

Future Trends in Obolodiplosis robiniae Distribution across Eurasian Continent under Global Climate Change


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. The Species Occurrence Data and Environmental Variables
2.2. Optimization of the Model Parameter
2.3. Classification of CPD and FPD
2.4. Assessing the Accuracy of the Model
3. Results
3.1. The Main Parameters and Performance of the Model
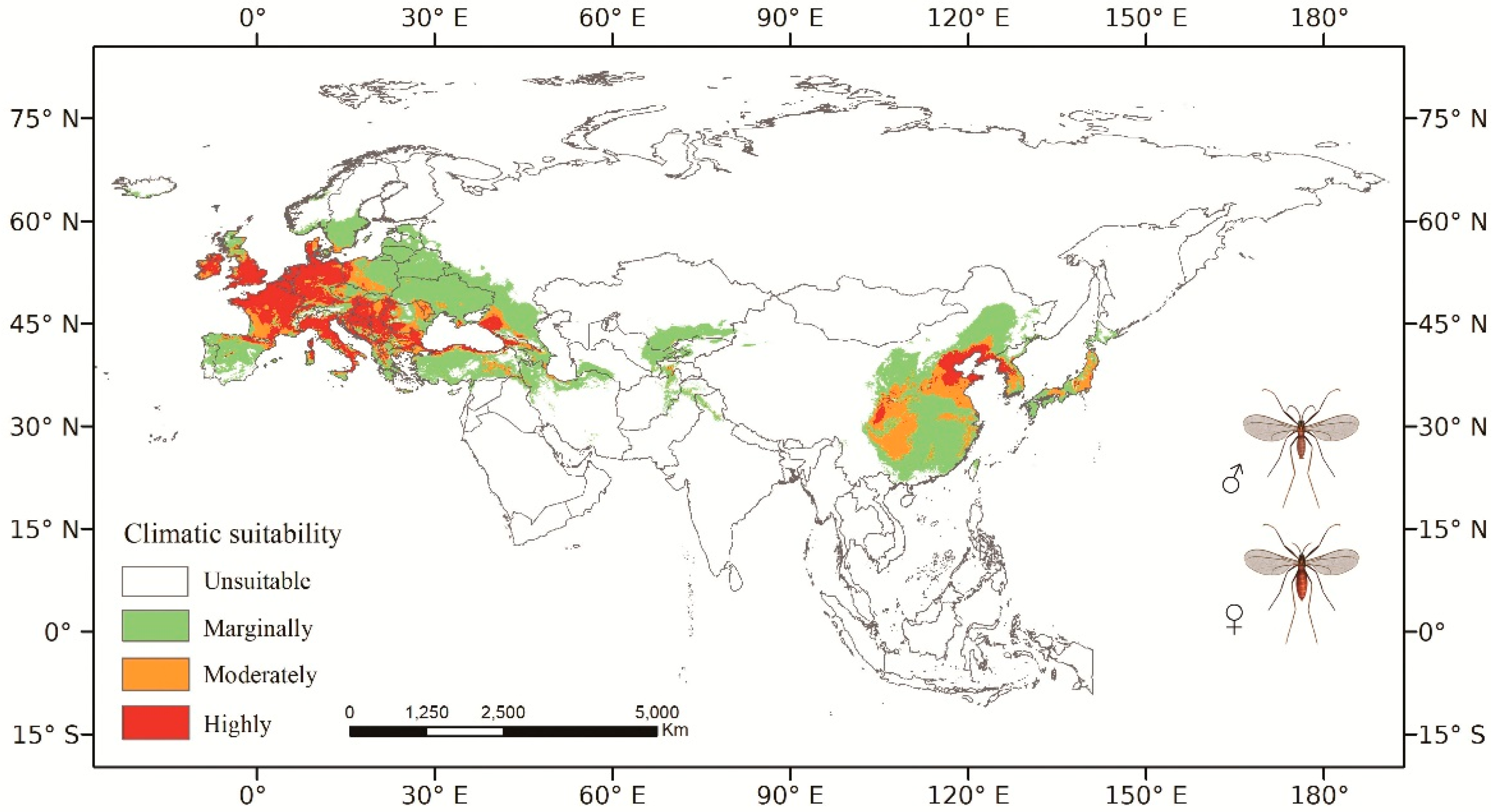
3.2. Predicted Current Potential Distribution
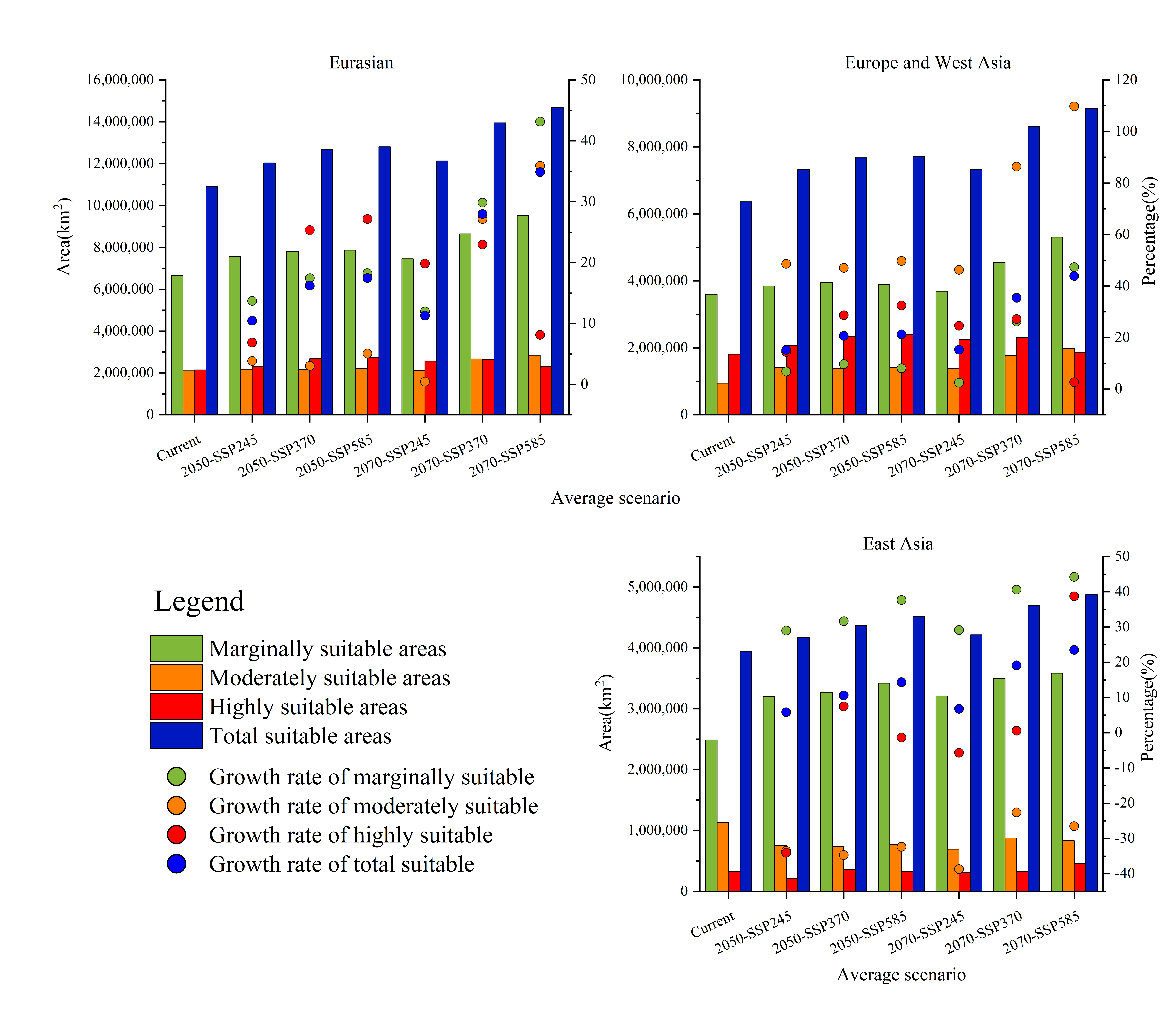
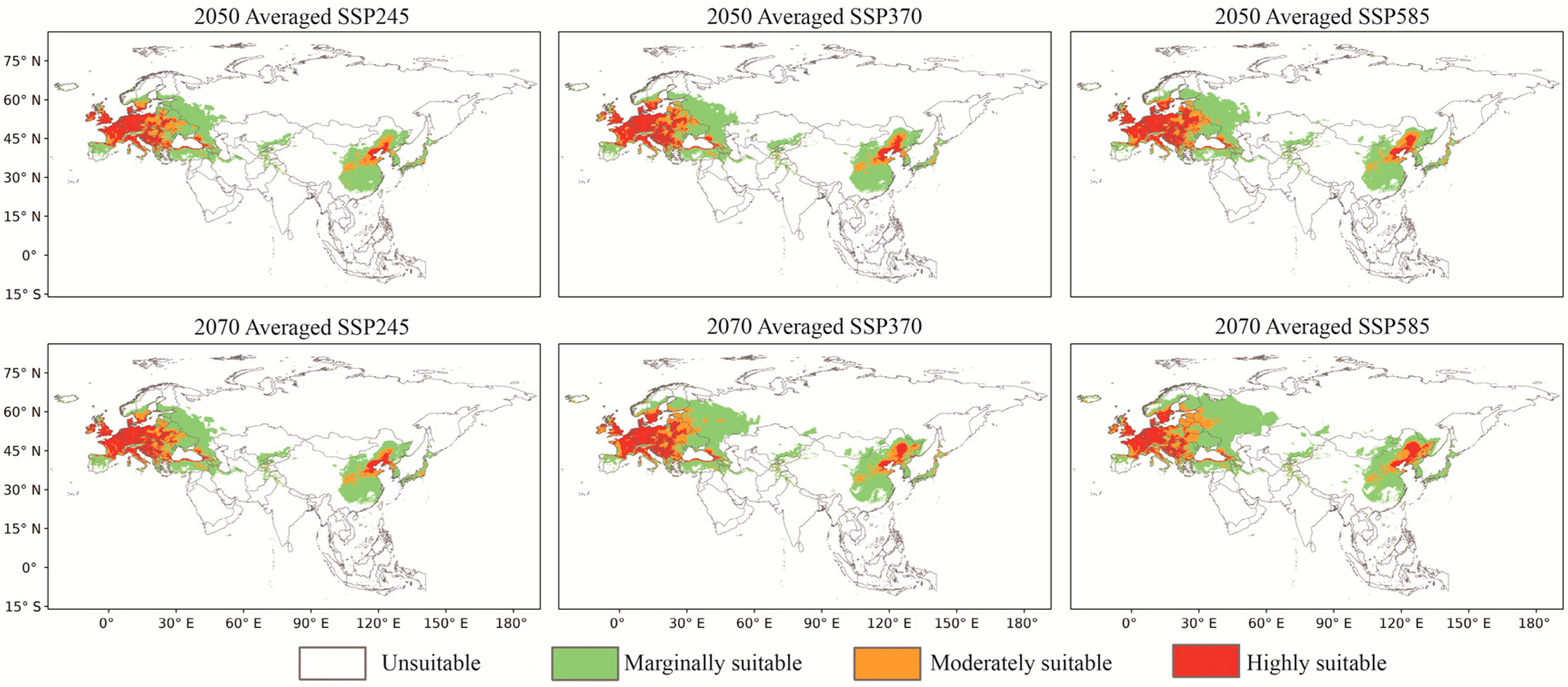
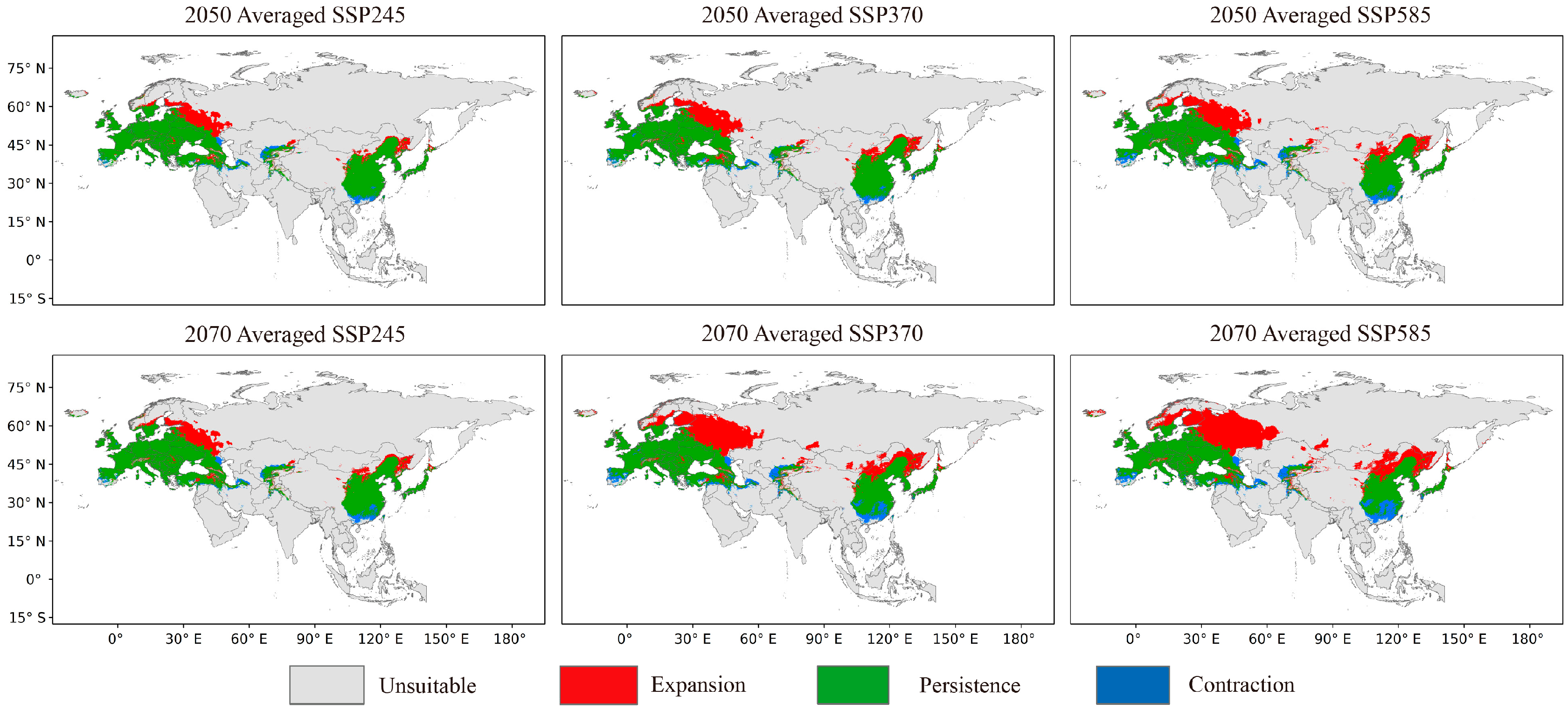
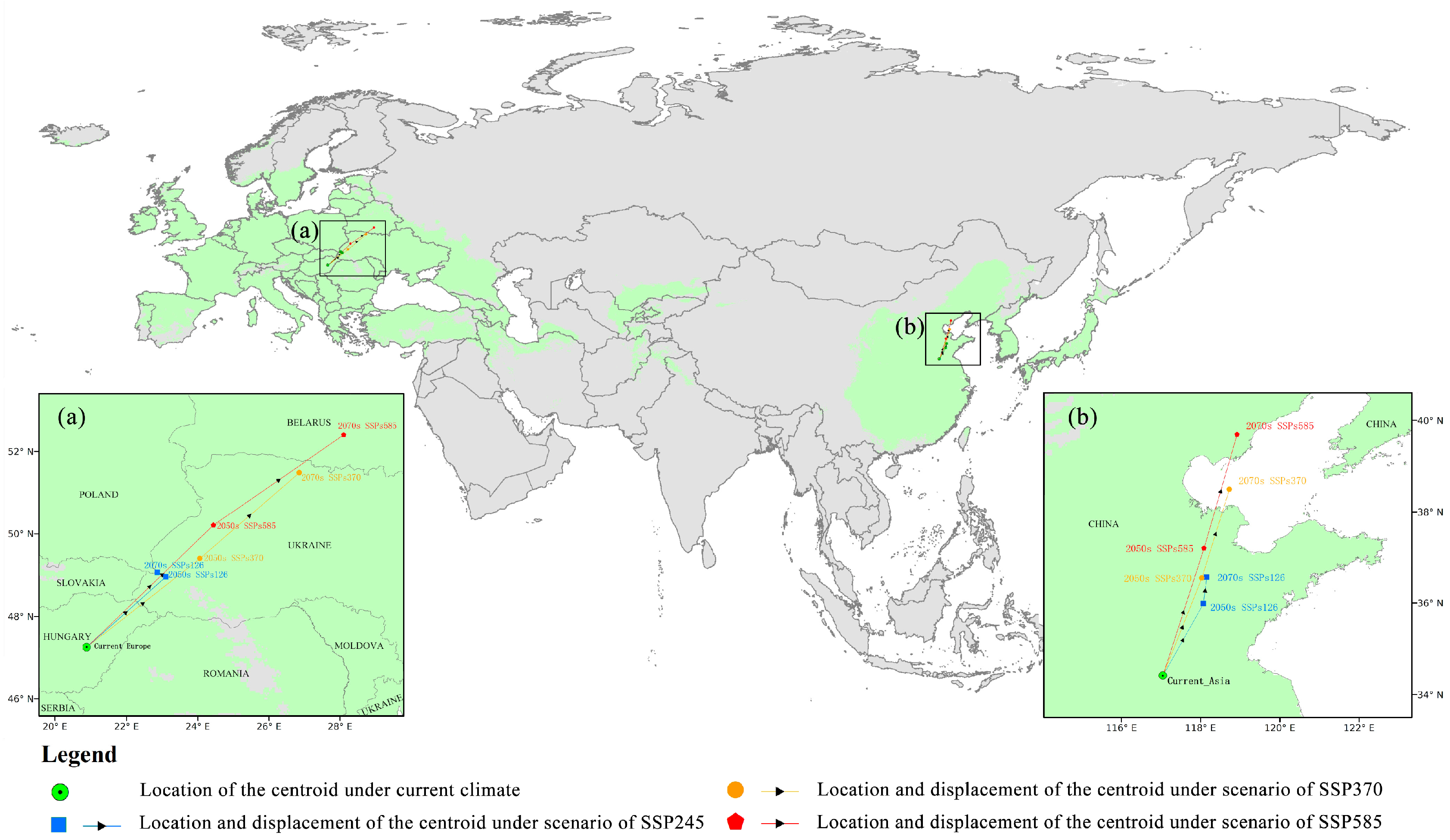
3.3. Predicted Future Potential Distribution and Analyzed Tendencies
4. Discussion
4.1. Analysis of Model Results and CPD
4.2. Analysis of the Trends of the FPD of O. robiniae
4.3. Final Considerations and Quarantine Management Measures
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seebens, H.; Bacher, S.; Blackburn, T.M.; Capinha, C.; Dawson, W.; Dullinger, S.; Genovesi, P.; Hulme, P.E.; van Kleunen, M.; Kühn, I. Projecting the continental accumulation of alien species through to 2050. Glob. Chang. Biol. 2021, 27, 968–969. [Google Scholar] [CrossRef]
- Essl, F.; Lenzner, B.; Bacher, S.; Bailey, S.; Capinha, C.; Daehler, C.; Dullinger, S.; Genovesi, P.; Hui, C.; Hulme, P.E. Drivers of future alien species impacts: An expert-based assessment. Glob. Chang. Biol. 2020, 26, 4880–4893. [Google Scholar] [CrossRef]
