
Insulin-like Peptides of the Western Flower Thrips Frankliniella occidentalis and Their Mediation of Immature Development


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Host Plant Culture
2.2. Bioinformatics to Predict ILP Genes in F. occidentalis
2.3. RNA Extraction and cDNA Preparation
2.4. RT-PCR and RT-qPCR
2.5. RNA Interference (RNAi) of ILP Genes Using Gene-Specific Double-Stranded RNA (dsRNA)
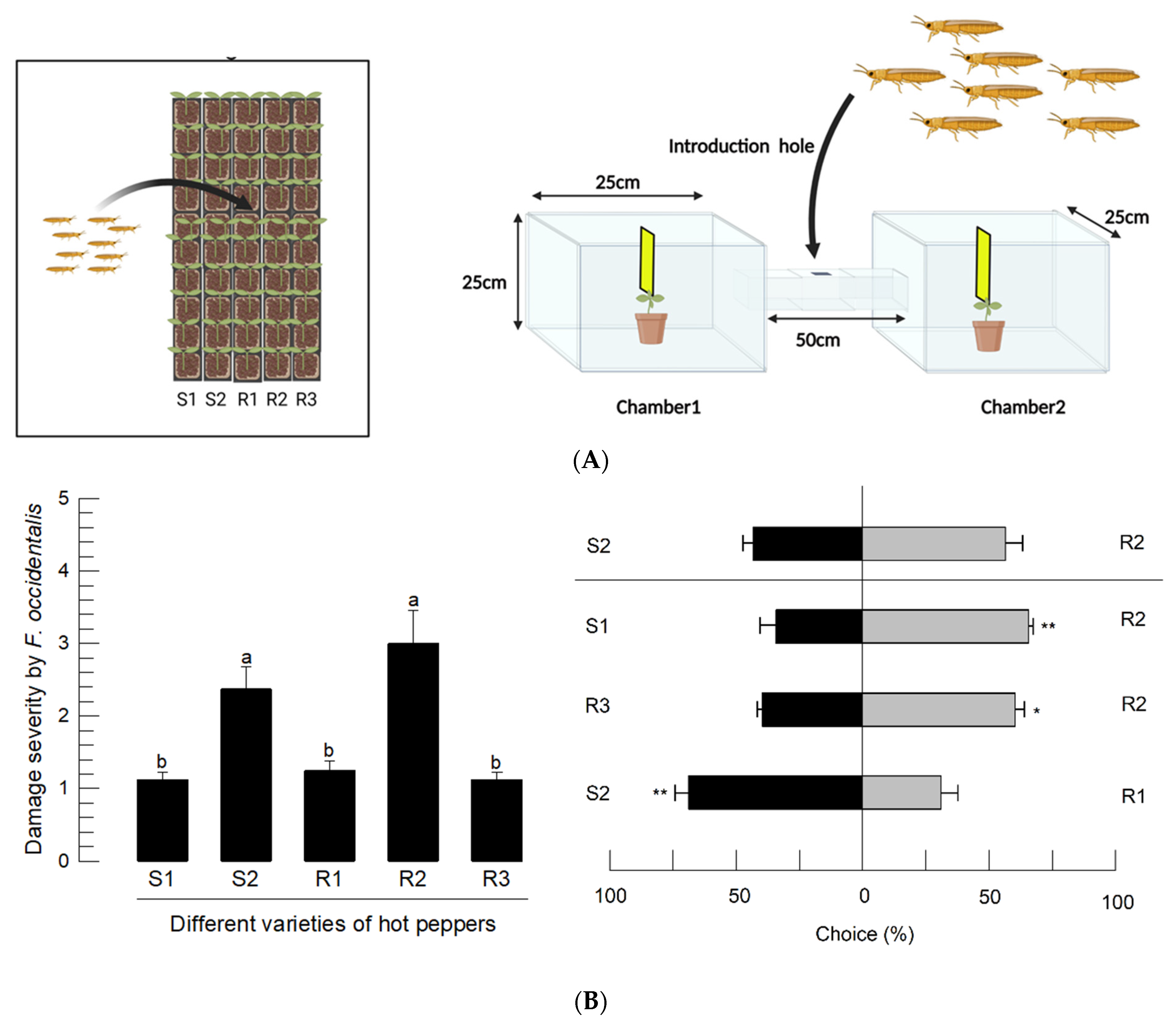
2.6. Screening Hot Pepper Varieties by Assessment of Feeding Damage
2.7. Choice Test Using a Two-Way Tunnel Assay
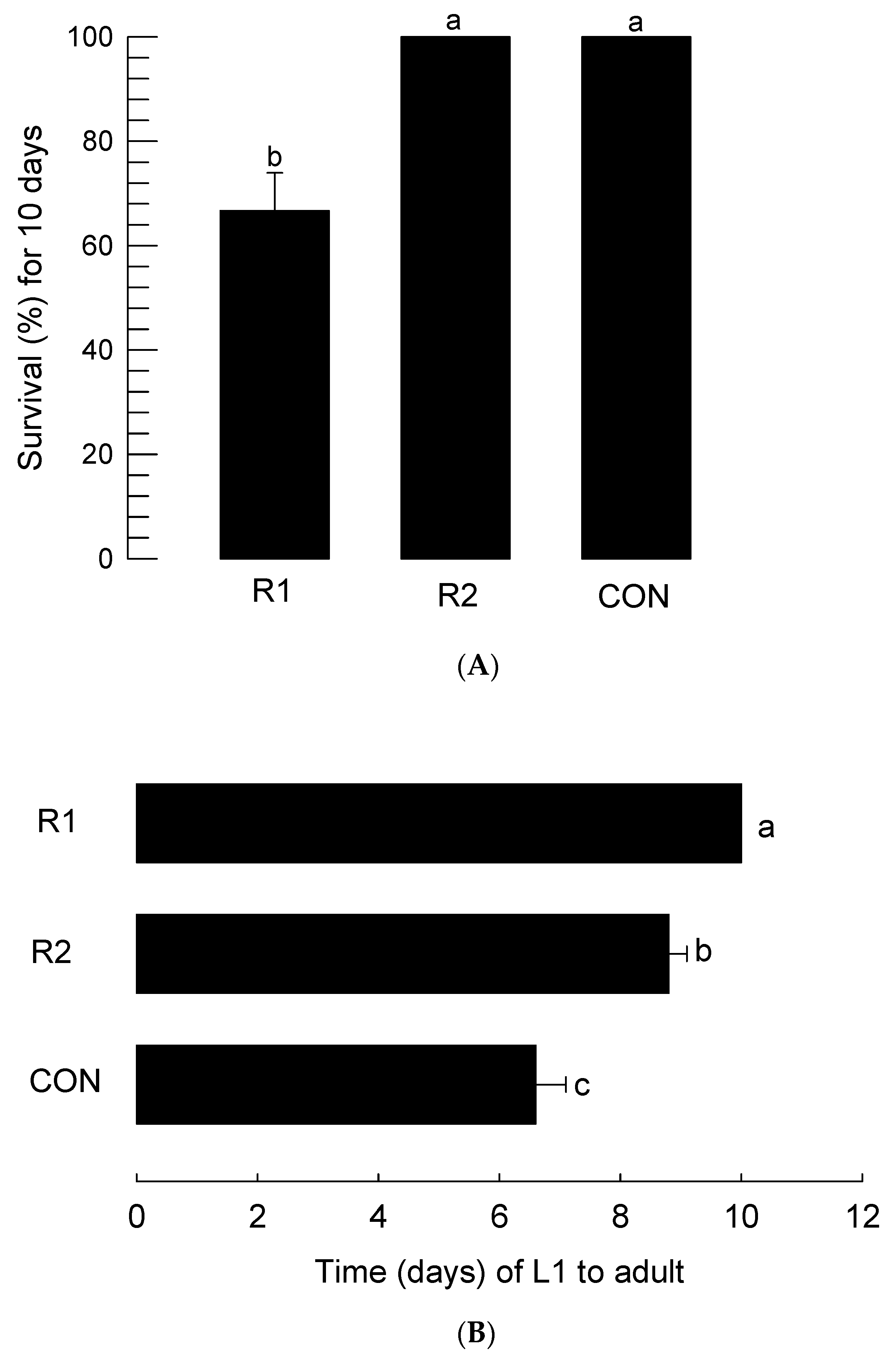
2.8. Assessment of Immature Development Period and Survival Rate for Two Hot Pepper Varieties
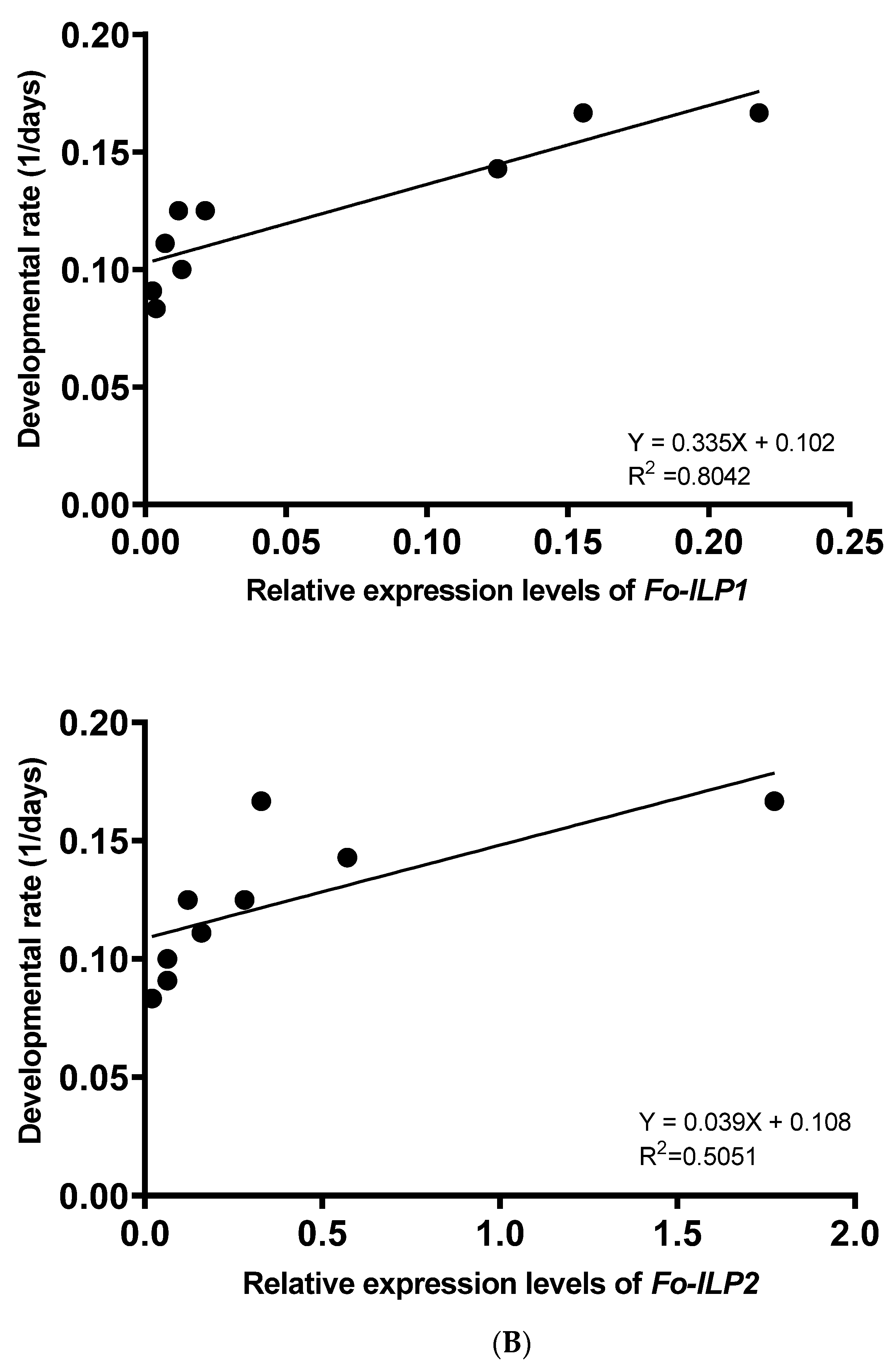
2.9. Correlation Analysis between Developmental Rate and ILP Expression
2.10. Statistical Analysis
3. Results
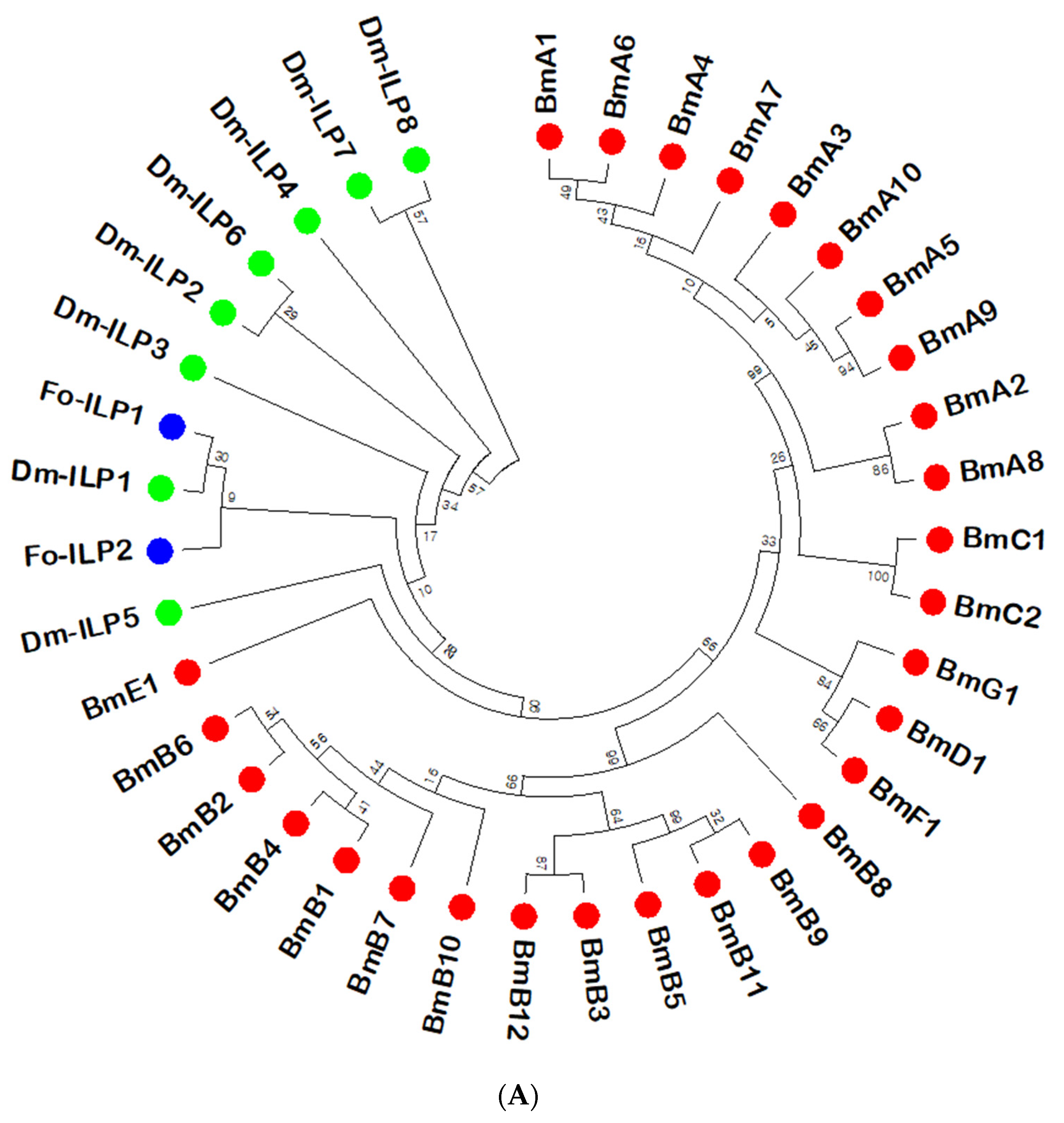
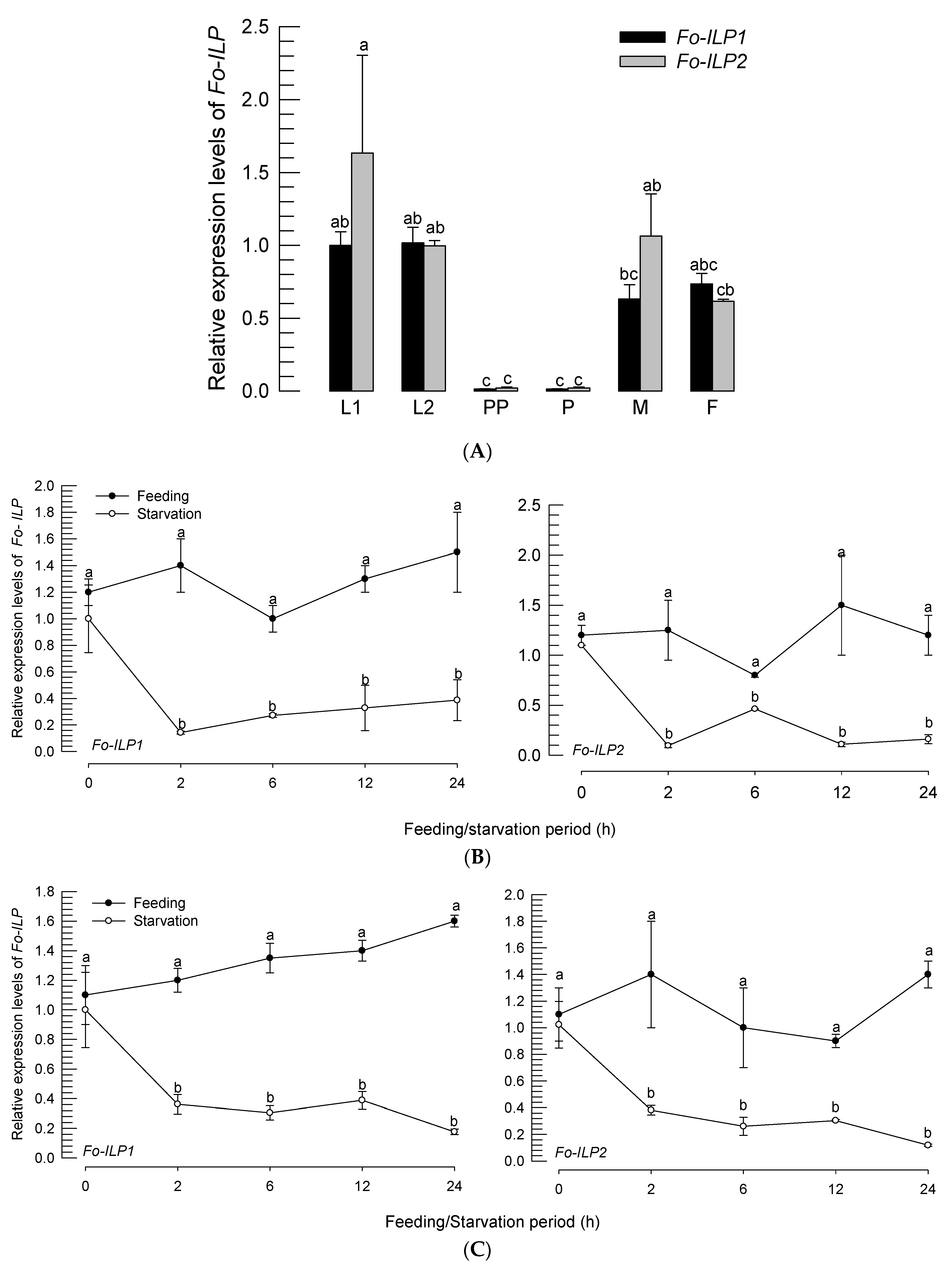
3.1. ILPs of F. occidentalis (Fo-ILPs) and Their Expression Profiles
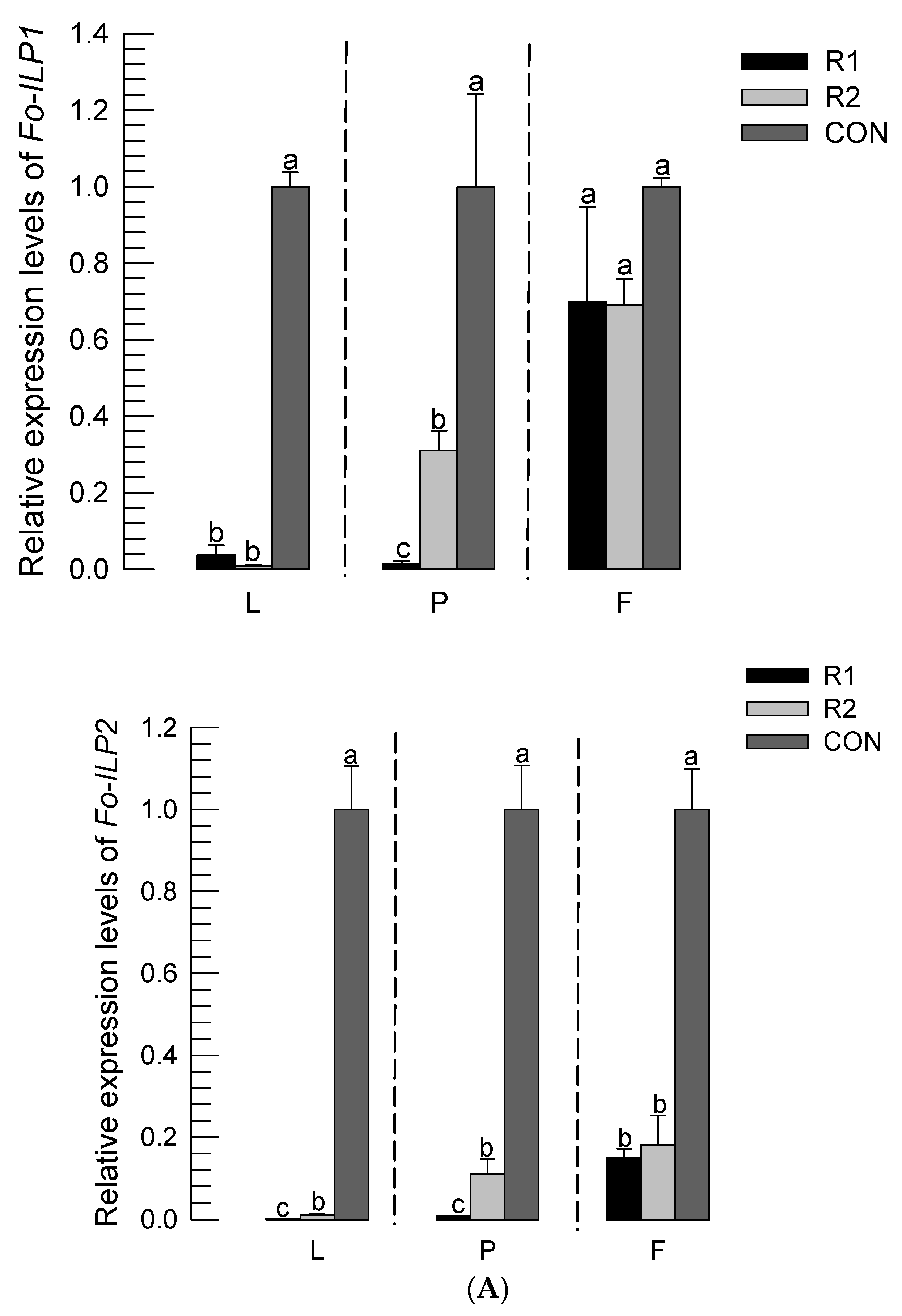
3.2. RNAi of ILP Gene Expression Led to Developmental Retardation
