Y.; Reyes-Lugo, M.; Gordicho, V.; Ribolla, P.; Viveiros, B.; Silva, A.C.; Pinto, J.; et al. Origin and Expansion of the Mosquito Aedes aegypti in Madeira Island (Portugal). Sci. Rep. 2019, 9, 2241. [Google Scholar] [CrossRef] [Green Version]
- Alves, M.J.; Fernandes, P.L.; Amaro, F.; Osório, H.; Luz, T.; Parreira, P.; Andrade, G.; Zé-Zé, L.; Zeller, H. Clinical Presentation and Laboratory Findings for the First Autochthonous Cases of Dengue Fever in Madeira Island, Portugal, October 2012. Eur. Surveill 2013, 18, 20398. [Google Scholar] [CrossRef]
- Cochet, A.; Calba, C.; Jourdain, F.; Grard, G.; Durand, G.A.; Guinard, A.; Investigation team; Noël, H.; Paty, M.-C.; Franke, F. Autochthonous Dengue in Mainland France, 2022: Geographical Extension and Incidence Increase. Eur. Surveill 2022, 27, 2200818. [Google Scholar] [CrossRef]
- Robino, D.M. Global Destination Cities Index 2019. MasterCard. Available online: https://www.mastercard.com/news/media/wexffu4b/gdci-global-report-final-1.pdf (accessed on 16 December 2022).
- Shanks, G.D. Reintroduced Malaria in Queensland, Australia during the Second World War. Intern. Med. J. 2021, 51, 1348–1351. [Google Scholar] [CrossRef] [PubMed]
- Grillet, M.E.; Hernández-Villena, J.V.; Llewellyn, M.S.; Paniz-Mondolfi, A.E.; Tami, A.; Vincenti-Gonzalez, M.F.; Marquez, M.; Mogollon-Mendoza, A.C.; Hernandez-Pereira, C.E.; Plaza-Morr, J.D.; et al. Venezuela’s Humanitarian Crisis, Resurgence of Vector-Borne Diseases, and Implications for Spillover in the Region. Lancet Infect. Dis. 2019, 19, e149–e161. [Google Scholar] [CrossRef] [Green Version]
- Hotez, P.J.; Basáñez, M.-G.; Acosta-Serrano, A.; Grillet, M.E. Venezuela and Its Rising Vector-Borne Neglected Diseases. PLoS Negl. Trop. Dis. 2017, 11, e0005423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soghigian, J.; Gloria-Soria, A.; Robert, V.; Le Goff, G.; Failloux, A.-B.; Powell, J.R. Genetic Evidence for the Origin of Aedes aegypti, the Yellow Fever Mosquito, in the Southwestern Indian Ocean. Mol. Ecol. 2020, 29, 3593–3606. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Song, Z.; Luo, L.; Wang, Q.; Zhou, G.; Yang, D.; Zhong, D.; Zheng, X. Molecular Evidence for New Sympatric Cryptic Species of Aedes albopictus (Diptera: Culicidae) in China: A New Threat from Aedes albopictus Subgroup? Parasit. Vectors 2018, 11, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greni, S.E.; Demari-Silva, B.; de Oliveira, T.M.P.; Suesdek, L.; Laporta, G.Z.; Sallum, M.A.M. A Multi-Gene Analysis and Potential Spatial Distribution of Species of the Strodei Subgroup of the Genus Nyssorhynchus (Diptera: Culicidae). J. Med. Entomol. 2018, 55, 1486–1495. [Google Scholar] [CrossRef] [PubMed]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-Based Registry for All Animal Species: The Barcode Index Number (BIN) System. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laporta, G.Z.; Potter, A.M.; Oliveira, J.F.A.; Bourke, B.P.; Pecor, D.B.; Linton, Y.-M. Global Distribution of Aedes aegypti and Aedes albopictus in a Climate Change Scenario of Regional Rivalry. Insects 2023, 14, 49. https://doi.org/10.3390/insects14010049
Laporta GZ, Potter AM, Oliveira JFA, Bourke BP, Pecor DB, Linton Y-M. Global Distribution of Aedes aegypti and Aedes albopictus in a Climate Change Scenario of Regional Rivalry. Insects. 2023; 14(1):49. https://doi.org/10.3390/insects14010049
Chicago/Turabian StyleLaporta, Gabriel Z., Alexander M. Potter, Janeide F. A. Oliveira, Brian P. Bourke, David B. Pecor, and Yvonne-Marie Linton. 2023. "Global Distribution of Aedes aegypti and Aedes albopictus in a Climate Change Scenario of Regional Rivalry" Insects 14, no. 1: 49. https://doi.org/10.3390/insects14010049