- Mack, R.; Smith, M. Invasive plants as catalysts for the spread of human parasites. NeoBiota 2011, 9, 13–29. [Google Scholar] [CrossRef] [Green Version]
- Vanbergen, A.J.; Espíndola, A.; Aizen, M.A. Risks to pollinators and pollination from invasive alien species. Nat. Ecol. Evol. 2018, 2, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Diagne, C.; Leroy, B.; Vaissière, A.-C.; Gozlan, R.E.; Roiz, D.; Jarić, I.; Salles, J.-M.; Bradshaw, C.J.A.; Courchamp, F. High and rising economic costs of biological invasions worldwide. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef]
- Vilà, M.; Corbin, J.D.; Dukes, J.S.; Pino, J.; Smith, S.D. Linking plant invasions to global environmental change. In Terrestrial Ecosystems in a Changing World; Springer: Berlin/Heidelberg, Germany, 2007; pp. 93–102. [Google Scholar]
- Descombes, P.; Pitteloud, C.; Glauser, G.; Defossez, E.; Kergunteuil, A.; Allard, P.-M.; Rasmann, S.; Pellissier, L. Novel trophic interactions under climate change promote alpine plant coexistence. Science 2020, 370, 1469–1473. [Google Scholar] [CrossRef]
- McCain, C.M.; Garfinkel, C.F. Climate change and elevational range shifts in insects. Curr. Opin. Insect Sci. 2021, 47, 111–118. [Google Scholar] [CrossRef]
- Kenis, M.; Branco, M. Impact of alien terrestrial arthropods in Europe. Chapter 5. BioRisk 2010, 4, 51–71. [Google Scholar] [CrossRef] [Green Version]
- Raza, M.M.; Khan, M.A.; Arshad, M.; Sagheer, M.; Sattar, Z.; Shafi, J.; Haq, E.u.; Ali, A.; Aslam, U.; Mushtaq, A. Impact of global warming on insects. Arch. Phytopathol. Plant Prot. 2015, 48, 84–94. [Google Scholar] [CrossRef]