3.3. Developmental Variations of F. occidentalis for Different Varieties of Hot Pepper
3.4. Expression Levels of Fo-ILPs Are Correlated with Immature Development of F. occidentalis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Nagasawa, H.; Kataoka, H.; Isogai, A.; Tamura, S.; Suzuki, A.; Ishizaki, H. Amino-terminal amino acid sequence of the silkworm prothoracicotropic hormone: Homology with insulin. Science 1984, 226, 1344–1345. [Google Scholar] [CrossRef] [PubMed]
- Aslam, A.F.; Kiya, T.; Mita, K.; Iwami, M. Identification of novel bombyxin genes from the genome of the silkmoth Bombyx mori and analysis of their expression. Zool. Soc. 2011, 28, 609–616. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.; Nässel, D.R. Drosophila insulin-like peptide 8 (DILP8) in ovarian follicle cells regulates ovulation and metabolism. Front. Endocrinol. 2020, 11, 461. [Google Scholar] [CrossRef] [PubMed]
- Al Baki, M.A.; Jung, J.K.; Kim, Y. Physiological alterations in mutants of two insulin-like peptides encoded in Maruca vitrata using CRISPR/Cas9. Front. Physiol. 2021, 12, 701616. [Google Scholar] [CrossRef] [PubMed]