- Tóth, P.; Váňová, M.; Lukáš, J. The distribution of Obolodiplosis robiniae on black locust in Slovakia. J. Pest Sci. 2009, 82, 61–66. [Google Scholar] [CrossRef]
- Zang, Z. Technical points for the control of Obolodiplosis robiniae. China Agric. Inf. 2015, 10, 69–70. [Google Scholar]
- Shao, X.; Ma, X.; Shao, K.; Lu, J.; Kao, G. Occurrence, endangered grade and prevention and control experiments of Oblodiplosis robiniae. Liaoning For. Sci. Technol. 2010, 31–32, 45–49. [Google Scholar] [CrossRef]
- Wermelinger, B.; Skuhrava, M. First records of the gall midge Obolodiplosis robiniae (Haldeman) (Diptera: Cecidomyiidae) and its associated parasitoid Platygaster robiniae Buhl & Duso (Hymenoptera: Platygastridae) in Switzerland. Bull. Soc. Entomol. Suisse. 2007, 80, 217–221. [Google Scholar]
- Yao, Y.; Zhao, W.; Shang, X. Development of polymorphic microsatellite markers of Obolodiplosis robiniae (Haldeman)(Diptera: Cecidomyiidae), a North American pest invading Asia. J. Insect Sci. 2015, 15, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Glavendekić, M.; Roques, A.; Mihajlović, L. An ALARM case study: The rapid colonization of an introduced tree, black locust by an invasive North-American midge and its parasitoids. In Atlas of Biodiversity Risk; Settele, J., Penev, L.D., Georgiev, T.A., Grabaum, R., Grobelnik, V., Hammen, V., Klotz, S., Kotarac, M.I.K., Eds.; Pensoft: Sofia, Bulgaria, 2009; pp. 22–23. [Google Scholar]
- Haldeman, S.S. Description of several new and interesting animals. Am. J. Agric. Sci. 1847, 6, 191–194. [Google Scholar]
- Uechi, N.; Yukawa, J.; Usuba, S. Recent distributional records of an alien gall midge, Obolodiplosis robiniae (Diptera: Cecidomyiidae) in Japan, and a brief description of its pupal morphology. Kyushu Plant Prot. Res. 2005, 51, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Kodoi, F.; Lee, H.-S.; Uechi, N.; Yukawa, J. Occurrence of Obolodiplosis robiniae (Diptera: Cecidomyiidae) in Japan and South Korea. Esakia 2003, 43, 35–41. [Google Scholar] [CrossRef]
- Woo, K.; Choe, H.; Kim, H. A report on the occurrence of yellow locust midge Obolodiplosis robiniae (Haldeman, 1987) from Korea. Korean J. Appl. Entomol. 2003, 42, 77–79. [Google Scholar] [CrossRef]
- Duso, C.; Fontana, P.; Tirello, P. Spread of the gall midge Obolodiplosis robiniae (Haldeman) injurious to black locust in Italy and Europe. Inf. Fitopatol. 2005, 55, 30–33. [Google Scholar]
- Bálint, J.; Neacşu, P.; Balog, A.; Fail, J.; Vétek, G. First record of the black locust gall midge Obolodiplosis robiniae (Haldeman)(Diptera: Cecidomyiidae) in Romania. North-West. J. Zool. 2010, 6, 319–322. [Google Scholar] [CrossRef]
- Berest, Z.; Titar, V. Robinieva krajova galicja (Obolodiplosis robiniae)(Diptera: Cecidomyiidae), mozlivist podalsogo rozsirennja arealu v Ukraini. Karantin I Zachist Rostl. 2007, 7, 24–26. [Google Scholar]