- Antonova, Y.; Arik, A.J.; Moore, W.; Riehle, M.A.; Brown, M.R. Insulin-like peptides: Structure, signaling, and function. In Insect Endocrinology; Gilbert, L.I., Ed.; Academic Press Deletion: New York, NY, USA, 2012; pp. 63–92. [Google Scholar]
- Mathieu, C.; Gillard, P.; Benhalima, K. Insulin analogues in type 1 diabetes mellitus: Getting better all the time. Nat. Rev. Endocrinol. 2017, 13, 385–399. [Google Scholar] [CrossRef]
- Géminard, C.; Rullifson, E.J.; Leopold, P. Remote control of insulin secretion by fat body cells in Drosophila. Cell Metab. 2009, 10, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, A.E.; Ullman, D.E.; German, T.L. Tospovirus-thrips interactions. Annu. Rev. Phytopathol. 2005, 43, 459–489. [Google Scholar] [CrossRef]
- Morse, J.G.; Hoddle, M.S. Invasion biology of thrips. Annu. Rev. Entomol. 2005, 1, 67–89. [Google Scholar] [CrossRef] [Green Version]
- Pearsall, I.A. Flower preference behaviour of western flower thrips in the Similkameen valley, British Columbia, Canada. Entomol. Exp. Appl. 2000, 95, 303–313. [Google Scholar] [CrossRef]