- Skuhravá, M.; Skuhravý, V.; Csóka, G. The invasive spread of the gall midge Obolodiplosis robiniae in Europe. Cecidology 2007, 22, 84–90. [Google Scholar]
- Molnar, B.; BodduM, T.; Szőcs, G.; HillBur, Y. Occurrence of two pest gall midges, Obolodiplosis robiniae (Haldeman) and Dasineura gleditchiae (Osten Sacken)(Diptera: Cecidomyiidae) on ornamental trees in Sweden. Entomol. Tidskr. 2009, 130, 113–120. [Google Scholar]
- Yang, Z.; Qiao, X.; Bu, W.; Yao, Y.; Xiao, Y.; Han, Y. First discovery of an important invasive insect pest, Obolodiplosis robiniae (Diptera: Cecidomyiidae) in China. Acta Entomol. Sin. 2006, 49, 1050–1053. [Google Scholar] [CrossRef]
- Wang, R.; Wang, L.; Wang, S.; Li, B.; Wang, Z. A New Pest of Robinia -Preliminary observations on Obolodiplosis robiniae. J. Liaoning Agric. Coll. 2006, 8, 24–25. [Google Scholar]
- Zhang, W.; Fu, J.; Wei, G.; Sun, Y. Obolodiplosis robiniae was found in Jilin City. North Hortic. 2008, 6, 207. [Google Scholar]
- Mu, X.; Sun, J.; Lu, W.; Li, M.; Qu, H.; Gao, Z. Bionomics and control of Obolodiplosis robiniae in Beijing. For. Pest Dis. 2010, 29, 15–18. [Google Scholar] [CrossRef]
- Yan, J.; Wang, S.; Li, D. A new foreign forest pest found in Shandong-Obolodiplosis robiniae. Shandong For. Sci. Technol. 2007, 6, 60. [Google Scholar]
- Zhang, D.; Lu, C.; Wang, X.; Gao, B. Potential Risk Assessment of Obolodiplosis Robininae (Haldemann) in China. Acta Ecol. Sin. 2009, 29, 2155–2161. [Google Scholar] [CrossRef]
- Zhao, J.; Shi, J. Prediction of the potential geographical distribution of Obolodiplosis robiniae (Diptera: Cecidomyiidae) in China based on a novel maximum entropy model. Sci. Silvae Sin. 2019, 55, 118–127. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. A J. R. Meteorol. Soc. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Tingley, R.; Vallinoto, M.; Sequeira, F.; Kearney, M.R. Realized niche shift during a global biological invasion. Proc. Natl. Acad. Sci. USA 2014, 111, 10233–10238. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Illan, J.G.; Looney, C.; Crowder, D.W. Assessing the ecological niche and invasion potential of the Asian giant hornet. Proc. Natl. Acad. Sci. USA 2020, 117, 24646–24648. [Google Scholar] [CrossRef]
- Tóth, P.; Váňová, M.; Lukáš, J. Impact of natural enemies on Obolodiplosis robiniae invasion. Biologia 2011, 66, 870–876. [Google Scholar] [CrossRef]
- Bakay, L.; Kollár, J. Insect pests on the black locust (Robinia pseudoacacia L.) in the locality Čebovce. Acta Hortic. Regiotect. 2010, 13, 55–57. [Google Scholar]
- Han, L.; Wu, J.; Wang, D. Occurrence of Obolodiplosis robiniae Haldemann and Corresponding Measures to control. Shaanxi For. Sci. Technol. 2010, 3, 41–43. [Google Scholar] [CrossRef]
- Petrov, D.L. The teratoforming phytophagous arthropods damaging ornamental woody plants, expanded on the territory of belarus in the current century. J. Belarus. State Univ. Ecol. 2019, 1, 24–31. Available online: https://217.21.43.57/~ojs/ojs.bsu.by/index.php/ecology/article/view/2694 (accessed on 1 March 2021). (In Russian).