- Mainali, B.P.; Lim, U.T. Behavioral response of western flower thrips to visual and olfactory cues. J. Insect Behav. 2011, 24, 436–446. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, D.; Ahmed, S. First report of insulin receptor in Thysanoptera and its expression variation with development of western flower thrips, Frankliniella occidentalis. J. Asia Pac. Entomol. 2021, 24, 1004–1009. [Google Scholar] [CrossRef]
- Post, S.; Karashchuk, G.; Wade, J.D.; Sajid, W.; De Meyts, P.; Tatar, M. Drosophila insulin-like peptides DILP2 and DILP5 differentially stimulate cell signaling and glycogen phosphorylase to regulate longevity. Front. Endocrinol. 2018, 9, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 4. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Vatanparast, M.; Kim, Y. Optimization of recombinant bacteria expressing dsRNA to enhance insecticidal activity against a lepidopteran insect, Spodoptera exigua. PLoS ONE 2017, 8, e0183054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.; Roy, M.C.; Al Baki, M.A.; Jung, J.K.; Lee, D.; Kim, Y. CRISPR/Cas9 mutagenesis against sex pheromone biosynthesis leads to loss of female attractiveness in Spodoptera exigua, an insect pest. PLoS ONE 2021, 16, e0259322. [Google Scholar] [CrossRef]