- Peterson, A.; Papeş, M.; Eaton, M. Transferability and model evaluation in ecological niche modeling: A comparison of GARP and Maxent. Ecography 2007, 30, 550–560. [Google Scholar] [CrossRef]
- Qin, Y.; Zhang, Y.; Clarke, A.R.; Zhao, Z.; Li, Z. Including Host Availability and Climate Change Impacts on the Global Risk Area of Carpomya pardalina (Diptera: Tephritidae). Front. Ecol. Evol. 2021, 9, 724441. [Google Scholar] [CrossRef]
- Teixeira, C.M.; Krüger, A.P.; Nava, D.E.; Garcia, F.R.M. Potential global distribution of the south American cucurbit fruit fly Anastrepha grandis (Diptera: Tephritidae). Crop Prot. 2021, 145, 105647. [Google Scholar] [CrossRef]
- Wu, S.; Liu, L.; Gao, J.; Wang, W. Integrate Risk From Climate Change in China Under Global Warming of 1.5 and 2.0 °C. Earth’s Future 2019, 7, 1307–1322. [Google Scholar] [CrossRef]
- Fan, X.; Miao, C.; Duan, Q.; Shen, C.; Wu, Y. Future Climate Change Hotspots Under Different 21st Century Warming Scenarios. Earth’s Future 2021, 9, e2021EF002027. [Google Scholar] [CrossRef]
- Bakay, L.; Kollár, J. Influence of urban environment on the population biology of alien insect species: Obolodiplosis robiniae (Haldeman, 1847), Parectopa robiniella Clemens, 1863 and Phyllonorycter robiniella Clemens, 1859 in conditions of Slovakia. Pensee 2014, 76, 88–94. [Google Scholar]
- Puchałka, R.; Dyderski, M.K.; Vítková, M.; Sádlo, J.; Klisz, M.; Netsvetov, M.; Prokopuk, Y.; Matisons, R.; Mionskowski, M.; Wojda, T. Black locust (Robinia pseudoacacia L.) range contraction and expansion in Europe under changing climate. Glob. Chang. Biol. 2021, 27, 1587–1600. [Google Scholar] [CrossRef]
- The Intergovernmental Panel on Climate Change. The IPCC and the Sixth Assessment Cycle. 2020. Available online: https://www.ipcc.ch/assessment-report/ar6/. (accessed on 1 March 2021).