- SAS Institute, Inc. SAS/STAT User’s Guide; SAS Institute, Inc.: Cary, NC, USA, 1989. [Google Scholar]
- Wu, Q.; Brown, M.R. Signaling and function of insulin-like peptides in insects. Annu. Rev. Entomol. 2006, 51, 1–24. [Google Scholar] [CrossRef]
- Rajan, A.; Perrimon, N. Drosophila cytokine unpaired 2 regulates physiological homeostasis by remotely controlling insulin secretion. Cell 2012, 151, 123–137. [Google Scholar] [CrossRef] [Green Version]
- Sano, H.; Nakamura, A.; Texada, M.J.; Truman, J.W.; Ishimoto, H.; Kamikouchi, A.; Nibu, Y.; Kume, K.; Ida, T.; Kojima, M. The nutrient-responsive hormone CCHamide-2 controls growth by regulating insulin-like peptides in the brain of Drosophila melanogaster. PLoS Genet. 2015, 11, e1105481. [Google Scholar]
- Das, D.; Arur, S. Conserved insulin signaling in the regulation of oocyte growth, development, and maturation. Mol. Reprod. Dev. 2017, 84, 444–459. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, R.S. Genetic analysis of insulin signaling in Drosophila. Trends Endocrinol. Metab. 2002, 13, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Gulia, M.; Clark, K.D.; Dhara, A.; Crim, J.W. Two insulin-like peptide family members from the mosquito Aedes aegypti exhibit differential biological and receptor binding activities. Mol. Cell. Endocrinol. 2010, 328, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iga, M.; Smagghe, G. Relationship between larval-pupal metamorphosis and transcript expression of insulin-like peptide and insulin receptor in Spodoptera littoralis. Peptides 2011, 32, 531–538. [Google Scholar] [CrossRef]
- Al Baki, M.A.; Jung, J.K.; Kim, Y. Regulation of hemolymph trehalose titers by insulin signaling in the legume pod borer, Maruca vitrata (Lepidoptera: Crambidae). Peptides 2018, 106, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Al Baki, M.A.; Lee, D.; Jung, J.K.; Kim, Y. Insulin signaling mediates previtellogenic development and enhances juvenile hormone-mediated vitellogenesis in a lepidopteran insect, Maruca vitrata. BMC Dev. Biol. 2019, 19, 14. [Google Scholar] [CrossRef] [Green Version]
- Ding, B.Y.; Shang, F.; Zhang, Q.; Xiong, Y.; Yang, Q.; Niu, J.Z.; Smagghe, G.; Wang, J.J. Silencing of two insulin receptor genes disrupts nymph-adult transition of alate brown citrus aphid. Int. J. Mol. Sci. 2017, 18, 357. [Google Scholar] [CrossRef]
- Puig, O.; Tjian, R. Transcriptional feedback control of insulin receptor by dFOXO/FOXO1. Genes Dev. 2005, 19, 2435–2446. [Google Scholar] [CrossRef] [Green Version]
- Puig, O.; Marr, M.T.; Ruhf, M.L.; Tjian, R. Control of cell number by Drosophila FOXO: Downstream and feedback regulation of the insulin receptor pathway. Genes Dev. 2003, 17, 2006–2020. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Liu, J.; Li, C.R.; Momen, B.; Kohanski, R.A.; Pick, L. Deletion of Drosophila insulin-like peptides causes growth defects and metabolic abnormalities. Proc. Natl. Acad. Sci. USA 2009, 106, 19617–19622. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Voorrips, R.E.; Steenhuis-Broers, G.; Van’t Westende, W.; Vosman, B. Reduced phloem uptake of Myzus persicae on an aphid resistant pepper accession. BMC Plant Biol. 2018, 18, 138. [Google Scholar] [CrossRef] [Green Version]
- Sedaghat, A.R.; Sherman, A.; Quon, M.J. A mathematical model of metabolic insulin signaling pathways. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E1084–E1101. [Google Scholar] [CrossRef] [PubMed]
- Nijhout, H.F.; Callier, V. A new mathematical approach for qualitative modeling of the insulin-TOR-MAPK network. Front. Physiol. 2013, 4, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, D.Y.; Kim, Y. Transcriptome analysis of female western flower thrips, Frankliniella occidentalis, exhibiting neo-panoistic ovarian development. PLoS ONE 2022, 17, e0272399. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, C.-Y.; Kim, Y.-G. Insulin-like Peptides of the Western Flower Thrips Frankliniella occidentalis and Their Mediation of Immature Development. Insects 2023, 14, 47. https://doi.org/10.3390/insects14010047
Kim C-Y, Kim Y-G. Insulin-like Peptides of the Western Flower Thrips Frankliniella occidentalis and Their Mediation of Immature Development. Insects. 2023; 14(1):47. https://doi.org/10.3390/insects14010047
Chicago/Turabian StyleKim, Chul-Young, and Yong-Gyun Kim. 2023. "Insulin-like Peptides of the Western Flower Thrips Frankliniella occidentalis and Their Mediation of Immature Development" Insects 14, no. 1: 47. https://doi.org/10.3390/insects14010047