- Klisz, M.; Puchałka, R.; Netsvetov, M.; Prokopuk, Y.; Vítková, M.; Sádlo, J.; Matisons, R.; Mionskowski, M.; Chakraborty, D.; Olszewski, P. Variability in climate-growth reaction of Robinia pseudoacacia in Eastern Europe indicates potential for acclimatisation to future climate. For. Ecol. Manag. 2021, 492, 119194. [Google Scholar] [CrossRef]
- Gilbert, M.; Guichard, S.; Freise, J.; Grégoire, J.C.; Heitland, W.; Straw, N.; Tilbury, C.; Augustin, S. Forecasting Cameraria ohridella invasion dynamics in recently invaded countries: From validation to prediction. J. Appl. Ecol. 2005, 42, 805–813. [Google Scholar] [CrossRef]
- Pernek, M.; Matošević, D. Black Locust Gall Midge (Obolodiplosis robiniae), New Pest on Black Locust Trees and first Record of ParasitoidPlatygaster robinaein Croatia. Šumarski List 2009, 133, 157–163. [Google Scholar]
- Duso, C.; Boaria, A.; Surian, L.; Buhl, P. Seasonal abundance of the nearctic gall midge Obolodiplosis robiniae in Italy and the impact of its antagonist Platygaster robiniae on pest populations. Ann. Entomol. Soc. Am. 2011, 104, 180–191. [Google Scholar] [CrossRef]
- Yu, J.; Lu, C.; Wen, X.; Wang, J.; Zhang, L.; Song, G.; Zhao, Z. The Functional Response of Chrysopa sinica Tjeder to Obolodiplosis robiniae (Haldemann) Larvae. J. Hebei Norml. Univ. Sci. Technol. 2009, 23, 21–24. [Google Scholar] [CrossRef]
- Yao, Y.; Yang, Z.; Yan, J. A new species of the genus Systasis Walker, 1834 (Hymenoptera: Pteromalidae) parasitizing an important invasive alien insect pest Obolodiplosis robiniae (Diptera: Cecidomyiidae) from China. Sci. Silvae Sin. 2009, 45, 88–90. [Google Scholar] [CrossRef]
- Bella, S. Presenza di Obolodiplosis robiniae (Haldeman, 1847) in Italia centro-meridionale e in Sicilia (Diptera Cecidomyiidae). Boll. Di Zool. Agrar. E Di Bachic. 2007, 39, 239–242. [Google Scholar]
- Lu, C.; Buhl, P.N.; Duso, C.; Zhao, C.; Zhang, J.; Ji, Z.; Gao, S.; Yu, J.; Wen, X. First discovery of Platygaster robiniae(Hymenoptera: Platygastridae) parasitizing the invasive Obolodiplosis robiniae Diptera:Cecidomyiidae), a gallmaker in China. Acta Entomol. Sin. 2010, 53, 233–237. [Google Scholar] [CrossRef]
- Buhl, P.N.; Duso, C. Platygaster robiniae n. sp.(Hymenoptera: Platygastridae) parasitoid of Obolodiplosis robiniae (Diptera: Cecidomyiidae) in Europe. Ann. Entomol. Soc. Am. 2008, 101, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.-K.; Park, J.-D.; Shin, S.-C.; Park, I.-K. Prolonged embryonic stage and synchronized life-history of Platygaster robiniae (Hymenoptera: Platygastridae), a parasitoid of Obolodiplosis robiniae (Diptera: Cecidomyiidae). Biol. Control 2011, 57, 24–30. [Google Scholar] [CrossRef]
- Park, J.-D.; Shin, S.-C.; Kim, C.-S.; Jeon, M.-J.; Park, I.-K. Biological characteristic of Obolodiplosis robiniae and control effects of some insecticides. Korean J. Appl. Entomol. 2009, 48, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Fitter, A.; Fitter, R. Rapid changes in flowering time in British plants. Science 2002, 296, 1689–1691. [Google Scholar] [CrossRef]
- Ma, Q.; Huang, J.G.; Hänninen, H.; Li, X.; Berninger, F. Climate warming prolongs the time interval between leaf-out and flowering in temperate trees: Effects of chilling, forcing and photoperiod. J. Ecol. 2021, 109, 1319–1330. [Google Scholar] [CrossRef]
- Bella, S. Invasive insect pests and their associated parasitoids on ornamental urban plants on Corfu island-Steyskal and Spencer 1978 (Diptera, Agromyzidae) a new record in Greece. Hell. Plant Prot. J. 2014, 7, 53–59. [Google Scholar] [CrossRef]
- Zhao, C.; Gao, S.; Xue, H.; Song, G.; Zhao, Z. Biological Features of Obolodiplosis robiniae (Haldemann) Adults. J. Hebei Norml Univ. Sci. Technol. 2011, 25, 61–65. [Google Scholar] [CrossRef]
- Vaes-Petignat, S.; Nentwig, W. Environmental and economic impact of alien terrestrial arthropods in Europe. NeoBiota 2014, 22, 23. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Environmental Variables | Percent Contribution |
|---|---|---|
| Bio1 | Annual Mean Temperature | 59.2 |
| Bio12 | Annual Precipitation | 14.3 |
| Bio14 | Precipitation of Driest Month | 13.1 |
| Bio3 | Isothermality | 5.5 |
| Bio15 | Precipitation Seasonality | 4.9 |
| Bio6 | Min. Temperature of Coldest Month | 3.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.-Q.; Gao, T.; Du, J.-J.; Shi, J. Future Trends in Obolodiplosis robiniae Distribution across Eurasian Continent under Global Climate Change. Insects 2023, 14, 48. https://doi.org/10.3390/insects14010048
Zhao J-Q, Gao T, Du J-J, Shi J. Future Trends in Obolodiplosis robiniae Distribution across Eurasian Continent under Global Climate Change. Insects. 2023; 14(1):48. https://doi.org/10.3390/insects14010048
Chicago/Turabian StyleZhao, Jia-Qiang, Tai Gao, Jing-Jing Du, and Juan Shi. 2023. "Future Trends in Obolodiplosis robiniae Distribution across Eurasian Continent under Global Climate Change" Insects 14, no. 1: 48. https://doi.org/10.3390/insects14010048